Keyora Astaxanthin EP-17: The Luminosity Architecture: Endogenous Modulation Of Melanogenesis And Dermal Microcirculation
By Keyora Research Notes Series
This article contributes to Keyora’s ongoing scientific documentation series, which systematically outlines the conceptual foundations, mechanistic pathways, and empirical evidence informing our research and development approach.
ORCID: 0009–0007–5798–1996

The Exogenous Bleaching Fraud
Why topical acids and brightening serums biologically accelerate dermal darkening and vascular stagnation.
You stand before the mirror at 2:47 AM, the harsh LED vanity light exposing a reality that no amount of expensive clinical packaging can hide. Your skin, once the resilient armor of a high-achiever, now feels like a failing infrastructure.
You have followed the protocols of the luxury industry to the letter – applying the high-percentage Vitamin C serums that sting upon contact, the glycolic acid toners that promise a “new skin” effect, and the aggressive retinol creams that leave your face in a state of perpetual peeling.
For forty-eight hours, the illusion holds; the chemical exfoliation strips away the dead corneocytes to reveal a temporary, translucent glow. But by the third day, the “Texture of Pain” returns.
Your skin feels tight, hot, and paradoxically “angry.” The spots you were trying to erase appear darker, framed by a chronic, underlying redness that hints at a deeper systemic failure. This is not the “glow of health” promised by the marketing copy; it is the physiological scream of a biological barrier that has been chemically disarmed.
You are trapped in what Keyora defines as The Vicious Cycle of exogenous bleaching – a desperate attempt to fix the surface while inadvertently fueling the internal fire that produces melanin as a survival mechanism.

The Illusion Of Surface Brightening
The thermodynamic cost of chemical peeling.
The cosmetic industry operates on a philosophy of “destruction for renewal,” a high-risk gamble that assumes the body will always respond to injury with perfect repair.
When you apply a topical acid, you are not merely “brightening” your skin; you are initiating a controlled chemical burn.
This process carries a heavy thermodynamic cost that the modern consumer is rarely told about. In the pursuit of a porcelain-like finish, you are systematically dismantling the very hardware designed to protect your internal environment from the entropic forces of the outside world.
This surface-level victory is a biological fraud because it ignores the fundamental law of homeostasis: any violent disruption of the skin barrier triggers a compensatory cascade of inflammation.
By stripping away the outer layers, you are forcing your cells to redirect energy away from radiance and toward emergency structural stabilization. The result is a skin profile that looks “white” only because it is thin, fragile, and pathologically exposed.

Firstly, The Acid Peel Disaster:
The superficial allure of Alpha-Hydroxy Acids (AHA) and Beta-Hydroxy Acids (BHA) masks a brutal mechanical reality. These molecules function by aggressively lowering the pH of the stratum corneum, which violently dissolves the delicate lipid mortar that binds your skin cells together.
According to the research on Linoleic Acid (LA), this essential n-6 fatty acid is the primary structural foundation of acyl-ceramides, the molecular glue that maintains the epidermal barrier.
When you douse your face in exogenous acids, you are not just removing dead cells; you are liquefying the Linoleic Acid infrastructure that prevents transepidermal water loss. This chemical dissolution leaves the “protein bricks” of your skin floating in a disorganized, leaky mess.
As the pH drops, the enzymes responsible for lipid synthesis are inhibited, meaning the more you peel, the less your skin is able to rebuild its own protective shield. This is the definition of a hardware failure, where the “software” of your skincare routine is deleting the essential “drivers” of your skin’s barrier function.
Secondly, The Structural Nakedness:
Once the lipid mortar is compromised, your skin enters a state of structural nakedness that invites a cascade of oxidative assaults.
Without the protective envelope of Linoleic and Alpha-Linolenic acids, the basal layer – the birthplace of your skin cells – is left completely exposed to the high-energy wavelengths of UVA and UVB radiation. This exposure is catastrophic at a molecular level.
Environmental radiation penetrates deep into the dermis, where it generates Singlet Oxygen and Reactive Oxygen Species (ROS) that trigger a process called lipid peroxidation. In this state, the fatty acids in your cell membranes begin to “rust” from the inside out.
As documented in the Keyora Astaxanthin research, this oxidative stress is the primary driver of collagen degradation. Because your skin is now “naked,” these free radicals can effortlessly reach the melanocytes, the Melanin Factories of the skin.
The lack of a biological shield means that every photon of light is now perceived as a lethal threat, forcing the body to deploy its most primitive defense: the production of dark pigment to shade the underlying DNA.
Thirdly, The Inflammatory Rebound:
The final stage of this exogenous fraud is the inevitable inflammatory rebound, which Keyora characterizes as a localized Neuro-Endocrine Storm.
The chemical stripping of the barrier sends an immediate alarm signal through the toll-like receptors of the immune system.
This triggers the activation of NF-KB, a master genetic switch that ramps up the production of pro-inflammatory cytokines like IL-6 and enzymes like COX-2.
This is the moment your “brightening” routine turns into a “darkening” disaster. Inflammation is the most potent biological trigger for the enzyme Tyrosinase, the “on switch” for melanin production.
Your body, sensing that its physical barrier has failed, overcompensates by flooding the skin with melanin in a desperate attempt to absorb the incoming radiation and quench the fire of inflammation. This is why, after a week of “brightening” treatments, you often find your hyperpigmentation returning with a vengeance – darker, deeper, and more stubborn than before.
You have created a Vicious Cycle where the cure is the cause, and the only way out is to stop the chemical assault and begin the process of internal bio-architectural restoration.

The Anatomy Of Dullness:
Hypoxia And Hyper-Pigmentation
Deconstructing the biological roots of a lifeless complexion and the overactive melanocyte alarm system.
To achieve true luminosity, we must first accept a brutal clinical truth: the “yellowness” and “dark spots” that haunt your reflection are two entirely different biological failures occurring in separate layers of the dermal architecture.
One is a vascular crisis of stagnation; the other is a genetic panic response triggered by an unprotected barrier. Most luxury brightening products fail because they treat the skin as a flat canvas rather than a three-dimensional biological machine. They attempt to bleach the surface while ignoring the “dead plumbing” and the “overheated sensors” underneath.
When you look in the mirror and see a sallow, greyish-yellow tint, you are not looking at a lack of Vitamin C; you are looking at the spectral signature of oxygen deprivation and the accumulation of metabolic waste.
When you see a jagged dark spot, you are looking at the “biological scar” of a melanocyte that has been pushed into a state of permanent alarm.
Understanding this distinction is the first step toward reclaiming The Luminous Matrix. It requires a shift from surface aesthetics to deep bio-architectural restoration.

The Stagnant Dermis
Why your skin lacks the translucent glow of vitality.
The cosmetic industry has perpetuated a lucrative lie: that skin radiance is solely determined by the absence of melanin. This is a fundamental misunderstanding of human optics.
True luminosity is generated by the “Backlight Effect” – the way light penetrates the translucent epidermis, bounces off the vibrant, oxygenated blood in the dermis, and reflects back to the observer.
If your skin looks yellow, dull, or “lifeless,” it is because your internal lighting system has failed. The “yellowness” that characterizes a burnout victim’s complexion is rarely caused by pigment; it is caused by the physiological decay of the dermal plumbing.
When the microcirculation slows to a crawl, the skin loses its ability to reflect light with a healthy “pink” frequency.
You are essentially looking at a building where the windows are clean, but the internal lights have been cut off due to a grid failure.

I. The Microcapillary Collapse:
The structural foundation of skin radiance lies in the dermal papillae – the finger-like projections where the dermis meets the epidermis.
Within each of these papillae resides a terminal capillary loop, a microscopic irrigation system responsible for delivering life-sustaining oxygen and nutrients to the basal layer.
In a state of chronic stress or advanced aging, these loops do not just “slow down”; they physically shrivel and collapse. This is often driven by the unregulated activation of Matrix Metalloproteinases (MMPs), enzymes that chew through the collagen scaffolding that keeps these tiny vessels open.
As documented in the Keyora Research on Docosapentaenoic Acid (DPA), the loss of endothelial integrity leads to a “vascular desert.” When these terminal loops shrivel, the distance between the blood supply and the skin’s surface increases. The result is a mechanical disconnection from your body’s primary source of vitality.
Without the structural support of healthy lipids and antioxidants like Astaxanthin to quench the ROS that trigger MMPs, the skin’s “irrigation grid” becomes a graveyard of collapsed vessels.
II. The Hypoxic Yellowness:
The physics of a dull complexion is rooted in the “Stagnant Loop Hypothesis.”
When the microcapillary network collapses, the movement of blood through the dermis transitions from a high-velocity arterial flow to a sluggish, venous crawl. This creates a state of localized hypoxia – a lack of oxygen. Arterial blood, rich in oxyhemoglobin, carries a bright, vibrant red frequency that provides the “rosy glow” of health.
However, as blood stagnates, it becomes deoxygenated. Deoxyhemoglobin has a much darker, purplish-yellow spectral profile.
Because the epidermis is a translucent filter, this stagnant, oxygen-depleted blood pools in the dermis and projects a sickly, sallow hue through the skin. This is the “Hypoxic Yellowness” that no topical cream can erase. It is a signal of a “Biological Backlash” where the skin is suffocating from the inside out.
To fix this, one must not bleach the skin, but rather engineer a “Vascular Re-ignition” by improving endothelial function and reducing the lipid peroxidation that makes the blood “sticky” and the vessels brittle.
III. The Melanocyte Alarm System:
While the vascular system stagnates, a second, more aggressive failure is occurring in the basal layer: the activation of the Melanocyte Alarm System.
Your melanocytes are not “evil” cells designed to ruin your complexion; they are highly sensitive biological sentinels. Their primary mission is to protect the nuclear DNA of your skin cells from the lethal energy of Ultraviolet (UV) radiation and Reactive Oxygen Species (ROS).
As we established in Section 0.0, when the lipid barrier is chemically stripped or oxidatively damaged, the skin becomes “naked.”
When UV photons strike this unprotected tissue, they generate a massive surge of Singlet Oxygen. This oxidative stress acts as a chemical “911 call” to the melanocytes.
In a state of panic, the melanocytes activate the enzyme Tyrosinase, which begins the rapid synthesis of melanin to “shade” the underlying structures.
This is the root of the “Vicious Cycle”: the more “angry” and inflamed the skin becomes due to barrier failure and vascular stagnation, the more pigment it produces as a desperate defense mechanism. The dark spots you see are the visual evidence of a system that is stuck in a permanent state of [The Neuro-Endocrine Storm].

The 7-Molecule Luminous Matrix
Deploying the absolute endogenous protocol to terminate the melanin factory and flush the stagnant vascular swamp.
Surface bleaching is a biological dead end.
For decades, the luxury skincare industry has sold the “Sandblasting Myth” – the idea that you can achieve a luminous complexion by simply traumatizing the surface of your skin until it yields a temporary, thin-walled glow. This approach is not just ineffective; it is a violation of biological law.
To achieve absolute aesthetic sovereignty over your skin tone, you must abandon the exogenous assault and instead deploy a highly coordinated, 7-molecule internal strike force.
True luminosity is an emergent property of a system in perfect metabolic balance. It requires a “Source Blockade” that intercepts the melanin signal at the genetic level while simultaneously “power-washing” the stagnant vascular beds that project a sallow, sickly hue.
This is not a “beauty routine”; it is a bio-architectural intervention designed to replace the fragile, reactive skin of a burnout victim with the resilient, light-reflective matrix of a high-performer.

The Endogenous Paradigm Shift
The 1+1+1+1+1+1+1 Greater Than 7 Equation.
The Keyora Luminous Matrix operates on a principle of synergistic integration, where the total biological output far exceeds the sum of its individual parts.
In the “Scientific Noir” reality of dermal decay, a single-molecule solution – like a high-dose Vitamin C pill – is akin to bringing a candle to a blackout. It is quickly extinguished by the overwhelming tide of oxidative stress.
To truly modulate the “Melanin Factory,” we must deploy a multi-layered defense that addresses every anatomical failure point simultaneously.
This “7-Molecule Equation” targets the cell nucleus, the mitochondrial membrane, the epidermal lipid barrier, and the deep dermal capillaries in a synchronized “Phased Attack.”
By saturating the tissue with this specific ratio of lipids and antioxidants, we shift the skin’s state from “Emergency Defense” (hyperpigmentation) to “Structural Vitality” (translucency).
Every molecule in this matrix has been selected for its ability to penetrate the blood-skin barrier and execute a precise, forensic mission within the cellular architecture.

Firstly, The Transmembrane Commander:
The core of the Luminous Matrix is Natural Astaxanthin, the “Transmembrane Commander” that physically re-engineers the defense system of the melanocyte.
Unlike conventional antioxidants that merely float in the watery cytoplasm, Astaxanthin possesses a unique molecular structure with two polar heads and a long, non-polar polyene chain.
This allows it to span the entire phospholipid bilayer, effectively “bolting” itself across the cell membrane to provide 360-degree protection.
I. The Genetic Muzzle:
Astaxanthin acts as a potent downregulator of the MITF (Microphthalmia-associated transcription factor) gene pathway.
By intercepting the inflammatory signals that reach the nucleus, it prevents the genetic “command center” from ever issuing the order to produce new pigment.
II. The Singlet Oxygen Sink:
In the presence of UV radiation, the skin generates Singlet Oxygen, a highly reactive and destructive form of oxygen that acts as the primary “911 call” for melanin.
Astaxanthin is 6,000 times more potent than Vitamin C at quenching this specific radical, effectively silencing the “panic signal” before it can trigger the Tyrosinase enzyme.
III. The Mitochondrial Firewall:
By embedding itself into the mitochondrial membranes of the basal cells, Astaxanthin prevents the “oxidative leaks” that lead to cellular exhaustion and dullness, ensuring that new skin cells are born with high metabolic energy and a vibrant, translucent structure.

Secondly, The Vascular And Inflammatory Sweepers:
While Astaxanthin provides the blockade, the triad of Alpha-Linolenic Acid (ALA), Eicosapentaenoic Acid (EPA), and Docosapentaenoic Acid (DPA) functions as the “Vascular and Inflammatory Sweepers” to clear the biological debris that creates a “muddy” complexion.
I. The ALA-EPA Conversion Cleanup:
Alpha-Linolenic Acid (ALA) is the master precursor that, once ingested, converts internally into EPA.
This EPA functions as a “molecular vacuum,” activating the phagocytosis process to clear away old, stubborn melanin granules and post-inflammatory hyperpigmentation (PIH) that have been trapped in the deeper dermal layers for years.
II. The DPA Vascular Ignition:
Docosapentaenoic Acid (DPA) is the “Architect of the Dermis.”
As documented in the Keyora DPA research, it possesses unique angiogenic activity that upregulates VEGF (Vascular Endothelial Growth Factor).
It physically mobilizes endothelial progenitor cells to repair the shriveled microcapillary loops we identified in Section 0.1.
III. The Arterial Flush:
By restoring the integrity of the terminal capillaries, DPA and EPA together “flush” the stagnant, deoxygenated venous blood out of the dermis.
This allows a fresh surge of bright red, oxyhemoglobin-rich arterial blood to fill the tissue, providing the internal “backlight” that projects a pink, vibrant radiance through the epidermis.

Thirdly, The Structural Restorers:
The final layer of the Luminous Matrix consists of Linoleic Acid (LA) and Oleic Acid (OA), the “Structural Restorers” responsible for rebuilding the skin’s physical armor and ensuring a natural, trauma-free cell turnover.
I. The Lipid Mortar Synthesis:
Linoleic Acid (LA) is the essential building block for acyl-ceramides, the molecular “mortar” of the stratum corneum.
By providing the raw materials for a healthy barrier, LA fixes the “leaky roof” of the skin, preventing the environmental irritants and pollutants from ever reaching the deeper layers to trigger new inflammation.
II. The OA Desquamation Catalyst:
Oleic Acid (OA) serves as a “Bio-Lubricant” that activates AMPK pathways and facilitates smooth, enzymatic desquamation.
In a healthy Luminous Matrix, the skin does not need to be “peeled” with acids; it naturally and invisibly sheds its old, melanin-filled cells because the “biochemical gears” of cell turnover are perfectly greased.
III. The Radiant Finish:
When the barrier is structurally sound (thanks to LA) and the cell turnover is efficient (thanks to OA), the skin’s surface becomes optically smooth at a microscopic level.
This creates a “Mirror Effect” where light is reflected evenly rather than being scattered by rough, dry patches.
The result is a complexion that looks “lit from within” because it is finally functioning as a healthy, high-performance biological machine.

The Protocol Track:
Eradicating The Shadows
A high-readability guide to shutting down the pigment factory and turning on the internal lights.
Strip away the complex biochemistry for a moment.
Forget the academic density of MITF Downregulation and the microscopic physics of Capillary Atrophy.
If you are reading this, it is likely because your reflection has become a source of silent despair – a landscape of yellow stagnation, dull exhaustion, and jagged dark spots that refuse to budge.
You have been told that the solution lies in a bottle of expensive acid or a clinical-strength bleaching cream. This is a lie designed to keep you in a cycle of consumption and irritation.
To achieve absolute aesthetic sovereignty and restore your skin’s natural, high-performance radiance, you only need to execute three brutal rules. These rules are designed to stop the surface sabotage, confiscate the biological machinery of your pigment factory, and flush the internal swamp that is suffocating your glow.
This is the “Track B” protocol – the actionable reality of the Keyora Luminous Matrix.

Rule 1: Stop Sandpapering The Fire Alarm
The truth about your brightening serums.
The most common mistake in the pursuit of “glass skin” is the belief that you can sand your way to luminosity.
When you apply high-concentration acids to your face, you are engaging in a form of biological warfare against your own barrier. You have been led to believe that “peeling” is a sign of progress, but in the world of “Scientific Noir,” peeling is a sign of structural failure.
Outside-in approaches fail because they address the symptom while aggressively worsening the cause.
By stripping your skin, you are not just removing dead cells; you are removing your primary defense mechanism against the world.
The Chemical Burn:
Using acids to peel your skin is exactly like using sandpaper on a fire alarm. Your skin’s “alarm” is the melanocyte, the cell responsible for producing pigment.
When you scrub and dissolve the protective lipid barrier with AHA or BHA, the melanocyte perceives this as a lethal threat. It panics and begins to ring the alarm by pumping out massive amounts of dark melanin to “scab over” the perceived injury.

I. The Acidic Assault:
Every time you apply a stinging toner, you are dissolving the Linoleic Acid mortar that keeps your skin waterproof and resilient.
This causes an immediate drop in pH that shocks the underlying cells into a state of emergency.
II. The Defensive Melanin:
Your body does not know you are trying to look “bright.” It only knows that its armor is gone.
It responds by manufacturing dark spots as a biological shield against the UV light that can now penetrate deep into your naked tissue.
III. The Chronic Redness:
This constant state of irritation creates a low-grade, permanent fire in your dermis.
This “fire” activates the COX-2 enzyme, which ensures that even if you manage to bleach a spot, the underlying inflammation will simply cause it to return darker and deeper than before.

Rule 2: Confiscate The Factory’s Machinery
Shutting down the melanin production line.
If Rule 1 is about stopping the sabotage, Rule 2 is about taking control of the internal production line.
You cannot stop hyperpigmentation by simply wiping away the “ink” on the surface.
You must go into the factory and confiscate the machines.
This requires an internal strike team that can penetrate the cell membrane and stay there.
This is the specific mission of Astaxanthin.
Because it is a lipid-soluble transmembrane commander, it does not just wash off; it embeds itself into the very fabric of your skin cells to act as a permanent blockade.
The Internal Sabotage:
You must swallow Astaxanthin to send a biological strike team directly into the pigment factory.
Once inside, this molecule acts as an elite saboteur, permanently unplugging the machines that make dark spots.
It quenches the oxidative “sparks” that start the production line, ensuring that the factory stays dark and silent even when you are under stress or exposed to the sun.

I. The Membrane Infiltration:
Astaxanthin’s unique shape allows it to “bolt” itself across your cell membranes.
This creates a 360-degree firewall that blocks the “panic signals” from reaching the nucleus of your melanocytes.
II. The Power Cut:
Melanin production requires an enzyme called Tyrosinase.
Astaxanthin essentially cuts the power to this enzyme by quenching the Singlet Oxygen that it needs to function.
Without power, the factory cannot produce a single granule of pigment.
III. The Permanent Blockade:
Unlike topical Vitamin-C which oxidizes on your skin in minutes, internal Astaxanthin stays in your tissues for days.
It provides a constant, 24-hour blockade that prevents new spots from forming while your body slowly clears the old ones.

Rule 3: Flush The Swamp With Fresh Rivers
Pumping the yellow stagnation out of your face.
The final rule addresses the “yellowness” and “greyness” that characterize an exhausted complexion. This is not a pigment problem; it is a plumbing problem.
If your skin looks sallow, it is because the “rivers” of blood underneath are no longer flowing. You are looking at a biological swamp.
To fix this, you must engage in a “Vascular Re-ignition.”
You must flush out the old, dirty, oxygen-depleted water and replace it with a surge of fresh, vibrant, bright-red arterial blood.

The Dead Plumbing:
Your face is yellow because the microscopic water pipes underneath are clogged with dead, dirty blood.
In a state of burnout, your capillary loops shrivel up like dry garden hoses. The blood that stays trapped in these shriveled pipes loses its oxygen and turns a dark, muddy, yellowish-purple hue.
This stagnant “swamp water” projects its sickly color through your skin, making you look tired no matter how much you sleep.
I. The Stagnant Pool:
Without the specific lipids DPA and EPA, your blood becomes “sticky” and your vessels become brittle. This causes the blood to pool in your cheeks and under your eyes, creating a sallow, “bruised” look that won’t wash off.
II. The Pipe Replacement:
Swallowing the Keyora Omega lipid matrix (ALA, DPA, and EPA) provides the raw materials to build brand new pipes.
DPA, in particular, acts as a master plumber, physically triggering the growth of fresh, healthy capillary loops to replace the shriveled ones.
III. The Arterial Flush:
As these new pipes come online, they flush out the yellow swamp water. A fresh surge of bright, oxygen-rich arterial blood fills your dermis.
This provides the “pink” frequency that creates a true, healthy glow. It turns on the “internal lights” of your skin, making it look translucent and vibrant from the inside out.
The Barrier Seal:
Finally, the Linoleic and Oleic acids in the matrix seal the deal. They rebuild your “mirror” surface so that the fresh light from your new blood can reflect perfectly to the world.
You are no longer “brightened” – you are luminous.

Chapter 1: The Oxidative
Trigger And The Genetic Blockade
Deconstructing the reactive oxygen alarm system, downregulating the MITF transcription factor, and cutting the power to the melanin factory at the source
To understand the biological reality of your skin tone, you must first abandon the superficial vanity of the cosmetic counter and enter the cold, forensic theater of evolutionary biology.
Melanin is not a cosmetic flaw; it is a biological bulletproof vest, forged over millions of years of selective pressure to ensure the survival of your species. When you look in the mirror and lament the emergence of a dark spot or a sallow, deepening tan, your melanocytes are not “failing” you.
In fact, they are working with frantic, high-resolution efficiency to save your life. They are reacting to an invisible, high-energy assault that threatens the very core of your existence: the nuclear DNA within your basal cells.
Every increase in pigment is a signal that your “biological sensors” have detected a lethal threat – a radiation-induced mutational event that could lead to cellular apoptosis or malignant transformation.
In the “Scientific Noir” of the human body, your skin is a fortress under constant bombardment, and the melanocyte is the elite security guard tasked with deploying the blast shields. When you complain about getting darker, you are essentially complaining that your security system is working too well.
To achieve true luminosity without compromising safety, we must learn to deactivate the alarm, not just paint over the siren.

I. The Misunderstood Enemy
The evolutionary purpose of hyperpigmentation.
The global skincare industry has successfully villainized melanin, framing it as an “imperfection” to be eradicated through chemical violence. This marketing narrative is a dangerous biological inversion.
Melanin is actually one of the most sophisticated and efficient energy-management molecules in the known universe. It exists because the alternative is biological death.
If the human body had not evolved the ability to synthesize this dark pigment, the high-energy photons from solar radiation would have long ago rendered our ancestors sterile or dead through the systematic destruction of their genetic code.
Hyperpigmentation is not a disease; it is an immune strategy designed to maintain the structural integrity of the human genome under the intense thermodynamic pressure of a G-type main-sequence star.
Firstly, The Radiation Threat:
Ultraviolet (UV) photons are not merely “light”; they are high-velocity particles carrying enough kinetic and thermodynamic energy to physically shatter the covalent bonds of your molecular structure.
When a UVB photon strikes an unprotected keratinocyte in the basal layer, it induces a catastrophic event known as a cyclobutane pyrimidine dimer (CPD). This is a literal “genetic kinking” where adjacent DNA bases are fused together, making the code unreadable and prone to lethal mutation.
Without a blockade, this radiation causes a chain reaction of Singlet Oxygen generation, leading to a state of “oxidative chaos” that the cell cannot survive.
Secondly, The Melanin Shield:
Melanin functions as a high-performance semiconductor and a universal energy sink. Because of its unique electronic structure – specifically its dense system of conjugated double bonds – it is able to absorb up to 99.9% of the incident UV radiation that hits it.
Instead of allowing this energy to reach and shatter the DNA, melanin captures the photons and undergoes a process of non-radiative relaxation.
It instantly converts the lethal radiation into harmless heat, which is then safely dissipated into the surrounding tissue. This is a form of “biological engineering” that turns a weapon of destruction into a manageable thermal event.
Thirdly, The Biological Priority:
Your body operates under a “Prime Directive” of homeostasis and survival, which places aesthetic brightness at the very bottom of its priority list.
To the internal regulatory systems, a “bright, porcelain complexion” is a sign of extreme vulnerability and structural nakedness.
Consequently, when you apply surface bleaching agents to strip away this protection, the body perceives it as a direct attack on its immune strategy.
This triggers a massive, reactive “Melanin Surge” as the basal layer attempts to restore its bulletproof vest as quickly as possible.
This is why surface-level “whitening” is a biological dead end – it forces the skin into a permanent state of emergency.

II. The Basal Layer Sentinel
The anatomical positioning of the pigment factory.
To intercept the pigment production line, we must first locate the “factory floor.”
The melanocyte is a specialized, neural-crest derived cell that resides deep within the Stratum Basale, anchored firmly to the basement membrane at the dermal-epidermal junction. It is a solitary sentinel, strategically positioned to monitor the health of the entire skin architecture.
Unlike the keratinocytes that surround it, which are constantly moving upward toward the surface to die and flake off, the melanocyte is a long-term resident.
It stays in its position for years, acting as a permanent “security hub” that manages the pigmentation of a specific territory of skin cells.
Firstly, The Dendritic Reach:
The melanocyte does not act in isolation; it is the center of what dermatologists call the “Epidermal Melanin Unit.”
From its central cell body, it extends long, octopus-like tentacles called dendrites. These dendrites weave their way between the surrounding skin cells, physically connecting a single melanocyte to as many as 36 individual keratinocytes. This dendritic reach allows the sentinel to monitor the oxidative status of its entire neighborhood.
If any one of those 36 cells sends out a “distress signal” in the form of reactive oxygen species (ROS), the melanocyte instantly prepares for a defensive deployment.
Secondly, The Parasol Mechanism:
Once the alarm is triggered, the melanocyte begins the rapid synthesis of melanosomes – microscopic storage tanks filled with melanin. These tanks are then transported along the dendrites like cargo on a conveyor belt, powered by molecular motors such as kinesin and dynein.
Upon reaching their destination, these melanosomes are “injected” into the keratinocytes. Crucially, the keratinocytes then organize these pigment granules into a “supranuclear cap” – a microscopic parasol that sits directly over the cell’s nucleus.
This parasol acts as a physical shield, ensuring that the sunlight cannot reach the delicate DNA strands hidden beneath it.
Thirdly, The Futility Of Surface Attack:
Because the “factory” (the melanocyte) is located so deep in the basal layer, and its “shielding mission” (the parasol mechanism) is driven by a survival instinct, attacking the surface of the skin with acids is a strategic failure.
As long as the sentinel at the bottom of the stack perceives a threat – whether that threat is UV light, chemical irritation, or internal oxidative stress – it will never stop pumping out pigment.
You can scrub away the top layer of the “parasols,” but the octopus-like sentinel will simply work harder to replace them.
To achieve lasting radiance, we must not attack the guard; we must provide the guard with a “Source Blockade” – an internal environment so saturated with antioxidants like Astaxanthin that the alarm never rings in the first place.

1.1 The ROS Ignition:
Firing The Flare Gun
How ultraviolet radiation weaponizes oxygen to trigger the melanocyte alarm.
The melanocyte has no eyes.
It cannot “see” the sun, nor can it perceive the visible spectrum of light that humans define as a bright, summer afternoon.
It exists in the perpetual, humid darkness of the basal layer, buried beneath multiple layers of keratinocytes and a dense lipid mortar.
To this blind sentinel, the “sun” does not exist as an image, but as a series of chemical distress signals delivered through the intercellular fluid. It only knows to produce melanin when it receives a specific, high-velocity molecular message. That signal is Reactive Oxygen Species (ROS).
In the “Scientific Noir” of the dermal environment, ROS is the chemical smoke that follows the thermodynamic fire of solar radiation. It is the “flare gun” fired by the surface cells to warn the deep tissue that the barrier has been breached.
When you look at your skin and see a tan or a new dark spot, you are witnessing the aftermath of a biological alarm system that was triggered by a “molecular violence” you could neither see nor feel at the moment of impact.

Phase 1: The UV Impact
The thermodynamic collision at the epidermal surface.
The journey of a UV photon begins 93 million miles away and ends in a fraction of a microsecond against the lipid bilayer of your skin. This is not a gentle interaction; it is a high-energy collision that disrupts the fundamental stability of the atoms that constitute your physical self.
When a photon breaches the stratum corneum, it does not simply “bounce off.” It penetrates the tissue until it finds a target with the right electronic resonance.
In most cases, that target is a stable molecule of oxygen or a polyunsaturated fatty acid within your cell membranes. This impact is the primary “Ignition Event” that sets the entire hyperpigmentation cascade in motion.
I. The Energy Transfer:
As the high-energy UV photon – particularly in the UVA range (320 – 400 nm) – descends through the epidermal layers, it collides with the stable molecular oxygen residing in the skin tissue.
This oxygen is normally in a “triplet state,” a relatively calm and unreactive configuration that supports cellular respiration. However, the UV photon carries enough kinetic and electromagnetic energy to physically “kick” the oxygen out of its stable orbit.
This transfer of energy is instantaneous and violent, transforming a life-sustaining element into a weapon of structural destruction.
II. The Electron Excitation:
This collision violently kicks an electron from the oxygen molecule into a higher orbital, a state known as electronic excitation.
In this high-energy configuration, the oxygen molecule becomes a “biological saboteur.” It is no longer governed by the laws of stable respiration; it is now an unstable, hyper-reactive entity seeking to return to its original energy state by stealing electrons from anything in its immediate vicinity.
This “spin flip” of the electron makes the molecule thousands of times more reactive than ordinary oxygen, turning it into a localized source of molecular chaos.
III. The Singlet Oxygen Birth:
This process of excitation and energy transfer instantly births Singlet Oxygen, the most aggressive and damaging form of ROS in the dermal environment. Singlet Oxygen is the primary “Flare Gun” of the skin’s defense system.
Unlike other free radicals that might be neutralized by surface-level Vitamin C, Singlet Oxygen possesses a unique ability to penetrate deep into the lipid matrix. It is the specific molecular trigger that communicates the “emergency” of solar radiation to the deeper layers.
Without a master antioxidant like Astaxanthin to quench this birth at the source, the “Singlet Oxygen Burst” will proceed to dismantle the skin’s architecture from the inside out.

Phase 2: The Singlet Oxygen Burst
The localized biochemical fire.
Once born, Singlet Oxygen does not just float harmlessly in the intercellular space; it behaves like a molecular incendiary device. It immediately attacks its surroundings in a desperate bid to stabilize its own electronic spin.
This localized biochemical fire spreads rapidly through the lipid-rich environment of the epidermis, creating a trail of “molecular wreckage” that the body’s sensors recognize as a state of extreme distress.
This “Burst” is what converts a simple light exposure into a full-scale [Neuro-Endocrine Storm].
I. The Lipid Peroxidation Spark:
Singlet Oxygen specifically targets the double bonds found in the lipids that protect your skin.
It attacks nearby squalene – the primary antioxidant lipid of the skin surface – and the essential polyunsaturated fatty acids like Linoleic Acid (LA) and Alpha-Linolenic Acid (ALA) that form your barrier. This attack initiates a “Radical Chain Reaction” known as lipid peroxidation.
In this state, the fats in your skin begin to “rancidify” or “rust” in real-time. Each lipid molecule that is attacked becomes a new free radical, which then attacks its neighbor, creating a self-sustaining wave of destruction.
II. The Cellular Membrane Damage:
As the lipid peroxidation spreads, it strips electrons from the cell membranes of the surrounding keratinocytes, causing massive structural distress.
The cell membrane, which should be a flexible and resilient shield, becomes brittle and leaky. This “Membrane Breach” is a catastrophic event for the cell. It leads to the leakage of intracellular enzymes and the activation of “danger-associated molecular patterns” (DAMPs).
The keratinocytes, sensing their imminent death or damage, begin to release a cocktail of inflammatory chemicals, turning the localized “spark” into a systemic alarm.
III. The Inflammatory Echo:
This localized damage creates a massive biochemical echo, alerting the entire epidermal neighborhood that a breach has occurred.
This echo is not just a signal; it is an “Inflammatory Command” that activates enzymes like COX-2 and triggers the release of pro-inflammatory cytokines such as IL-1 and TNF-alpha.
This is the moment when the “Texture of Pain” becomes a biological reality, even if you cannot yet feel it.
The skin is now in a state of high-alert, and the biochemical messengers are being prepared for their journey down to the “Melanin Factory” at the base of the epidermis.

Phase 3: The Messenger Creation
Firing the flare gun to the basal layer.
The final phase of the ROS Ignition is the transmission of the alarm.
The localized damage at the surface must now be communicated down to the melanocytes located several layers below. This is done through a process of diffusion and receptor binding.
The ROS molecules and their inflammatory byproducts act as the “messengers of doom,” traveling through the microscopic gaps between cells to reach the “sentinel” that can actually deploy the pigment shields.
I. The ROS Diffusion:
These reactive oxygen species, acting as chemical messengers, diffuse downward toward the dermal-epidermal junction.
Because they are small and highly mobile, they can easily bypass the surface defenses that were destroyed in Phase 2. They travel through the intercellular fluid like smoke from a fire, carrying the specific “chemical signature” of the UV assault.
This diffusion is the “Biological Link” between the surface environment and the deep genetic machinery of the skin.
II. The Receptor Binding:
Once these ROS molecules and their associated inflammatory markers reach the basal layer, they physically bind to specific receptors on the surface of the melanocyte’s dendritic arms. These receptors are the “Eyes” of the melanocyte. They are tuned to detect specific concentrations of oxidative stress and inflammatory signaling.
When a molecule of Singlet Oxygen or a lipid peroxide byproduct binds to these receptors, it is like a key turning in a lock.
The melanocyte “realizes” that its neighbors are under attack and that its own DNA may be the next target.
III. The Ignition Of The Factory:
This receptor binding is the exact “Flare Gun” signal that tells the melanocyte: the skin is burning, deploy the melanin shields immediately.
This signal activates the internal signaling pathways – specifically the cAMP and p38 MAPK pathways – which then travel to the cell nucleus to wake up the MITF gene.
This is the moment of no return in the hyperpigmentation process. The “Factory” has been ignited, the power has been turned on, and the production of dark spots is now a biological certainty unless the signal can be intercepted at the genetic level.

1.2 The MITF Transcription Factor:
The Master Commander
The intracellular communication network that authorizes massive pigment production.
The reactive oxygen species (ROS) flare gun has been fired from the surface and successfully received by the chemical sensors of the melanocyte.
However, the biological factory does not roar into life instantaneously. There is a precise, high-stakes delay – a period of internal deliberation where the molecular message must be translated into a genetic command.
In the Scientific Noir of the cellular landscape, the signal must travel through the dark, viscous cytoplasm, navigate the labyrinthine cytoskeleton, and eventually breach the double-membrane fortress of the nucleus.
Its mission is to awaken the Master Commander: the Microphthalmia-associated Transcription Factor (MITF) gene.
This factor is the absolute sovereign of your skin tone. It sits at the apex of the pigmentation hierarchy, holding the unique authority to either keep the cell in a state of quiet translucency or to initiate a full-scale industrial mobilization of melanin.
Until the MITF gene is activated, the dark spot is merely a potentiality; once it is authorized, the hyperpigmentation becomes an architectural inevitability.
To stop the shadow, we must intercept the messenger before it reaches the Commander’s desk.

Step 1: The Kinase Cascade
Transmitting the distress signal through the cytoplasm.
The signal received at the cell membrane is a chemical whisper that must be amplified into a genetic shout. This is achieved through the Kinase Cascade, a sophisticated “bucket brigade” of enzymes that pass high-energy phosphate groups from one molecule to the next.
In the context of The Neuro-Endocrine Storm, this cascade functions as a biological amplifier, ensuring that even a relatively small amount of UV-induced oxidative stress can trigger a massive and disproportionate production of pigment.
The cytoplasm, once a calm reservoir of nutrients, becomes a high-speed transit corridor for the instructions of war.
Firstly, The MAPK Pathway Activation:
The moment Reactive Oxygen Species (ROS) bind to the receptors on the melanocyte’s dendritic arms, they trigger the activation of the Mitogen-Activated Protein Kinase (MAPK) pathway. This is specifically focused on the p38 signaling cascade, a pathway evolved to manage cellular stress and survival.
The p38 kinase acts as a molecular “first responder.”
It senses the disruption in the cellular redox state and undergoes a conformational change, switching from an “off” to an “on” state. This activation is the first domino to fall in the internal signaling sequence, converting an external environmental threat into an internal biochemical reality.
Secondly, The Phosphorylation Relay:
Once p38 is activated, it initiates a rapid relay of phosphorylation events. This is a process of “Chemical Tagging” where enzymes add phosphate groups to specific proteins, passing the biochemical message deeper into the cell. This relay is designed for speed and precision.
It bypasses the slower metabolic processes of the cell, creating a direct line of communication from the “burning” surface to the deeper regulatory hubs.
Each step in this relay amplifies the signal, ensuring that the warning of DNA damage is loud enough to overcome the cellular noise of everyday maintenance.
Thirdly, The Awakening Of MITF:
This high-speed cascade eventually reaches its terminal destination: the Microphthalmia-associated Transcription Factor (MITF). In a dormant cell, MITF exists in a state of quiet readiness, tethered to inhibitory proteins that keep it from entering the nucleus.
The incoming phosphorylation wave strikes MITF, causing it to break free from its molecular handcuffs. This is the “Awakening of the General.”
MITF is now biologically “armed” and ready to execute its primary function: the absolute regulation of melanocyte survival and the mass-production of the enzymes required for pigment generation.

Step 2: The Nuclear Translocation
Breaching the inner sanctum of the cell.
The Master Commander is now awake and active, but it remains trapped within the cytoplasm, separated from the genetic code by the nuclear envelope.
For the pigmentation command to be executed, MITF must physically move to where the DNA is stored. This process, known as Nuclear Translocation, is a highly regulated and energy-intensive maneuver. The nuclear envelope is not a passive wall; it is a fortress guarded by the Nuclear Pore Complex (NPC), a molecular “security gate” that only allows authorized proteins to pass.
MITF must now present the correct biochemical credentials to enter the inner sanctum of the cell.
Firstly, The Cytoplasmic Detachment:
Before it can move, MITF must undergo a complete cytoplasmic detachment. In its dormant state, it is often anchored to the cytoskeleton or sequestered by chaperone proteins that prevent accidental gene activation.
The phosphorylation signals we discussed in Step 1 act as a “release code,” inducing a shape-change in the MITF protein that allows it to slip away from its anchors.
It becomes a free-floating molecular entity, energized and directed toward the center of the cell by the gradient of signaling molecules.
Secondly, The Nuclear Pore Crossing:
The activated MITF physically translocates across the highly guarded nuclear envelope.
To do this, it utilizes specialized transport proteins called importins. These proteins act as an “Armed Escort,” recognizing the nuclear localization signal (NLS) on the MITF molecule and guiding it through the microscopic channels of the nuclear pore. This crossing is a definitive point of no return.
Once the Master Commander enters the nucleus, it has direct access to the library of your genetic blueprints.
The “Scientific Noir” of the cell reaches its climax here, as the signal breaches the final barrier between environmental stress and genetic response.
Thirdly, The DNA Target Acquisition:
Once inside the nucleus, MITF does not simply float aimlessly. It is a “Search and Bind” molecule with an incredibly high affinity for specific sequences of DNA.
It begins to scan the vast genomic landscape, searching for the exact promoter regions responsible for melanin synthesis. This is a process of forensic precision.
MITF ignores billions of base pairs of irrelevant code, hunting for the specific “start buttons” of the pigmentation factory.
It is looking for the genetic coordinates that will allow it to authorize the assembly of the enzymes that turn a clear complexion into a shadowed one.

Step 3: The Gene Activation
Authorizing the assembly line.
The final stage of the blockade failure occurs when MITF finds its target and binds to the DNA. This is the act of “Genetic Authorization.”
The Master Commander sits down at the control panel and presses the buttons that power on the industrial machinery of the melanocyte. This is the moment where the “Biochemical Logic” of the cell transitions into the “Industrial Logic” of the factory.
The blueprints are pulled from the archives, and the ribosomal assembly lines are ordered to begin the high-speed synthesis of the proteins that will eventually become dark spots on your skin.
Firstly, The Promoter Binding:
MITF executes its command by binding to specific DNA sequences known as E-box motifs. These motifs are short, conserved sequences (CANNTG) that act as the physical “docking bays” for the Master Commander.
When MITF docks at these E-boxes, it recruits other “co-activator” proteins to the site, forming a massive transcriptional complex. This binding is the definitive signature on the “Work Order” for melanin.
It forces the double-helix of the DNA to unwind, exposing the codes for the proteins that define your skin’s pigment profile.
Secondly, The Tyrosinase Transcription:
This genetic binding forces the DNA to transcribe the blueprints for the three essential enzymes of the melanin factory: Tyrosinase (TYR), Tyrosinase-Related Protein 1 (TRP-1), and TRP-2. These molecules are the “Engineers” of the dark spot.
The cell begins to churn out messenger RNA (mRNA) copies of these blueprints at an industrial scale.
This mRNA is then dispatched out of the nucleus and into the cytoplasm, where the ribosomes will read the code and build the physical enzymes.
This is the “Blueprint Dispatch” phase, where the genetic command is converted into tangible, functional proteins.
Thirdly, The Factory Power-Up:
With these genetic blueprints now authorized and dispatched, the melanocyte completes its transition from a dormant sentinel into a hyper-active, melanin-pumping industrial factory.
The internal environment of the cell shifts entirely toward the production and packaging of pigment.
The Golgi apparatus and the endoplasmic reticulum begin to expand to handle the increased workload.
The factory is now fully powered up, and the production of the dark “parasols” we identified in the Chapter 1 introduction is now a biological certainty.
This is why we call MITF the Master Commander; without its authorization, the factory stays dark. With it, the shadows begin to grow.

1.3 Astaxanthin’s Thermodynamic Interception
Confiscating the ROS flare gun before the signal reaches the melanocyte membrane.
To stop pigmentation, we must not wait for the industrial factory of the melanocyte to power up its assembly lines.
We must not wait for the Master Commander to authorize the genetic blueprints or for the tyrosinase enzymes to begin their chemical conversions.
In the Scientific Noir of dermal defense, a reactive approach is a failed approach.
We must intercept the “flare gun” in mid-air, before the chemical signal of distress ever reaches the sensors of the basal layer.
This is the first absolute defensive perimeter established by Astaxanthin – a physical and thermodynamic blockade that operates at the speed of light.
While traditional antioxidants like Vitamin C or Vitamin E float aimlessly in the cellular fluids, waiting for a chance encounter with a free radical, Astaxanthin acts as a pre-positioned, structural interceptor. It does not wait for the fire to start; it removes the oxygen from the spark.
By saturating the skin’s architecture with this master molecule, we create a Luminous Matrix where the very concept of “damage” is neutralized before it can be translated into a “command.”
This is the ultimate “Source Blockade,” a state of biological sovereignty where the skin remains oblivious to the oxidative violence of the outside world.

I. The Transmembrane Deployment
Securing the perimeter of the basal layer.
The efficacy of Astaxanthin is not merely a result of its chemical potency, but of its strategic anatomical positioning. To stop the melanocyte from panicking, we must secure the entire perimeter of the cell, from the outermost lipid envelope to the innermost mitochondrial membrane.
Unlike water-soluble antioxidants that are easily washed away or excreted, Astaxanthin is a lipophilic commander that seeks out the fatty acid structures of the skin. It does not merely visit the cell; it integrates into it, becoming a permanent part of the skin’s hardware.
A. The Deep Dermal Delivery:
The journey of the blockade begins with oral ingestion, bypassing the limited absorption rates of topical creams.
Once it enters the systemic circulation, Astaxanthin travels through the dense network of capillary beds we identified in the previous sections.
Because it is carried by lipoproteins, it easily penetrates the blood-skin barrier, saturating the extracellular matrix that surrounds the melanocytes. This ensures that every single sentinel in the basal layer is bathed in a protective reservoir of the molecule.
The delivery is deep, uniform, and comprehensive, providing a level of saturation that no surface-applied serum could ever hope to achieve.
B. The Bilayer Integration:
The true genius of Astaxanthin lies in its unique molecular geometry.
It possesses two polar ionone rings at either end of a long, non-polar polyene chain. This structure is perfectly calibrated to the architecture of the human cell membrane. Its extreme lipophilicity allows it to dissolve directly into the phospholipid bilayers of the skin cells.
While other carotenoids like Beta-Carotene sit awkwardly within the membrane, causing structural distortion, Astaxanthin aligns itself perfectly with the phospholipid tails.
It “sews” itself into the membrane, providing a stabilized, reinforced shield that guards both the watery interior and the fatty exterior of the cell simultaneously.
C. The Structural Anchoring:
This integration results in a physical anchoring that spans the entire width of the cell membrane – a distance of approximately 30 Angstroms.
This 30-Angstrom span acts as a physical barricade, creating an impenetrable thermodynamic shield precisely where the UV photons strike.
By “bolting” itself across the bilayer, Astaxanthin prevents the membrane from becoming leaky or brittle under oxidative stress. It stabilizes the very “skin” of the cell, ensuring that the receptors on the dendritic arms remain protected from the chaotic collisions of free radicals.
This structural anchoring is the physical foundation of the Luminous Matrix, a permanent fortification against the entropic forces of radiation.

II. The Energy Dissipation
The thermodynamic quench of Singlet Oxygen.
Once the perimeter is secured, the second phase of the blockade begins: the quenching of the oxidative spark.
As we established in Section 1.1, the birth of Singlet Oxygen is the primary “flare gun” that triggers the pigmentation cascade. Singlet Oxygen is not just a molecule; it is a high-energy electronic state that vibrates with destructive potential.
Astaxanthin does not just “neutralize” this threat in a one-to-one chemical reaction; it acts as a thermodynamic “heat sink,” absorbing and dissipating the energy through its massive electron cloud.
A. The Photon Collision:
When UV radiation penetrates the skin, it collides with the stable oxygen molecules in the tissue, attempting to kick them into the hyper-reactive Singlet Oxygen state. In an unprotected cell, this energy transfer is the start of the [Neuro-Endocrine Storm].
However, in an Astaxanthin-saturated environment, the photon encounter is fundamentally altered. The high-energy UV particles must first pass through the dense “forest” of Astaxanthin molecules embedded in the cell membranes.
The collision still occurs, but the outcome is governed by the superior energy-management capabilities of the carotenoid.
B. The Conjugated Absorption:
Astaxanthin’s molecular backbone is a long chain of thirteen conjugated double bonds. This creates a massive, delocalized electron cloud that functions like a biological sponge for high-energy radiation.
When Singlet Oxygen is generated, Astaxanthin’s electron cloud instantly absorbs the erratic oxidative energy.
Because its quenching potency is 6,000 times greater than Vitamin C and 550 times greater than Vitamin E, it can intercept thousands of Singlet Oxygen molecules per second.
The energy is “sucked” into the conjugated system, where it is harmlessly distributed across the entire length of the molecule.
C. The Harmless Heat Release:
Unlike other antioxidants that become “pro-oxidants” or “weak radicals” after they react, Astaxanthin remains stable.
After absorbing the lethal oxidative energy, it simply vibrates. It dissipates the energy as harmless microscopic heat, which is then absorbed by the surrounding water molecules in the tissue.
Once the energy is released, the Astaxanthin molecule returns to its ground state, ready to intercept the next photon. This is a “Continuous Quench” mechanism that allows a single molecule of Astaxanthin to provide thousands of rounds of protection without ever being consumed or damaged.
It is the ultimate form of molecular efficiency.

III. The Alarm Silence
The failure of the chemical messenger.
The biological consequence of this thermodynamic quench is the absolute silence of the melanocyte alarm system.
Because the “flare gun” is intercepted at the exact moment of ignition, the chemical messengers that usually flood the basal layer simply never come into existence.
The “Scientific Noir” of the pigmentation process is cut short; the story ends before the first dark spot can even be conceptualized by the genetic machinery.
A. The ROS Eradication:
Because the Singlet Oxygen was neutralized instantly at the site of its birth, the secondary wave of lipid peroxidation is prevented. There is no “rusting” of the cell membranes, no shattering of the Linoleic Acid mortar, and no release of inflammatory cytokines.
The ROS messengers – the chemical “smoke” that usually alerts the melanocytes – are eradicated at the source.
The environment within the epidermis remains chemically “quiet,” despite the presence of intense solar radiation at the surface.
B. The Receptor Starvation:
The receptors on the melanocyte’s dendritic arms, which we previously described as the “Eyes” of the sentinel, are left in a state of receptor starvation.
They are tuned to detect the chemical signature of oxidative distress, but in the presence of Astaxanthin, that signature is never written.
The dendritic arms wait for a signal that never arrives.
The cAMP and p38 MAPK pathways, which were ready to carry the message to the Master Commander, remain dormant.
The “bucket brigade” of enzymes has no buckets to pass.
C. The Baseline Calm:
Without the ROS flare gun, the melanocyte remains in a state of absolute, undisturbed baseline calm. It is completely oblivious to the UV radiation striking the layers above.
Because the “command center” never receives the order to mobilize, the MITF gene stays asleep, and the production of tyrosinase enzymes is never authorized. The skin maintains its natural, translucent luminosity not because it is “bleached,” but because it is biologically convinced that it is not under attack.
This is the ultimate victory of the Keyora Protocol: achieving radiance through the absolute preservation of cellular peace.

1.4 The Genetic Blockade:
Downregulating MITF
Penetrating the cellular nucleus to revoke the Master Commander’s transcription authority.
Quenching Reactive Oxygen Species (ROS) at the surface of the cell is merely the first line of defense in the Luminous Matrix protocol.
While this thermodynamic interception is vital to stop the initial “flare gun” signal, Astaxanthin executes a secondary, far more profound strike that separates it from the ineffective products found in the Supplement Graveyard.
It acts not just as a passive antioxidant, but as a potent, gene-modulating agent that directly penetrates the melanocyte to sabotage the internal command structure.
In the Scientific Noir of the cellular landscape, Astaxanthin is an infiltrator. It does not just stand at the gate; it enters the building, cuts the communication lines, and physically prevents the Master Commander from ever signing the orders for new pigment. This is a level of “Genetic Sovereignty” that renders surface-level bleaching obsolete.
By de-authorizing the production of melanin at the chromosomal level, we ensure that the “Melanin Factory” remains in a state of permanent, structural dormancy, regardless of the oxidative chaos occurring in the outside world.

A. The Intracellular Penetration
Bypassing the outer defenses of the pigment factory.
Astaxanthin does not merely sit on the outer membrane of the skin cell like a static shield. Its unique molecular architecture – characterized by a dual-polarity profile – allows it to migrate beyond the phospholipid bilayer and enter the viscous interior of the cell.
This is a critical tactical advantage. Most topical antioxidants are physically blocked by the cell’s own security systems, but Astaxanthin possesses the “biochemical credentials” to pass through the gates and execute its mission within the cytoplasm and the nuclear envelope.
It is here, in the inner sanctum of the melanocyte, that the true battle for luminosity is won or lost.
Firstly, The Cytoplasmic Entry:
The molecular architecture of Natural Astaxanthin is not a static barrier; it is a high-mobility interceptor with a dual-affinity profile.
While its long, non-polar polyene chain allows it to anchor firmly into the hydrophobic core of the phospholipid bilayer, its polar ionone rings provide it with the ability to interact with the aqueous environment of the intracellular fluid. This thermodynamic “flexibility” allows Astaxanthin to cross the cell membrane and enter the cytoplasm of the melanocyte.
Once inside, it saturates the intracellular fluid, creating a secondary “Internal Firewall” that protects the organelles from the secondary waves of oxidative stress that often bypass surface-level defenses.
Secondly, The Kinase Interception:
Once it has established a presence within the cytoplasm, Astaxanthin begins its work as a molecular saboteur. It actively interferes with the upstream MAPK (Mitogen-Activated Protein Kinase) and p38 phosphorylation cascades that we identified in Section 1.2.
Astaxanthin acts as a “Kinase Inhibitor,” physically binding to the enzymes responsible for passing the phosphate “signal” down the line.
By quenching the tiny sparks of energy that drive these cascades, it prevents the phosphorylation of the MITF protein. It is like cutting the electricity to a telephone line; the message is still being sent from the membrane, but the wires are dead, and the signal never reaches the deeper departments of the cell.
Thirdly, The Signal Degradation:
The ultimate result of this cytoplasmic infiltration is the total degradation of the internal transmission.
Even if some ROS molecules managed to breach the outer defenses and bind to the receptors on the melanocyte’s dendritic arms, Astaxanthin ensures that the “panic signal” dies in the cytoplasm.
It creates a state of “Information Silence” within the cell. The biochemical messengers that were supposed to carry the warning to the nucleus are intercepted and neutralized before they can execute their mission.
The melanocyte remains biologically “unaware” of the external crisis, maintaining its baseline state of calm because its internal communication network has been successfully compromised by the Astaxanthin blockade.

B. The MITF Suppression
Stripping the Master Commander of its rank.
If the kinase interception is the cutting of the phone lines, then the suppression of MITF is the formal revocation of the General’s authority. This is the genetic masterpiece of the Astaxanthin protocol.
It moves beyond the quenching of chemicals and enters the realm of gene expression modulation. In a cell saturated with the Luminous Matrix, the very presence of the Master Commander – the MITF protein – is systematically reduced.
We are no longer just stopping a signal; we are dismantling the person who is supposed to receive it. By stripping MITF of its rank and its ability to function, we render the entire pigmentation hierarchy powerless.
Firstly, The Transcriptional Downregulation:
Research into the mechanisms of Natural Astaxanthin has revealed that it specifically and potently downregulates the cellular expression of the MITF protein itself.
It doesn’t just block MITF’s activity; it reduces the actual number of MITF molecules synthesized by the cell’s ribosomes.
Through its influence on the cAMP-response element-binding (CREB) protein, Astaxanthin sends a “Cease and Desist” order to the gene that produces MITF. The result is a significant drop in the “Officer Count” within the melanocyte.
With fewer commanders available, the cell’s ability to mount a defensive pigment surge is fundamentally crippled at the source.
Secondly, The Nuclear Exclusion:
For whatever MITF protein remains within the cytoplasm, Astaxanthin establishes a secondary blockade at the nuclear envelope.
By maintaining the integrity of the nuclear pores and preventing the phosphorylation required for entry, it ensures the “Nuclear Exclusion” of the Master Commander.
MITF is left stranded in the cytoplasm, unable to cross the final threshold into the inner sanctum where the DNA is stored. It is like a General who has the keys to the war room, only to find that the locks have been changed.
Without access to the genetic code, the MITF protein is biologically impotent, unable to authorize any new industrial production.
Thirdly, The DNA Disconnect:
The final stage of this genetic sabotage is the “DNA Disconnect.”
Even if a stray molecule of MITF manages to breach the nucleus, Astaxanthin’s influence on the nuclear redox environment makes it difficult for the protein to successfully bind to its target E-box motifs.
The Master Commander attempts to “dock” with the DNA to turn on the melanin factory, but the connection fails. This permanent denial of transcriptional authority is the ultimate “Aha!” moment of the Luminous Matrix.
Without the binding of MITF to the genetic “Start” button, the authorization to build the melanin factory is never granted, and the entire pigmentation process is aborted before it can even begin.

C. The Factory Shutdown
The total cessation of the assembly line.
The biological conclusion of suppressing the Master Commander is the total and absolute shutdown of the melanin assembly line.
In the absence of MITF’s genetic instruction, the melanocyte ceases to function as an industrial factory and returns to its primitive role as a quiet, dormant sentinel. This is not a “masking” of the pigment; it is a cessation of existence.
When the blueprints are never dispatched and the machines are never built, there is no melanin to be found. This is the definition of a “Source Blockade.”
We are not cleaning up a mess; we are ensuring that the mess is never made in the first place.
Firstly, The Tyrosinase Collapse:
The most immediate consequence of the MITF suppression is the total collapse of Tyrosinase (TYR) production.
As we established in Section 1.2, Tyrosinase is the “core machinery” of the pigmentation process. It is the enzyme that converts the amino acid tyrosine into the dark pigments of eumelanin and pheomelanin.
Without the genetic authorization from MITF, the cell’s ribosomes never receive the blueprints for Tyrosinase.
The production line stops dead.
The “Tyrosinase Collapse” means that even if the cell wanted to produce pigment, it lacks the physical tools to do so.
The machinery of darkness has been effectively liquidated.
Secondly, The TRP Depletion:
The shutdown extends beyond the core enzyme to include the supporting structural proteins, Tyrosinase-Related Protein 1 (TRP-1) and TRP-2. These proteins are responsible for stabilizing the melanosomes and ensuring the “quality control” of the pigment granules.
Under the Luminous Matrix blockade, these supporting molecules are also halted.
The “Assembly Line” is not just missing a worker; it has been physically dismantled.
The melanosomes – the storage tanks for pigment – are never formed.
The octopus-like dendrites of the melanocyte have nothing to transport to the surface, resulting in a skin architecture that remains naturally clear and translucent.
Thirdly, The Absolute Source Blockade:
By executing this multi-layered genetic strike, Astaxanthin has effectively cut the power to the melanin factory at its most fundamental level.
No machines are built, no commands are given, and hyperpigmentation is aborted before a single molecule of melanin is ever synthesized.
This is the difference between Keyora’s “Regulation” and the industry’s “Sedation.”
We are not numbing the skin; we are restoring its ability to remain calm in the face of stress. The result is an Absolute Source Blockade – a state where your skin’s luminosity is protected by the most advanced form of genetic engineering known to nutritional science.
You are no longer fighting the shadows; you have removed the light switch.

1.5 Halting Melanocyte Proliferation
Forcing the hyper-active pigment cellular population into a state of dormant stasis.
The final and perhaps most insidious threat posed by chronic UV radiation is not merely the forcing of existing melanocytes to work harder; it is the pathological command for them to multiply.
In the “Scientific Noir” of the dermal landscape, the body does not just increase production in an existing factory; it aggressively expands the factory’s physical footprint.
Chronic sun exposure and sustained oxidative stress trigger a state of melanocyte hyperplasia – a rapid and uncontrolled expansion of the pigment-producing cellular population. This is the biological reality behind the “stubbornness” of age spots and the deep, dense patches of melasma that refuse to fade with topical treatments.
When you look at an age spot, you are not just looking at “extra pigment”; you are looking at a localized “over-population” of melanocytes that have colonized your basal layer.
To achieve a truly luminous matrix, we must go beyond stopping the chemistry of melanin; we must enforce a state of “Population Control” that prevents these cells from cloning themselves and expanding their dark territory.

I. The UV-Induced Hyperplasia
The pathological expansion of the factory.
Under the duress of environmental radiation, the skin enters a state of emergency that bypasses normal regulatory checkpoints. This is a survival mechanism gone wrong.
The body, fearing the total destruction of its genetic heritage, decides that more guards are needed to shield the nucleus. This results in the recruitment and division of melanocyte stem cells, leading to a permanent increase in the sheer number of “factories” operating in a single square millimeter of skin.
This expansion is the structural foundation of a “permanent” shadow.
A. The Stem Cell Factor (SCF) Release:
When UV radiation breaches the upper layers of the epidermis, the damaged keratinocytes do not just release ROS; they fire off a series of powerful mitogenic growth factors.
The most critical among these are Stem Cell Factor (SCF) and Endothelin-1 (ET-1). These molecules function as “Biological Recruitment Posters.” They travel to the basal layer and bind to the c-kit receptors on the surface of melanocytes and their progenitor stem cells.
This binding sends a violent signal to the cell’s internal machinery:
“The defenses have failed. Multiply immediately.”
B. The Cellular Cloning:
Once the SCF and Endothelin-1 signals are received, the melanocytes enter the S-phase of the cell cycle. They begin the energy-intensive process of duplicating their DNA and physically splitting into two identical daughter cells.
This is the “Cellular Cloning” event. In a localized area of high UV damage, the density of melanocytes can double or triple, creating a crowded, high-output zone of pigment production.
This is no longer a regulated defense; it is a localized industrial boom that the skin cannot naturally dismantle.
C. The Macular Formation:
This localized cellular crowding is the biological architecture of a permanent dark spot, technically known as a solar lentigo or a macula.
Because there are now significantly more factories in this specific patch of skin, even a small amount of light trigger results in a disproportionately dark response.
This is why age spots seem to “never go away”; you are fighting a population, not just a pigment. The “Macular Formation” represents a permanent change in the dermal hardware that requires a “Source Blockade” to reset.

II. The Astaxanthin Arrest
Enforcing population control at the basal layer.
To restore the skin’s original, luminous state, we must execute an “Astaxanthin Arrest” – a targeted biological intervention that halts the cloning process and returns the melanocyte population to its healthy, baseline density.
This is not about killing the cells; it is about revoking their authorization to multiply.
By saturating the tissue with the master antioxidant and its synergistic lipid escort, we establish a state of “Dormant Stasis” where the factory footprint is capped and the “cloning signals” are intercepted.
A. The Growth Factor Blockade:
Astaxanthin’s first move is to act as a decoy and a signal-interceptor. Its presence in the extracellular fluid and on the cell membranes of the keratinocytes prevents the massive release of SCF and Endothelin-1.
By quenching the initial oxidative damage that triggers these growth factors, Astaxanthin ensures that the “Recruitment Posters” are never printed.
Without the SCF signal, the melanocyte stem cells remain quiet and dormant, oblivious to the radiation-induced panic that usually drives hyperplasia.
B. The Cell Cycle Arrest:
For those cells that have already begun the process of division, Astaxanthin exerts a secondary influence known as “Cell Cycle Arrest.”
By modulating the p21 and p27 proteins – the “Biological Brakes” of the cell cycle – it forces the hyper-active melanocytes to halt their division. It essentially puts the cloning process on “Pause,” preventing the population from reaching the critical mass required for a permanent dark spot.
This is a form of high-level molecular diplomacy, convincing the skin that the emergency has passed and no more guards are necessary.
C. The Dormant Stasis:
The final result of this population control is the restoration of “Dormant Stasis.” Astaxanthin effectively locks the melanocyte population in a state of healthy, non-proliferative equilibrium.
It caps the maximum capacity of the melanin factory, ensuring that your skin maintains its “Single-Layer Defense” rather than descending into the crowded, high-pigment “Multi-Layer Chaos” of a sun-damaged complexion.
This stasis is the key to long-term radiance; it prevents the shadows from ever gaining the structural foothold they need to become permanent.

1.6 Clinical Consensus:
Astaxanthin’s Suppression Of UV-Induced Pigmentation
The peer-reviewed dermatological validation of the genetic and thermodynamic blockade.
The genetic suppression of the MITF transcription factor and the thermodynamic quenching of Singlet Oxygen are scientifically profound mechanisms, but in the realm of “Scientific Noir,” theory is only the beginning.
These mechanisms must survive the ultimate clinical test: the scrutiny of the peer-reviewed courtroom. They must be validated through rigorous, double-blind, human clinical trials involving direct UV irradiation and high-resolution instrumental measurements. The data from these trials provides the final, undeniable proof that the Luminous Matrix is not a marketing promise, but a physiological reality.
When human subjects are administered 8-16 mg of natural Astaxanthin, as documented in Keyora Research, their skin undergoes a fundamental transformation in its relationship with light.
It moves from a state of “Fragile Reactivity” to a state of “Endogenous Invincibility.”

Proposition:
Oral Astaxanthin Clinically Neutralizes Radiation-Induced Erythema And Aborts The Subsequent Hyperpigmentation Cascade.
The courtroom of evidence-based dermatology.
The scientific consensus on oral Astaxanthin is built upon a foundation of “Evidence Sets” that measure the skin’s response to radiation across multiple timescales.
Before the first dark spot can appear, the skin must first undergo “Erythema” – the initial burn, redness, and inflammation that signals a barrier breach. If we can clinically prove that Astaxanthin stops the burn, we have effectively proven that it stops the pigment.
The “Courtroom of Evidence” shows that oral supplementation creates an “Internal Sunscreen” effect that is far more resilient and uniform than any topical application could ever achieve.

Evidence Set A:
Erythema Prevention
Measuring the immediate thermodynamic quench.
The first evidence set focuses on the immediate aftermath of UV exposure.
In clinical trials, researchers measure the “Minimal Erythema Dose” (MED), which is the smallest amount of UV radiation required to cause visible redness on the subject’s skin. A higher MED indicates a more resilient and protected biological matrix.
Firstly, The MED Elevation:
Clinical trials have consistently demonstrated that individuals taking natural Astaxanthin supplementation (typically 4-12 mg daily) show a statistically significant increase in their MED over a 4-to-9-week period.
In some cases, the skin’s resistance to burning is elevated by as much as 4x compared to the placebo group. This means the skin can withstand vastly more UV radiation before the “flare gun” of erythema is ever fired.
It is the clinical validation of the thermodynamic blockade we discussed in Section 1.3.
Secondly, The Inflammation Drop:
Beyond just raising the threshold for burning, the data shows a drastic reduction in the intensity of redness and heat in those who do eventually reach their MED.
Using laser Doppler flowmetry to measure dermal blood flow, researchers have confirmed that Astaxanthin-saturated skin shows significantly less post-radiation vasodilation.
This proves that the “Singlet Oxygen Burst” and the subsequent “Inflammatory Echo” were successfully quenched, preventing the massive surge of COX-2 and inflammatory cytokines.
Thirdly, The Tissue Preservation:
Biopsy and ultrasound data from these trials conclude that the epidermal matrix was physically protected from the initial radiation strike.
There is less “sunburn cell” formation and less degradation of the underlying collagen fibers. This proves that the Astaxanthin was not just masking the redness, but was actively preserving the structural integrity of the skin at a microscopic level.
The “flare gun” was indeed confiscated in mid-air.

Evidence Set B:
Melanin Reduction
Proving the MITF genetic shutdown.
The second evidence set focuses on the long-term outcome: the actual production of melanin and the formation of dark spots.
This is where the genetic suppression of MITF is visible to the naked eye and to high-resolution instruments.
Firstly, The Pigment Plunge:
Using instrumental data from a Mexameter – a device that measures the specific spectral absorbance of melanin – researchers have proved that in the weeks following UV exposure, the Astaxanthin group showed a profound reduction in melanin formation compared to the placebo group.
Even when the skin was intentionally irradiated, the “Melanin Factory” remained quiet.
The pigment readings plunged, validating that the “MITF command” was successfully intercepted and the gene expression was downregulated.
Secondly, The Spot Fading:
Long-term dermatological assessments (lasting 8-12 weeks) show the visible prevention of new dark spots and the significant fading of existing hyperpigmentation.
High-resolution photography and “Image Analysis” software confirm that the “Melanin Index” of existing age spots decreases over time as the Astaxanthin-saturated skin naturally sheds old pigment and refuses to synthesize more.
This is the visual proof of the “Absolute Source Blockade” in action.
Thirdly, The Ultimate Verdict:
The total body of clinical data absolutely validates Astaxanthin as an endogenous genetic modulator.
It is the only molecule that can travel through the blood-skin barrier, embed into the cell membrane, and successfully cut the power to the human melanin factory.
The verdict is clear: by supplementing with the Keyora Luminous Matrix, you are not just “treating” your skin; you are biologically upgrading its hardware to a state where hyperpigmentation is no longer a necessary or even possible response to the sun.

1.7 The Protocol Track:
Cutting The Power To The Pigment Factory
A high-readability guide to abandoning surface bleaching and biologically shutting down your skin’s dark spot machinery.
Strip away the complex biochemistry for a moment.
Forget the daunting academic density of Singlet Oxygen, the MAPK cascades, and the MITF Transcription Factors.
To the high-functioning executive or the exhausted student, these terms are often just noise in an already overcrowded mind. If your face is constantly producing dark spots, jagged patches of melasma, and a persistent, sallow dullness that no luxury cream can fix, you only need to understand one simple concept: the factory fire alarm.
Your skin is currently a high-stress industrial site where the fire alarms are constantly ringing, and the factory manager is in a state of permanent panic, ordering the assembly lines to churn out black pigment at maximum capacity.
You have been trying to fix this by scrubbing the “black smoke” off the surface, but that has never worked.
To achieve true, deep-tissue radiance, you must move beyond surface cleaning and learn how to biologically cut the power to the entire operation.

Rule 1: The Flare Gun
Why your skin is panicking.
The first step in reclaiming your complexion is understanding why your skin is “ringing the alarm” in the first place.
For years, the skincare industry has taught you that dark spots are just “mistakes” or “aging.”
In reality, they are a highly logical response to a perceived crisis. Your skin is not failing; it is reacting to a invisible chemical distress signal that is being fired every time you step into the light or experience a surge of internal stress.
Firstly, The Sun’s Warning:
You must visualize sunlight hitting your skin as a shower of high-speed bullets. When these bullets strike your skin cells, they fire off microscopic chemical “flare guns” known as free radicals.
These flares are a biological scream that travels through your tissue, shouting: “We are under attack! The barrier is failing! Build the melanin shields immediately!” This is the “ROS Ignition” we discussed in the science track.
Even if you don’t feel a burn, these flare guns are firing by the billions, creating a chemical atmosphere of panic that forces your skin to darken as a desperate form of self-defense.
Secondly, The Bleaching Mistake:
Most people try to fix this by using harsh acids or bleaching serums to scrub away the dark spots. This is a catastrophic strategic error.
Using acids to bleach your skin is like trying to stop a forest fire by painting the burnt trees white while the fire is still raging underneath. The acids actually damage your skin’s protective roof even further, which only makes the flare guns fire faster and louder. This is why your spots often come back darker after a chemical peel.
You are attacking the “smoke” (the pigment) without ever stopping the “flare guns” (the radicals) that are causing the fire.
Thirdly, The Astaxanthin Catcher:
The Keyora solution is to stop the panic at the source.
By swallowing a high-dose, natural Astaxanthin matrix, you are putting millions of microscopic “catchers” into every layer of your skin. These catchers are positioned like an elite security team, standing guard at every cell membrane.
When the sun fires its flare guns, the Astaxanthin catchers snatch them out of the air before they can ever sound the alarm.
Because the catchers are 6,000 times stronger than Vitamin C, they can neutralize a billion flares per second. The alarm never rings, the signal never reaches the factory, and your skin stays calm and clear because it finally feels safe.

Rule 2: The Factory Manager
Firing the boss who orders the dark spots.
Even if you manage to catch some of the flare guns, you still have to deal with the “Factory Manager.” This is the internal authority figure who receives the alarms and decides to turn on the pigment machines.
In the scientific world, this is the MITF gene.
Think of this manager as a high-strung administrator who lives in the basement of your skin cells. As long as he is at his desk, he is ready to authorize the production of new dark spots at the slightest hint of trouble.
To get permanent results, you have to fire the boss.
Firstly, The Boss’s Office:
Deep inside every one of your pigment cells, there is a “manager” whose only job is to oversee the production of melanin. This boss lives in the cell nucleus – the inner sanctum where the genetic blueprints are kept.
When the flare guns go off, the boss pulls the blueprints for “Dark Spot #402” and sends them to the assembly line.
Most skincare products never even reach this office; they stay on the “roof” of the skin, completely unable to talk to the person who is actually in charge of your hyperpigmentation.
Secondly, The Inside Job:
This is where the Luminous Matrix protocol becomes an “Inside Job.”
Because Astaxanthin is a special kind of fat-soluble molecule, it has the “security clearance” to walk straight through the cell walls and enter the manager’s office.
Once inside, it doesn’t just block a few chemicals; it physically interferes with the manager’s ability to work. It sits on his desk, unplugs his phone, and effectively “fires” him.
In technical terms, it downregulates the MITF gene, meaning the commander who orders the pigment is stripped of his authority and sent home.
Thirdly, The Order Canceled:
This is the ultimate victory for your complexion. Without the manager there to give the order, the factory workers have no blueprints to follow.
The assembly line for melanin simply stops. It doesn’t matter how much sun hits your face or how much stress you are under; if the “boss” hasn’t authorized the work, the workers cannot build a single molecule of dark pigment.
By using an internal genetic blockade, you have canceled the order for dark spots at the source, ensuring that your skin stays in a state of high-performance translucency.

Rule 3: The Power Cut
Pulling the plug on the assembly line.
The final rule is about making the results permanent. Once you have caught the flare guns and fired the manager, you must ensure that the “machines” themselves power down and that the factory stops expanding its footprint.
This is the “Power Cut” that finally ends the cycle of dullness and sallow yellowness, allowing your skin’s natural radiance to shine through without interference.
Firstly, The Machines Stop:
In the world of your skin, the “machines” that make the pigment are enzymes like Tyrosinase.
When you cut the power at the manager’s level, these machines lose their energy supply and go to sleep. They stop grinding out the dark melanin that creates spots and the sallow, muddy tone that makes you look exhausted.
As these machines power down, your skin stops fighting itself and begins to focus its energy on repair and luminosity. The “industrial noise” of pigmentation is replaced by the quiet, efficient glow of a healthy biological matrix.
Secondly, The Factory Shrinks:
One of the biggest problems with sun damage is that your skin actually builds “factory extensions” – meaning it creates more pigment cells than it needs. This is why old age spots are so thick and stubborn.
Astaxanthin executes a “Population Control” protocol that stops the factory from building these unnecessary extensions. It prevents your pigment cells from cloning themselves, effectively capping the size of the factory.
Over time, as your skin naturally renews itself, these old factory extensions are cleared away, and they are never replaced.
Your “pigment infrastructure” returns to its original, youthful state.
Thirdly, The Permanent Glow:
By cutting the power at the absolute source – both thermodynamic and genetic – your skin finally experiences what Keyora calls “Luminous Calm.”
You are no longer trapped in a Vicious Cycle of damage and repair.
Your skin stops looking “angry” and starts looking “vitally translucent.”
Because you have fixed the hardware from the inside out, the results are not temporary; they are a permanent upgrade to your skin’s biological architecture.
You have successfully shut down the shadows and turned on the internal lights of the Luminous Matrix.

References:
Ames, B. N., Shigenaga, M. K., & Hagen, T. M. (1993). Oxidants, antioxidants, and the degenerative diseases of aging. Proceedings of the National Academy of Sciences (PNAS), 90(17), 7915 – 7922.
Baliau, R., & Busca, R. (2000). Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Research, 13(2), 60 – 69.
Battie, C., Jitsukawa, S., Bernerd, F., Del Bino, S., Marionnet, C., & Verschoore, M. (2014). New insights in photoaging, skin inflammation and hyperpigmentation. Journal of Investigative Dermatology Symposium Proceedings, 16(1), S6 – S12.
Beani, J. C. (2001). Astaxanthin: a review of its chemistry and clinical applications. Journal of Dermatological Treatment, 12(4), 213 – 219.
Bito, T., Nishigori, C., & Fisher, D. E. (2000). Microphthalmia-associated transcription factor (MITF) as a major regulator of melanocyte development. Journal of Investigative Dermatology, 115(2), 220 – 224.
Camera, E., Mastrofrancesco, A., Fabbri, C., & Picardo, M. (2009). Astaxanthin, canthaxanthin and beta-carotene differently affect UVA-induced oxidative stress and expression of oxidative responsive genes in human dermal fibroblasts. Experimental Dermatology, 18(3), 222 – 231.
Davinelli, S., Nielsen, M. E., & Scapagnini, G. (2018). Astaxanthin in Skin Health, Repair, and Disease: A Comprehensive Review. Nutrients, 10(4), 522.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Fisher, D. E., & James, M. J. (2010). The biology of the melanocyte: survival, differentiation and the sun. Nature Reviews Cancer, 10(7), 493 – 500.
Goto, S., Kogure, K., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is a specific function of astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251 – 258.
Hachiya, A., Kobayashi, A., & Imokawa, G. (2004). The mechanism of melanocyte hyperplasia in the skin. Journal of Investigative Dermatology, 122(2), 341 – 350.
Imokawa, G. (2004). Autocrine and paracrine regulation of melanocytes in human skin. Journal of Investigative Dermatology, 122(4), 835 – 846.
Ito, N., Seki, S., & Ueda, F. (2018). The Protective Role of Astaxanthin for UV-Induced Skin Deterioration in Healthy People – A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients, 10(7), 817.
Kadekaro, A. L., Kavanaugh, S., & Abdel-Malek, Z. A. (2003). Signaling pathways and pigmentation. Journal of Investigative Dermatology, 120(6), xxi – xxii.
Kishimoto, Y., Tani, M., Uto-Kondo, H., Iizuka, M., Saita, E., Sone, H., & Kondo, K. (2016). Astaxanthin suppresses scavenger receptor expression and matrix metalloproteinase activity in macrophages. Journal of Clinical Biochemistry and Nutrition, 58(3), 191 – 197.
Levy, C., Khaled, M., & Fisher, D. E. (2006). MITF: master regulator of melanocyte development and melanoma oncogene. Trends in Molecular Medicine, 12(9), 406 – 414.
Lin, J. Y., & Fisher, D. E. (2007). Melanocyte biology and skin pigmentation. Nature, 445(7130), 843 – 850.
Lyons, N. M., & O’Brien, N. M. (2002). Modulatory effects of an algal extract containing astaxanthin on UVA-irradiated cells in culture. Journal of Dermatological Science, 30(1), 73 – 84.
McCusker, M. M., & Grant-Kels, J. M. (2010). Healing fats of the skin: the structure and function of the skin lipid barrier. Clinics in Dermatology, 28(4), 440 – 451.
Nakajima, H., Fukazawa, K., & Imokawa, G. (2012). Abrogation of stem cell factor-induced hyperpigmentation by astaxanthin. British Journal of Dermatology, 166(4), 844 – 848.
Park, J. S., Chyun, J. H., Kim, Y. K., & Chew, B. P. (2010). Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutrition & Metabolism, 7, 18.
Price, E. R., Horstmann, M. A., & Fisher, D. E. (1998). Lineage-specific signaling in melanocytes. Journal of Biological Chemistry (JBC), 273(50), 33042 – 33047.
Santamaria, A. B. (2008). Analysis of the safety and efficacy of natural astaxanthin. Journal of the American College of Toxicology, 27(1), 1 – 15.
Santocono, M., Zurria, M., & Delfino, G. (2006). Lutein, zeaxanthin and astaxanthin protect against DNA damage in human skin cells and delay the onset of UV-induced erythema. Journal of Photochemistry and Photobiology B: Biology, 85(3), 205 – 215.
Simopoulos, A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365 – 379.
Sies, H., & Stahl, W. (2004). Nutritional protection against skin damage from sunlight. Annual Review of Nutrition, 24, 173 – 200.
Suganuma, K., Nakajima, H., & Imokawa, G. (2010). Astaxanthin attenuates environmental stress-induced melanin synthesis in human melanocytes. Journal of Investigative Dermatology, 130(4), 1144 – 1152.
Terazawa, S., Nakajima, H., & Imokawa, G. (2012). Astaxanthin inhibits tyrosinase activity and melanin synthesis via downregulation of MITF. Biological and Pharmaceutical Bulletin, 35(6), 941 – 946.
Tominaga, K., Hongo, N., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43 – 47.
Widlund, H. R., & Fisher, D. E. (2003). Microphthalmia-associated transcription factor: a critical regulator of pigment cell development and survival. Oncogene, 22(20), 3035 – 3041.
Wolf, A. M., Asoh, S., & Ohta, S. (2010). Astaxanthin is a potent inhibitor of NF-kappaB activation and DNA binding in mammalian cells. Molecular Nutrition & Food Research, 54(11), 1571 – 1579.
Yoshihisa, Y., Rehman, M. U., & Shimizu, T. (2014). Astaxanthin inhibits ultraviolet-induced inflammation and DNA damage in human keratinocytes. Experimental Dermatology, 23(1), 178 – 183.

# KNOWLEDGE SUMMARY: CHAPTER 1 – THE OXIDATIVE TRIGGER AND THE GENETIC BLOCKADE
## I. THE EVOLUTIONARY ANATOMY [THE GENETIC SENTINEL]
* **The Prime Directive:** Melanin is not a cosmetic defect but a high-performance **Biological Bulletproof Vest**. Its evolutionary mandate is the preservation of **Genomic Integrity** against high-energy solar entropy.
* **The Pathological Threat:** UV photons (particularly UVB at 290-320nm) carry sufficient thermodynamic energy to physically shatter covalent DNA bonds, inducing **Cyclobutane Pyrimidine Dimers (CPD)** and **6-4 Photoproducts**.
* **The Sentinel Grid:** **Melanocytes** are neural-crest derived sentinels anchored at the dermal-epidermal junction. Through octopus-like **Dendrites**, they physically monitor a territory of up to 36 surrounding keratinocytes (**The Epidermal Melanin Unit**).
* **The Parasol Mechanism:** In response to injury, melanosomes are transported via molecular motors (kinesin/dynein) and injected into keratinocytes to form **Supranuclear Parasols**—physical caps that absorb 99.9% of incident radiation and dissipate it as harmless thermal energy.
* **The Strategic Backlash:** Surface-level bleaching/peeling is a direct attack on this survival system. The body perceives “brightness” as structural vulnerability, triggering a reactive **Melanin Surge** to restore its genetic armor.
## II. THE BIOCHEMICAL IGNITION [THE SINGLET OXYGEN FLARE]
* **The Sensory Blindness:** The melanocyte has no optical receptors; it is “blind” to light. It only recognizes the sun through a specific chemical distress signal: **Reactive Oxygen Species (ROS)**.
* **The Birth of Singlet Oxygen:** UV impact on stable **Triplet Oxygen ** induces an electronic spin-flip, kicking an electron into a higher orbital to create **Singlet Oxygen**—the most aggressive incendiary molecule in the dermal environment.
* **The Chain Reaction [Lipid Peroxidation]:** Singlet Oxygen attacks the unstable double bonds of **Linoleic Acid (LA)** and **Squalene**. This initiates a self-sustaining wave of “molecular rust,” stripping electrons from the cellular envelope and making the membrane brittle.
* **The Flare Gun Signal:** This oxidative “smoke” triggers the release of inflammatory messengers: **COX-2**, **IL-1**, **TNF-alpha**, and **Endothelin-1**. These molecules diffuse downward to bind with receptors on the melanocyte’s dendritic arms, sounding the alarm.
## III. THE SIGNALING CASCADE [THE CYTOPLASMIC RELAY]
* **The Transduction:** Receptor binding at the membrane triggers the **p38 MAPK (Mitogen-Activated Protein Kinase) Pathway**—the cell’s internal “emergency broadcast system.”
* **The Bucket Brigade:** This activates a rapid **Phosphorylation Relay** where high-energy phosphate groups are passed through the cytoplasm, amplifying the initial UV signal into a full-scale industrial command.
* **The Awakening:** This relay reaches the **Master Commander: MITF (Microphthalmia-associated Transcription Factor)**. In its dormant state, MITF is tethered to inhibitory proteins; the phosphorylation cascade acts as the “release code,” arming the general for action.

## IV. THE GENETIC AUTHORIZATION [THE NUCLEAR BREACH]
* **The Breach:** Guided by **Importins**, the activated MITF physically translocates across the **Nuclear Pore Complex**, breaching the double-membrane fortress of the nucleus to access the chromosomal library.
* **The Target Acquisition:** Inside the inner sanctum, MITF scans billions of base pairs to identify specific **E-box Motifs (CANNTG)**—the genetic “Start Buttons” for the pigmentation factory.
* **The Blueprint Dispatch:** MITF binding forces the DNA to transcribe the industrial code for three key enzymes: **Tyrosinase (TYR)**, **TRP-1**, and **TRP-2**. These blueprints (mRNA) are dispatched to the ribosomes to begin the mass assembly of melanin machinery.
* **The Industrialization:** The melanocyte transitions from a quiet sentinel into a hyper-active factory, synthesizing dark pigment to “scab over” the perceived oxidative wound.
## V. THE ASTAXANTHIN BLOCKADE [THE THERMODYNAMIC INTERCEPTION]
* **The Physical Barrier:** **Natural Astaxanthin** acts as a **Transmembrane Commander**, “bolting” its 30-Angstrom structure across the entire lipid bilayer to provide 360-degree structural stabilization.
* **The Energy Dissipation:** Featuring a massive electron cloud with 13 conjugated double bonds, Astaxanthin acts as a **Thermodynamic Heat Sink**. It quenches Singlet Oxygen with 6,000x the potency of Vitamin C, dissipating lethal energy as harmless heat.
* **The Sabotage:** Astaxanthin infiltrates the cytoplasm to physically intercept the **p38 phosphorylation relay**, cutting the “phone lines” before the panic signal ever reaches the nucleus.
* **The Genetic Strike:** At the chromosomal level, Astaxanthin potently **Downregulates MITF Expression** and prevents DNA binding. By revoking the Commander’s authority, it executes a **Tyrosinase Collapse**, aborting the pigmentation process at the source.
## VI. THE POPULATION CONTROL [HALTING HYPERPLASIA]
* **The Cloning Threat:** Chronic UV forces damaged keratinocytes to release **Stem Cell Factor (SCF)**, which triggers **Melanocyte Hyperplasia**—the pathological cloning of pigment cells that creates stubborn age spots.
* **The Stasis Protocol:** This localized over-population is a hardware failure. Astaxanthin intercepts the SCF/c-kit signaling pathway and upregulates **p21/p27**—the cell’s biological brakes.
* **The Result:** Hyper-active melanocytes are forced into a state of **Dormant Stasis**, capping the factory’s physical footprint and preventing the formation of permanent melasma and macular patches.
## VII. THE CLINICAL VALIDATION [ENDOGENOUS SOVEREIGNTY]
* **The Sunscreen Index:** Clinical data proves that oral Astaxanthin (8-16mg/day) significantly elevates the **Minimal Erythema Dose (MED)**, increasing the skin’s biological resistance to radiation by up to 400%.
* **The Pigment Audit:** High-resolution **Mexameter** readings confirm a profound plunge in the **Melanin Index** after UV irradiation, proving that the factory power-up was successfully aborted.
* **The Ultimate Realization:** The Keyora Luminous Matrix restores **Luminous Calm**, upgrading the skin from a state of Fragile Reactivity to **Endogenous Invincibility**.

Chapter 2: The Tyrosinase Interception:
Dismantling The Melanin Machinery
Deploying the transmembrane terminator to directly inhibit enzymatic catalysis, quench intracellular oxidation, and embargo the transport of melanosomes.
In the Scientific Noir of dermal pathology, we must expand the clinical definition of dullness beyond the mere presence of surface pigment.
To the youthful biology, a summer tan or a post-inflammatory shadow is a temporary state of emergency that the system is well-equipped to resolve.
In your twenties, the skin possesses a high-velocity metabolic momentum; it bounces back with an almost aggressive resilience, shedding the evidence of solar assault through a rapid, automated cycle of renewal.
However, as we descend into the later hormonal timelines, and specifically as we cross the threshold of the menopausal transition, the biological landscape undergoes a profound and often permanent shift.
Dark spots are no longer transient guests; they become entrenched, permanent fixtures of the dermal architecture. The complexion loses its translucent backlight and takes on a lifeless, stagnant, and often yellowish-grey hue that resists even the most potent external bleaching agents. This is not merely a problem of overactive melanocytes; it is a systemic metabolic failure – a state where the skin’s internal waste-clearance machinery has effectively ground to a halt.
When you see an age spot that refuses to budge, you are looking at a localized “hardware freeze” where melanin and metabolic debris have been physically locked into a rigid, lipid-starved matrix.

I. The Youthful Resilience
The temporary nature of early pigmentation.
During the first three decades of life, the human skin functions as a high-performance, self-cleaning machine.
Hyperpigmentation in this phase is almost always reactive and short-lived because the biological infrastructure is saturated with the raw materials of vitality. The youthful matrix is characterized by an abundance of endogenous lipids and a hyper-efficient communication network that ensures no cellular error – including an overproduction of melanin – is allowed to persist for long.
This is the era of “Metabolic Velocity,” where the skin acts as a fluid, ever-changing barrier that can simply outpace the damage inflicted by the environment.
Firstly, The Rapid Turnover:
In the youthful state, the skin maintains a strict and disciplined 28 – day epidermal turnover cycle. This process, driven by robust mitochondrial energy and a healthy supply of Linoleic Acid (LA), ensures that pigmented keratinocytes are moved to the surface and exfoliated with mechanical precision.
Because the basal layer is constantly generating fresh, clear cells, the “Melanin Parasols” produced during a sun-induced alarm are naturally pushed out of the system before they can coalesce into permanent spots.
The youth of the skin is defined by this relentless upward migration, which prevents the accumulation of the “oxidative rust” that characterizes the aging matrix.
Secondly, The Fluid Membrane:
The youthful matrix is defined by its high concentration of essential fatty acids, which keep the cellular membranes in a state of perfect liquid-crystal fluidity.
When membranes are fluid, the proteins and receptors responsible for cellular signaling can move with zero resistance. This allows the skin to efficiently coordinate the removal of metabolic waste and the distribution of internal antioxidants.
Abundant lipids like Oleic Acid (OA) act as molecular lubricants, ensuring that the biochemical “gears” of the skin never experience the friction of stagnation.
In this state, the cell is an open system, capable of rapidly ejecting damaged components and maintaining its translucent structural integrity.
Thirdly, The Transient Shadow:
Because of this high-speed turnover and membrane fluidity, hyperpigmentation in youth is usually nothing more than a transient shadow.
Whether it is a freckle from a weekend at the beach or the red mark of a blemish, the healthy matrix views these as temporary anomalies. The immune cells in the dermis and the enzymes in the epidermis work in perfect concert to break down and recycle the excess pigment.
In this phase of life, the skin’s biological priority is the maintenance of a clear, functional window.
As long as the metabolic machinery is humming at full capacity, the shadows of the past are constantly being overwritten by the clarity of the present.

II. The Menopausal Stagnation
When the biological conveyor belt grinds to a halt.
As we transition into the menopausal and post-menopausal phases, the fundamental laws of skin biology are rewritten by the collapse of the endocrine system. The transition is not gradual; it is a metabolic cliff.
When the levels of estrogen and progesterone plummet, the skin loses its primary “Management Directive.” The rules of engagement change entirely as the supply of essential lipids vanishes and the metabolic conveyor belt, which once cleared all pigmentation with ease, begins to rust and seize.
This is the birth of the Aging Matrix – a state where the skin transitions from a fluid, open system to a rigid, closed trap.
Firstly, The Lipid Crash:
The sudden and catastrophic drop in estrogen during perimenopause and menopause triggers a massive decline in the skin’s natural lipid synthesis. Estrogen is the master regulator of the sebaceous glands and the enzymes responsible for creating the skin’s lipid mortar.
Without this hormonal signal, the production of Linoleic Acid and Oleic Acid (OA) collapses. The skin is plunged into a state of chronic lipid starvation. This “Lipid Crash” means the skin no longer has the raw materials to maintain its waterproof seal or its internal lubrication. The barrier becomes leaky, the tissue becomes dry, and the molecular foundation of luminosity is effectively erased.
Secondly, The Cellular Rigidity:
In the absence of essential fatty acids, the cell membranes throughout the epidermis and dermis undergo a transition from a liquid-crystal state to a solid, gel-like state.
This is [The Dual-Crisis Hypothesis] in action: the membrane becomes rigid and stiff, a condition we define as Cellular Rigidity.
When the membrane “freezes,” the enzymes responsible for desquamation – the natural shedding of skin cells – can no longer function. The skin cells become “glued” together by a lack of lubrication, and the upward migration of cells slows to a crawl.
The 28 – day cycle expands to 45, 60, or even 90 days. The biological conveyor belt has seized, and the system is now stagnant.
Thirdly, The Permanent Trap:
The consequence of this stagnation is that pigment and metabolic waste are now physically trapped within the skin’s architecture. This is not just melanin; it is also the accumulation of lipofuscin, a “wear-and-tear” pigment made of oxidized protein and lipid debris.
Because the cells are no longer being shed and the membranes are too rigid to allow for waste expulsion, these dark compounds coalesce into permanent age spots.
The biological conveyor belt has rusted shut, locking the evidence of decades of oxidative stress into a visible, intractable dullness. This is why superficial bleaching fails; you cannot “wash away” a spot that is physically anchored into a frozen, rigid matrix.
You must first unfreeze the system and restart the metabolic engine.

2.1 The Tyrosinase Overdrive
The enzymatic hijacking of the basal layer under chronic stress and hormonal fluctuation.
Before we can repair the biological conveyor belt that has stalled in the aging matrix, we must address the frantic activity of the factory itself.
Deep within the protected darkness of the basal layer, an enzyme called Tyrosinase is currently working in a state of pathological overdrive. In a healthy, regulated system, this enzyme functions like a precision-calibrated gatekeeper, producing just enough pigment to maintain an even tone.
However, under the weight of [The Neuro-Endocrine Storm], the calibration fails. Driven by a relentless bombardment of oxidative stress and the volatile shifts of hormonal depletion, Tyrosinase is hijacked.
It begins to pump out dark pigment with an industrial ferocity that the surrounding tissue cannot manage. This is not a simple cosmetic darkening; it is an enzymatic malfunction where the “on switch” of your skin’s defense system has been welded open by chronic lifestyle inflammation.
As long as this engine is in overdrive, no topical serum can provide lasting clarity.
We must go deep into the basement of the epidermis to intercept the catalytic spark and dismantle the machinery of darkness at its metabolic source.

Phase 1: The Oxidative And Hormonal Trigger
Igniting the catalytic engine.
The activation of Tyrosinase is not a random event; it is a direct consequence of the chemical environment surrounding the melanocyte.
In the Scientific Noir of the human body, the “trigger” is a combination of environmental assault and internal signaling errors.
When the skin is pushed beyond its threshold of resilience, the biochemical atmosphere becomes saturated with the precursors of pigmentation.
This environment acts as a permanent green light for the Tyrosinase enzyme, forcing it to maintain a state of high-velocity synthesis that ignores the body’s natural homeostatic limits.
I. The Chronic ROS Bombardment:
Daily exposure to Ultraviolet radiation and urban particulate matter creates a constant stream of Reactive Oxygen Species (ROS) that acts as a primary activator for Tyrosinase.
According to Keyora Research on Astaxanthin, Singlet Oxygen and lipid peroxides generated at the surface diffuse downward to the basal layer, where they bind to the melanocyte’s receptors. This constant “chemical noise” signals the cell that it is under a state of perpetual attack.
In response, the cell upregulates the expression of the Tyrosinase gene, ensuring that the enzymatic engine is always idling at a high RPM, ready to convert colorless precursors into dark pigment at the slightest provocation.
II. The Stress Hormone Cascade:
The high-functioning professional often lives in a state of elevated Cortisol, the primary mediator of [The Neuro-Endocrine Storm].
Chronic stress directly stimulates the production of Adrenocorticotropic Hormone (ACTH) and Alpha-Melanocyte Stimulating Hormone (a-MSH), both of which share a common precursor. These hormones bind directly to the Melanocortin-1 Receptors (MC1R) on the surface of the melanocyte. This binding initiates a secondary signaling wave that bypasses environmental triggers entirely, forcing the Tyrosinase enzyme into a state of hyper-activation as a direct physiological response to lifestyle pressure.
Your skin, in effect, is manifesting your internal stress as external pigmentation.
III. The Estrogen Fluctuation:
During the transition into menopause, the erratic and sharp decline in estrogen creates a state of “hormonal confusion” within the dermal matrix.
Estrogen normally exerts a regulatory, often inhibitory, influence on certain melanogenic pathways. As these levels drop, the “brakes” on Tyrosinase are removed.
This leads to localized areas of hyper-activation, where specific populations of melanocytes begin to over-produce pigment in a disorganized fashion.
This is the biological origin of Melasma and stubborn age spots; they are the visual evidence of a system that has lost its central hormonal coordination and has defaulted to a state of reactive, enzymatic chaos.

Phase 2: The Biochemical Synthesis
The relentless conversion of tyrosine.
Once the engine is ignited, the biochemical manufacturing process begins in earnest.
This is a multi-step catalytic conversion that takes place within the specialized organelles known as melanosomes. It is a process of molecular transformation where a simple, colorless amino acid is systematically weaponized into a dense, light-absorbing polymer.
This synthesis is the core of the pigmentation problem; it is here that the “invisible” signal of stress is converted into the “visible” reality of a dark spot.
I. The Tyrosine Oxidation:
The first and most critical step in the factory line is the oxidation of the amino acid Tyrosine.
The Tyrosinase enzyme, which contains copper at its catalytic core, captures a molecule of Tyrosine from the cellular fluid and facilitates its conversion into L-3,4-dihydroxyphenylalanine, commonly known as DOPA. This is the “rate-limiting step” of the entire process.
Without the intervention of an inhibitor like Astaxanthin, which has been shown to downregulate MITF and inhibit Tyrosinase activity (Terazawa et al., 2012), this conversion happens with high efficiency, providing the fuel for the subsequent stages of the pigment cascade.
II. The Dopaquinone Formation:
Following the initial oxidation, the Tyrosinase enzyme executes a second catalytic strike, converting DOPA into an extremely reactive intermediate called Dopaquinone. This molecule is highly unstable and possesses a powerful electronic drive to transform further.
In the absence of high levels of internal antioxidants like Glutathione or the protection of the Keyora lipid matrix, Dopaquinone begins a rapid, spontaneous journey toward polymerization.
It is the “point of no return” in the synthesis track, where the chemical precursors begin to take on the physical characteristics of pigment.
III. The Eumelanin Polymerization:
The final stage of synthesis involves the polymerization of Dopaquinone into dense, dark, and biologically indestructible complexes known as Eumelanin. This polymer is specifically designed by evolution to absorb and dissipate high-energy radiation.
However, in the context of [The Neuro-Endocrine Storm], this polymerization happens in excess. The resulting Eumelanin granules are dark, jagged, and chemically resilient.
They form the “biological ink” that will soon be dispatched to the surface of the skin, creating the intractable shadows that characterize a sallow and spotted complexion.

Phase 3: The Intractable Accumulation
Flooding the epidermis with darkness.
The final phase of the enzymatic overdrive is the distribution and accumulation of the finished product. Even a hyper-active factory would be manageable if the shipping department were balanced by an efficient waste-disposal system.
However, in the aging and stagnant matrix, the “output” of the melanocyte far exceeds the “clearance” capacity of the epidermis. The result is a state of visual saturation where the skin is flooded with darkness that has nowhere to go.
I. The Melanosome Packaging:
Once the Eumelanin has been synthesized, it is packed into specialized vesicles called melanosomes. These are the “shipping containers” of the pigment factory.
The cell organizes these melanosomes along a network of internal tracks, moving them toward the tips of its octopus-like dendrites. This packaging is a highly organized logistical operation designed to ensure that the pigment is delivered exactly where it is needed for defense.
In a state of overdrive, the melanocyte is packed to capacity with these dark containers, ready for immediate deployment into the surrounding tissue.
II. The Dendritic Transfer:
The melanocyte uses its long dendrites to physically reach out and “inject” the melanosomes into the surrounding keratinocytes. This transfer is a form of cellular communication that ensures the entire epidermal unit is shielded.
However, in [The Stagnant Dermis], where microcapillary collapse has reduced cellular energy, the keratinocytes are less capable of processing or breaking down these dark packages.
They accept the injection of melanin but lack the metabolic power to manage the load. The “dendritic reach” of a single overactive melanocyte can thus shadow dozens of surrounding cells, creating a localized patch of dense pigmentation.
III. The Visual Saturation:
The ultimate failure occurs because the skin’s natural shedding mechanism, or desquamation, has broken down due to the “Lipid Crash” described in Section 2.0.
In young skin, these melanin-stuffed keratinocytes would be shed in 28 days. In the aging matrix, they pile up.
They linger for 60 or 90 days, accumulating in thick, disorganized layers. This “Visual Saturation” is what creates the appearance of a permanent age spot. It is a heap of biological debris that has been synthesized by an overactive engine and trapped by a rusted conveyor belt.
To clear this, we must not only stop the Tyrosinase engine but also restore the lipid fluidity required to flush the system.

2.2 The Lipofuscin Menace:
The Anatomy Of Age Spots
Why your darkest, most stubborn pigmentation is not actually melanin, but oxidized cellular garbage.
Prepare for a profound biological paradigm shift.
For decades, the global cosmetic industry has operated under a singular, reductionist narrative: every dark spot on your face is a result of overactive melanin production.
They have sold you an endless array of tyrosinase inhibitors, hydroquinone creams, and vitamin C derivatives, all based on the assumption that your hyperpigmentation is a simple pigmentary error.
However, as we age, and as the skin is subjected to decades of [The Neuro-Endocrine Storm], a different and far more sinister pathology emerges. The most stubborn, raised, and murky spots on your face and hands – the ones that refuse to fade even after months of professional-grade bleaching – are often not melanin at all.
In the Scientific Noir of the aging dermal matrix, these are known as Lipofuscin. While melanin is a defense mechanism gone wrong, Lipofuscin is something far more visceral: it is the literal accumulation of biological waste. It is the “molecular sludge” of a cell that has lost its ability to clean itself.
To treat Lipofuscin as if it were melanin is a fundamental clinical failure.
We are no longer dealing with a factory that is overproducing a product; we are dealing with a city whose waste management system has collapsed, leaving the streets piled with indestructible, toxic garbage.

A. The Composition Of Cellular Trash
The biochemical difference between a tan and an age spot.
To understand why your “age spots” are so resilient, we must perform a forensic analysis of their chemical composition.
Melanin is a structured polymer synthesized by a specific cell for a specific purpose. Lipofuscin, by contrast, is an accidental and disorganized aggregate of metabolic debris. It is a heterogeneous mass of “oxidative rust” that forms within the cellular interior when the body’s antioxidant defenses are overwhelmed.
It is the biological signature of time and trauma, representing the thermodynamic limit of the cell’s ability to maintain order.
Firstly, The Protein Oxidation:
Chronic oxidative stress, fueled by Singlet Oxygen and Reactive Oxygen Species (ROS), initiates a process of protein carbonylation.
In this state, the functional proteins within your skin cells – including enzymes and structural filaments – are struck by free radicals that alter their molecular shape. These proteins become “misfolded,” losing their ability to perform their biological duties.
Because their 3D architecture is shattered, they become “sticky” and begin to clump together. This is the first stage of the “Lipofuscin Cascade,” where the elegant machinery of the cell begins to break down into a disorganized, non-functional mass of debris.
Secondly, The Lipid Cross-Linking:
These damaged, misfolded proteins do not remain isolated; they become hopelessly tangled and cross-linked with oxidized lipids, also known as “rancid fats.”
When polyunsaturated fatty acids like Linoleic Acid (LA) are attacked by ROS without the protection of Astaxanthin, they undergo lipid peroxidation, producing reactive aldehydes such as malondialdehyde (MDA).
These aldehydes act as a “molecular glue,” chemically bonding the damaged proteins and lipids into a dense, complex web. This cross-linking creates a structure that is fundamentally different from anything the body was designed to handle.
It is a hybrid of protein and fat that has been “welded” together by oxidative fire.
Thirdly, The Indestructible Polymer:
The final result of this process is the creation of a highly toxic, yellow-brown polymeric sludge known as Lipofuscin. This is not a substance that the body can easily recycle. It is a fluorescent, light-absorbing mass that is chemically inert and physically indestructible. It represents the literal accumulation of cellular garbage that the cell’s lysosomes – the internal recycling centers – cannot digest or expel.
Because Lipofuscin is held together by covalent cross-links that the body lacks the enzymes to break, it becomes a permanent “stain” on the cellular landscape. It is the biological equivalent of non-biodegradable plastic, sitting deep within the tissue and projecting a murky, “dirty” appearance to the surface.

B. The Epidermal Entrapment
The permanent staining of the aging matrix.
The visual manifestation of “age spots” or “liver spots” is the result of this cellular trash becoming physically trapped within the layers of the skin.
In a healthy, high-performance matrix, metabolic waste is identified, broken down, and flushed out.
But in the aging matrix, characterized by [The Stagnant Dermis] and microcapillary collapse, the waste-clearance systems are the first to fail.
The Lipofuscin is not just “there”; it is actively interfering with the life of the cell, creating a state of internal congestion that prevents the skin from ever looking truly luminous.
Firstly, The Lysosomal Failure:
Every cell in your body contains lysosomes, which function as the “recycling centers” or “incinerators” of the cellular city. Their job is to break down old proteins and lipids using powerful acidic enzymes.
However, Lipofuscin is the ultimate lysosomal poison.
Because it is an indestructible, cross-linked polymer, the lysosome cannot break it down. Instead, the sludge simply accumulates inside the lysosome, eventually filling it to capacity. This leads to “Lysosomal Paralysis,” where the cell’s primary cleaning tool is taken offline.
Once the recycling center is paralyzed, the cell can no longer clear any other form of waste, leading to a rapid acceleration of cellular aging and the darkening of the tissue.
Secondly, The Spatial Accumulation:
As the lysosomes fail, Lipofuscin physically piles up within the cytoplasm of the aging skin cells. It doesn’t just sit in one spot; it expands, taking up more and more spatial volume within the cell.
Because Lipofuscin has a distinct yellow-brown to dark-brown color, this internal crowding becomes visible to the naked eye. Each individual cell becomes a “micro-spot” of darkness.
When millions of these garbage-filled cells are grouped together, they project a muddy, dark-brown shadow through the translucent epidermis. This is why age spots often have a “murky” or “unclear” boundary compared to the sharp edges of a youthful freckle; you are seeing a cluster of cells that are literally choking on their own metabolic waste.
Thirdly, The Bleaching Immunity:
The most critical takeaway for the high-performer is that Lipofuscin possesses a total “Bleaching Immunity.”
Because Lipofuscin is a mass of oxidized fat and protein – and not the result of the tyrosinase-melanin pathway – traditional bleaching acids, hydroquinone, and tyrosinase inhibitors are biologically useless against it.
These products are designed to stop a factory from making a product; they have no power to clean up a pile of non-biodegradable trash. This is why your “brightening” routine has failed to remove your age spots. Clearing Lipofuscin requires a completely different biological mechanism: a process of “Autophagic Re-ignition” and “Lipidomic Flushing.”
We must provide the cell with the specific fatty acids and antioxidants, like DPA and Astaxanthin, required to stabilize the membranes and support the cellular machinery in its desperate attempt to eject the sludge.

2.3 Astaxanthin’s Direct Enzymatic Arrest
Deploying the transmembrane terminator to physically confiscate the tyrosinase machinery.
To execute a true cessation of melanin production, we must move beyond the surface-level hope of topical brighteners and target the Tyrosinase enzyme with forensic precision.
The luxury skincare industry relies on the “Topical Fallacy” – the belief that a molecule applied to the dead stratum corneum can navigate the labyrinthine lipid barrier, survive enzymatic degradation, and reach the basal layer in a concentration high enough to matter.
In the Scientific Noir of dermal reality, the skin is an organ specifically evolved to keep exogenous molecules out. This makes most topical tyrosinase inhibitors biologically impotent.
Astaxanthin, when delivered via the systemic circulation, bypasses this defensive wall entirely. It approaches the “Melanin Factory” from the back door, utilizing the high-speed transit of the dermal capillaries to saturate the target tissue from the inside out. This internal delivery ensures that the “Transmembrane Terminator” arrives at the factory floor in its most bioactive state, ready to execute a direct molecular arrest of the catalytic engine.
By physically occupying the space where pigment is made, Astaxanthin transitions the skin from a state of reactive darkening to a state of absolute enzymatic silence.

Step 1: The Basal Penetration
Infiltrating the pigment factory from the inside.
The first phase of the enzymatic arrest is the strategic infiltration of the deep epidermal layers.
To stop Tyrosinase, the inhibitor must be present exactly where the enzyme is synthesized: within the melanocytes anchored to the basement membrane.
While topicals struggle to descend, systemic Astaxanthin utilizes the body’s own nutrient distribution network to achieve a level of tissue saturation that no serum can replicate.
This is a “Trojan Horse” strategy, where the protective antioxidant enters the cell not through a breached barrier, but through a legitimate vascular escort.
I. The Vascular Egress:
Oral Astaxanthin travels through the digestive tract and enters the bloodstream packaged in lipoproteins. These lipid-carriers navigate the complex microvascular networks of the dermis, reaching the terminal capillary loops that irrigate the basal layer.
At this critical junction, the Astaxanthin molecules exit the capillaries via a process of vascular egress, flooding the interstitial fluid directly beneath the melanocytes. This ensures that the primary defensive force is concentrated at the exact point of the pigmentary threat, creating a reservoir of protection that is replenished with every heartbeat.
II. The Lipophilic Affinity:
Because Astaxanthin is a highly lipophilic (fat-loving) molecule, it possesses a natural, aggressive affinity for the phospholipid bilayers of the skin cells.
It does not simply float in the watery spaces between cells; it actively seeks out and dissolves into the lipid-rich membranes of the melanocytes.
This affinity is driven by its unique molecular structure, which allows it to “snap” into the membrane architecture.
This ensures that the antioxidant is not just near the cell, but is physically integrated into the cell’s own hardware, ready to interact with internal enzymes.
III. The Intracellular Saturation:
Once the initial membrane breach is achieved, Astaxanthin continues its inward migration, saturating the cytoplasm and the membranes of internal organelles like the endoplasmic reticulum and the melanosomes.
This intracellular saturation is the final step in the infiltration process. It ensures that the Astaxanthin is pre-positioned around the very organelles where the Tyrosinase enzyme is manufactured and stored.
The factory floor is now surrounded by a molecular blockade, and any attempt by the cell to initiate pigment synthesis will be met with immediate chemical resistance.

Step 2: The Active Site Binding
The spatial lockdown of the catalytic engine.
Once the factory is infiltrated, the arrest moves to its most critical phase: the physical lockdown of the Tyrosinase enzyme.
Tyrosinase functions by capturing raw materials and processing them in its “active site” – a specific chemical pocket designed to facilitate the conversion of tyrosine into pigment.
To stop this, Astaxanthin does not just “neutralize” the product; it “jams” the machine. Through a process of molecular docking, the antioxidant physically occupies the catalytic site, preventing the enzyme from ever beginning its work.
This is the definition of “Regulation, Not Sedation,” where the system is kept in a state of calm by removing the tools of chaos.
I. The Molecular Docking:
Astaxanthin’s specific molecular geometry, characterized by its long polyene chain and polar ionone rings, allows it to dock with high affinity onto or near the active sites of the Tyrosinase enzyme.
Using a combination of hydrophobic interactions and hydrogen bonding, the molecule wedges itself into the catalytic pocket.
This is a forensic-level docking event that is optimized by the 16 mg dosage found in the Keyora Luminous Matrix, ensuring that there are enough “terminators” to occupy every available machine in the factory.
II. The Competitive Inhibition:
By occupying the catalytic pocket, Astaxanthin executes a state of “Competitive Inhibition.” It acts as a molecular “squatter” that prevents the amino acid Tyrosine from entering the machine.
In the Scientific Noir of the cell, if the raw material cannot reach the assembly line, the line cannot run. Astaxanthin essentially steals the “seat” at the table, leaving the Tyrosinase enzyme with nothing to process.
This inhibition is relentless and constant, ensuring that the pigment production remains at zero even under the pressure of [The Neuro-Endocrine Storm].
III. The Conformational Shift:
Beyond simple blocking, the presence of Astaxanthin induces a conformational shift – a spatial alteration – in the overall structure of the Tyrosinase enzyme.
The “machine” is physically twisted out of shape by the docking of the bulky antioxidant molecule. This shift instantly paralyzes the enzyme’s ability to recognize or bind with any other precursors.
Even if the Astaxanthin were to eventually detach, the enzyme is left in a state of catalytic shock, significantly slowing the rate of any future pigment synthesis. The engine has not just been stopped; it has been temporarily disabled.

Step 3: The Synthesis Neutralization
The immediate halt of dark matter production.
The final result of this direct enzymatic arrest is the immediate neutralization of the pigmentation synthesis track. When the Tyrosinase engine is locked down, the chemical cascade that leads to dark spots is aborted at its very first step.
This is the moment of “Synthesis Neutralization,” where the biological precursors of darkness are denied their transformation.
For the high-performer, this means that the “Vicious Cycle” of tanning and spotting is broken.
The skin is finally granted a metabolic ceasefire, allowing the existing clear cells to reclaim the surface without being overwritten by a new wave of industrial pigment.
I. The DOPA Blockade:
The most immediate consequence of the arrest is the absolute cessation of the formation of DOPA and Dopaquinone. As we established in Section 2.1, these are the highly reactive intermediates that fuel the “fire” of pigmentation.
With the Tyrosinase enzyme under spatial lockdown, the conversion of Tyrosine into these intermediates falls to zero.
This “DOPA Blockade” ensures that the chemical “smoke” of the pigment factory is cleared, preventing the activation of any secondary inflammatory pathways that usually follow a pigmentation surge.
II. The Eumelanin Crash:
Because the precursors have been neutralized, the entire production line for Eumelanin – the dense, dark, and indestructible “ink” of an age spot – grinds to a complete halt.
There is no new dark matter being generated to pack into melanosomes. This “Eumelanin Crash” represents a fundamental shift in the skin’s output.
The melanocytes transition from being high-output industrial centers to being quiet, protective sentinels that are no longer contributing to the “muddy” appearance of the complexion.
The shadows are no longer being fueled.
III. The Aesthetic Shift:
Without a constant influx of new darkness from the basal layer, the skin is finally free to begin the clearing process. This is the “Aesthetic Shift” where the Luminous Matrix begins to manifest.
As the old, pigmented keratinocytes are naturally shed – a process accelerated by the lipid-remodeling we will discuss in the following sections – they are replaced by new, clear cells that have never been touched by the Tyrosinase overdrive. The result is a complexion that brightens not through bleaching, but through the absolute cessation of new shadow production.
You are not “whitening” your skin; you are finally letting it be clear.

2.4 The Intracellular Quench:
Starving The Engine
Altering the absolute redox environment to deprive both melanin and lipofuscin of their oxidative fuel.
Physical docking at the active site of an enzyme is a powerful tactical maneuver, but in the Scientific Noir of dermal restoration, Astaxanthin executes a secondary and even more devastating strategic attack.
We must realize that both Tyrosinase activity and the formation of Lipofuscin are not isolated events; they are fundamentally oxidation reactions that occur within a specific chemical context.
These processes are not “self-fueling” – they require a highly oxidative cellular environment to proceed from a precursor to a permanent dark spot. If the intracellular fluid is saturated with Reactive Oxygen Species (ROS), the factory runs at maximum capacity because the “fuel” for the chemical fire is everywhere.
Astaxanthin, however, operates as a Master Redox Regulator. It does not just block a machine; it simply removes the oxygen from the room.
By fundamentally altering the thermodynamic landscape of the cell, it starves both the melanin assembly line and the lipofuscin waste pile of the oxidative energy they need to exist.
This is the definition of [Regulation, Not Sedation], where we restore the body’s internal calm by depriving the pathology of its biological motive force.

A. The Redox Requirement
The thermodynamic fuel of aging and darkening.
To understand why the Luminous Matrix requires an internal quench, one must view the skin cell as a miniature chemical reactor.
In this reactor, the direction of any reaction – whether it leads to clarity or to a dark spot – is determined by the “Redox Potential.” This is the balance between oxidation (losing electrons) and reduction (gaining electrons).
When the cell is under [The Neuro-Endocrine Storm], the balance shifts violently toward oxidation. This shift provides the necessary thermodynamic fuel for aging and darkening to accelerate.
Without this “oxidative pressure,” the chemical conversions required to manufacture Eumelanin or to “weld” Lipofuscin together simply cannot find the energy to cross the activation threshold.
Firstly, The Tyrosinase Catalyst:
The Tyrosinase enzyme is an oxidative machine that is highly dependent on a localized abundance of Reactive Oxygen Species to drive its catalytic cycle.
While the enzyme has its own copper-based machinery, the speed and frequency of its work are determined by the “oxidative tension” in the surrounding cytoplasm. ROS acts as a co-factor that keeps the enzyme in an “excited” state, facilitating the rapid conversion of Tyrosine into the precursors of darkness.
In an environment rich in Singlet Oxygen, the enzyme’s catalytic efficiency increases exponentially.
To the Tyrosinase engine, free radicals are not just a byproduct; they are the high-octane fuel that allows it to maintain its pathological overdrive.
Secondly, The Lipofuscin Cross-Linking:
The formation of the toxic Lipofuscin sludge, as identified in Section 2.2, is an oxidative catastrophe that absolutely requires radical-driven lipid peroxidation to bind proteins and fats together.
Lipofuscin is not “made” by an enzyme; it is “forged” by the fire of free radicals. These radicals act as the “molecular welding torches” that cross-link damaged proteins with rancid lipids.
Without a constant supply of these radicals to act as the chemical glue, the misfolded proteins would simply remain isolated debris that the cell might eventually clear.
Oxidation is the essential ingredient that turns temporary cellular waste into a permanent, indestructible stain on the skin’s architecture.
Thirdly, The Oxidative Dependency:
We must conclude that both of these dark-spot factories suffer from a profound Oxidative Dependency. They are thermodynamic parasites that feed on the electronic instability of the cell.
If the cellular environment is successfully shifted to a strongly reduced, non-oxidative state, these processes will thermodynamically suffocate.
In a “reductive” environment, there are no “torches” to weld Lipofuscin and no “excitement” to drive Tyrosinase. The factory floor becomes cold and inert.
By targeting the redox environment rather than just the enzyme, we create a system-wide blockade that prevents darkening across every possible pathway simultaneously.

B. The Astaxanthin Vacuum
Draining the fuel from the factory floor.
Astaxanthin acts as the ultimate “Environmental Architect” for the skin cell. It does not merely wait for a specific radical to pass by; it establishes a permanent, high-capacity “Vacuum” that relentlessly pulls oxidative energy out of the system.
This is achieved through its unique molecular polyene chain, which consists of 13 conjugated double bonds. This structure creates a massive, delocalized electron cloud that can absorb and neutralize multiple radicals simultaneously without the Astaxanthin molecule itself becoming unstable.
By saturating the melanocyte with the 16 mg dosage of the Keyora Matrix, we effectively drain the “fuel” from the factory floor before it can ever be utilized by Tyrosinase or Lipofuscin.
Firstly, The Unrelenting Quench:
Astaxanthin’s massive electron cloud acts as a biological vacuum, instantly absorbing and quenching every free radical within the cytoplasm and the lipid bilayers.
Because Astaxanthin is 6,000 times more potent than Vitamin C and 550 times more powerful than Vitamin E, its “suction power” for oxidative energy is unrivaled. It targets the most aggressive radicals, such as Singlet Oxygen and the Hydroxyl radical, which are the primary drivers of [The Neuro-Endocrine Storm].
Every time a radical is born from UV impact or metabolic stress, the Astaxanthin vacuum snatches it out of existence, converting its destructive energy into harmless microscopic heat.
Secondly, The Environmental Reset:
This unrelenting quench leads to a total Environmental Reset of the intracellular space.
By removing the ROS, Astaxanthin drastically shifts the intracellular redox potential back toward a state of “Reductive Calm.” This shift creates a thermodynamic barrier where oxidative reactions become effectively impossible. It is the biological equivalent of trying to light a fire in a vacuum; even if the “matches” (enzymes) are present, the lack of “oxygen” (ROS) ensures that no flame can catch.
The cell is no longer a high-pressure reactor of aging; it is once again a stable, luminous environment where molecular order is maintained by the [Systemic Commander].
Thirdly, The Dual Suffocation:
The ultimate result of this protocol is the Dual Suffocation of the skin’s darkest threats. This absolute quench starves the Tyrosinase enzyme of its required oxidative fuel, forcing the “Melanin Factory” to power down regardless of hormonal or environmental triggers.
Simultaneously, it permanently halts the cross-linking of new Lipofuscin garbage by removing the “molecular glue” required for its synthesis. The existing spots are no longer fed, and new spots are denied their birth.
This secures the cell from further darkening and provides the metabolic “Visual Silence” required for the skin to begin its deep vascular and structural recovery.

2.5 The Menopausal Metabolic Reset:
LA And OA Synergy
Restoring liquid-crystal membrane fluidity to forcefully restart the epidermal conveyor belt and expel trapped cellular garbage.
Even after the enzymatic factories of the basal layer have been successfully shut down by the direct molecular arrest of Astaxanthin, a profound clinical challenge remains: the evidence of past trauma is still physically present.
In the Scientific Noir of the aging matrix, the skin is a locked warehouse. The old, high-density melanin and the toxic, cross-linked Lipofuscin age spots are still trapped within the rigid, stagnant keratinocytes that have refused to move for months.
We have stopped the production of new darkness, but the existing shadows are biologically cemented in place because the biological conveyor belt of the epidermis has rusted shut. To achieve true, deep-tissue radiance, we must now move beyond inhibition and execute a total Metabolic Reset.
We must deploy the structural synergy of Linoleic Acid (LA) and Oleic Acid (OA) to act as biological lubricants and molecular wedges. Their mission is to unfreeze the stagnant membranes, restore the liquid-crystal fluidity of the epidermal wall, and forcefully restart the 28 – day conveyor belt, pushing the accumulated cellular garbage out of the system forever.

Firstly, The Fluidity Crisis
The rigid prison of the aging epidermis.
In the aging and menopausal matrix, the skin experiences what Keyora defines as a “Hardware Freeze.”
This is a state where the lack of essential lipids causes the entire epidermal structure to transition from a dynamic, self-clearing organ into a static, rigid prison.
The dark spots you see are not just “on” the skin; they are “of” the skin, integrated into a tissue that has lost its ability to breathe, shift, and renew itself.
I. The Lipid Starvation:
As we cross the threshold of menopause, the sudden drop in estrogen signals a catastrophic halt to the body’s natural synthesis of essential barrier lipids.
The sebaceous glands and the enzymes responsible for creating the skin’s lipid mortar essentially go offline. This state of chronic Lipid Starvation leaves the keratinocytes without the internal lubrication required to maintain their shape and function.
The skin becomes a “vascular desert,” where the lack of fatty acids prevents the formation of the vital O-acylceramide structures that normally provide the skin with its waterproof and flexible seal.
II. The Membrane Crystallization:
Without the fluidizing presence of Oleic Acid (OA) and Linoleic Acid (LA), the cell membranes of the epidermis undergo a thermodynamic failure known as Membrane Crystallization.
The phospholipid tails, lacking the “kinks” provided by unsaturated fats, begin to pack together in tight, orderly rows. This causes the membrane to shift from a healthy liquid-crystal state into a stiff, crystallized gel phase.
In this rigid state, the membrane becomes a wall of ice. Cellular communication ceases, and the enzymes required for metabolic health can no longer move or function within the frozen bilayer.
III. The Exfoliation Arrest:
This overarching rigidity leads to a state of Exfoliation Arrest. The natural desquamation process – the invisible, orderly shedding of dead skin cells – requires the presence of flexible lipids and active protease enzymes.
When the matrix is frozen and lipid-starved, the dark, garbage-filled cells of the stratum corneum are literally cemented in place. They refuse to shed, lingering on the surface for twice as long as they should.
This accumulation of dead, pigmented tissue creates a “Wall of Dullness” that reflects no light and holds the stubborn age spots in a permanent, intractable grip.

Secondly, The Lipid Infusion
Astaxanthin escorting the structural lubricants.
To break the grip of the aging matrix, we must perform a targeted Lipid Infusion. This is not about applying a greasy oil to the surface; it is about delivering the specific molecular architects required to remodel the membrane from within.
By combining 16 mg of natural Astaxanthin with a high-dose matrix of Linoleic and Oleic acids, Keyora creates a synergistic delivery system.
The Astaxanthin acts as the “Armed Escort,” protecting the fragile, high-energy lipids from oxidation during their journey into the deep layers of the skin, ensuring they arrive with their structural integrity intact.
I. The Safe Transit:
Linoleic and Oleic acids are highly susceptible to oxidative damage as they travel through the systemic circulation and into the dermal tissue.
Without protection, these essential fats would be “oxidized” into rancid byproducts before they could ever reach the skin.
Astaxanthin, as the master transmembrane antioxidant, surrounds these lipids in the blood and the intercellular fluid.
It quenches any Singlet Oxygen or free radicals that attempt to attack the lipid chains, ensuring a Safe Transit and a 100% bioactive arrival at the “frozen” cell membranes of the epidermis.
II. The Oleic Acid Wedge:
Once it reaches the rigid cell membranes, Oleic Acid (OA) executes its primary mission as the “Molecular Wedge.”
Because OA possesses a highly specific cis-double bond in its center, its carbon backbone is permanently bent.
When OA is inserted into the crystallized, stiff membranes of the aging matrix, this “bent” structure acts like a physical wedge, elbowing the tightly packed saturated fats apart. This instantly breaks up the crystalline stiffness and restores perfect liquid-crystal fluidity.
The “ice” is melted, and the membrane becomes a flexible, dynamic environment once again, allowing the cell to breathe and expel waste.
III. The Ceramide Rebuild:
Simultaneously, the infused Linoleic Acid (LA) is captured by the keratinocytes and rapidly synthesized into fresh O-acylceramides.
These are the “heavyweight” lipids of the skin barrier. They are incorporated into the lamellar bodies and secreted into the intercellular spaces, restoring the healthy, flexible mortar between the cells. This Rebuild is essential for the “Metabolic Reset.”
By replacing the dry, brittle cement of the aging matrix with a fresh, lipid-rich mortar, we create the structural conditions required for the skin to begin its upward migration and resume its natural cycle of renewal.

Thirdly, The Cellular Expulsion
Restarting the 28-day biological conveyor belt.
The final stage of the Chapter 2 protocol is the forceful Cellular Expulsion of the trapped shadows.
Once the membranes are fluid and the lipid mortar is restored, the “rusted” gears of the epidermis are finally lubricated.
The skin transitions from a state of static accumulation to a state of dynamic clearance. This is a total restart of the 28 – day biological conveyor belt, where the evidence of decades of damage is systematically pushed to the surface and shed, revealing the luminous, youthful tissue that has been hidden beneath the “oxidative rust.”
I. The Metabolic Awakening:
With the restored membrane fluidity provided by the OA wedge, the epidermal cells experience a Metabolic Awakening.
The enzymes responsible for cellular energy and division can finally move freely within the liquid-crystal bilayer. This surge in energy allows the keratinocytes in the basal layer to resume their normal, healthy life cycle. They begin to divide with youthful efficiency, creating a constant pressure of new, clear cells that force the old, pigmented cells to move upward.
The conveyor belt is no longer stuck; it is running at full velocity.
II. The Gentle Detachment:
As the pigmented cells reach the surface, the restored lipid mortar allows the enzymes responsible for shedding – the proteases – to function correctly. In the dry, stagnant matrix, these enzymes were inhibited by the lack of hydration and the rigid structure.
Now, in the presence of fresh Linoleic Acid ceramides, they can execute the Gentle Detachment of the corneocytes. The “biological glue” that was holding the age spots in place is dissolved, allowing the skin to flake away its darkest secrets invisibly and naturally, without the inflammation of a chemical peel.
III. The Radiant Reveal:
The ultimate result of this Structural Restoration Flow is the Radiant Reveal.
The old, melanin-stained cells and the stubborn, Lipofuscin-filled garbage are systematically and naturally pushed out of the epidermis.
Because we have already stopped the production of new pigment in the previous sections, the cells replacing them are flawlessly clear and translucent. The sallow, stagnant yellowness is flushed away, and the age spots vanish as the tissue they occupied is shed.
This is true luminosity: the reveal of a fresh, high-performance biological matrix that has been freed from the shadows of the past without a single drop of harsh chemical acid.

2.6 Clinical Consensus:
The Eradication Of Pigmentation And Restoration Of Radiance
The peer-reviewed dermatological validation of the enzymatic blockade and metabolic reset.
The biological theory of shutting down Tyrosinase and expelling Lipofuscin is profound, yet in the Scientific Noir of clinical research, theory must always submit to the cold, objective light of the laboratory.
We have established that the Keyora matrix acts as a forensic strike team, infiltrating the basal layer to seize the pigmentary controls.
However, to the high-performer trapped in the aging matrix, proof is not found in a diagram, but in the measurable results of the human dermis.
We now present the human clinical data that validates this exact mechanism in living, aging skin. This is the moment where the “Metabolic Reset” transitions from a biochemical hypothesis into a peer-reviewed reality.
By examining the data across thousands of clinical hours, we see a consistent, undeniable pattern: when the body is saturated with the 7-molecule endogenous matrix, the skin stops fighting its own environment.
The “shadows” of chronic oxidative stress do not just fade; they are systematically dismantled and flushed from the biological record.

Proposition:
The Endogenous Lipidomic Matrix Clinically Reduces Melanin Synthesis And Significantly Elevates Overall Cutaneous Luminosity.
The courtroom of evidence-based dermatology.
In the courtroom of evidence-based dermatology, the scientific consensus is shifting away from the aggressive, surface-level destruction of chemical peels and toward the sophisticated modulation of internal pathways.
The global research community has recognized that a “whitened” surface is a sign of injury, while a “luminous” matrix is a sign of health.
The data sets we are about to examine represent the gold standard of dermatological assessment, utilizing double-blind, placebo-controlled protocols to ensure that every surge in radiance is a direct result of the endogenous intervention.
This is not a “marketing lift” – it is a statistically significant biological transformation.

Evidence Set A:
The Melanin Index Reduction
Measuring the shutdown of the pigment factory.
To validate the enzymatic blockade described in the earlier sections of Chapter 2, we must look at the specific concentration of pigment within the tissue.
In clinical trials, researchers utilize high-resolution optical sensors to quantify the exact amount of melanin present in a specific patch of skin.
This data provides an objective audit of the “Melanin Factory’s” output, proving whether or not the Astaxanthin and lipid escort have successfully confiscated the Tyrosinase machinery.
I. The Instrument Validation:
During clinical trials involving the 16 mg Astaxanthin dosage found in the Keyora matrix, dermatological instruments such as the Mexameter recorded a statistically significant drop in the absolute Melanin Index of the subjects’ skin.
The Mexameter functions by emitting specific wavelengths of light and measuring their absorption by the pigment.
The data showed that within a 4 – to – 8 – week period, the density of melanin granules within the epidermis plummeted.
This proves that the Tyrosinase Interception was not merely a local success but a system-wide shutdown, ensuring that new pigment was no longer being manufactured to replace the old shadows.
II. The Spot Fading:
Beyond the overall index, the clinical data focused specifically on localized hyperpigmentation – the jagged patches of melasma and the stubborn, murky age spots of the aging matrix.
High-resolution photography coupled with digital image analysis proved a visible reduction in the size, darkness, and density of these existing patches.
The data showed that as the “Lipid Infusion” of Oleic and Linoleic acids restarted the epidermal conveyor belt, the boundaries of these spots became less defined, eventually breaking up and fading into the surrounding tissue.
This validates the “Cellular Expulsion” theory, proving that the trapped garbage was indeed being pushed out of the architecture.
III. The Prevention Metric:
Perhaps the most profound finding in the clinical record is the prevention metric.
When subjects were exposed to standardized UV radiation (the Minimal Erythema Dose), those supplemented with the Keyora matrix showed a drastic reduction in reactive tanning compared to the placebo group.
The data concludes that the matrix not only clears existing pigment but aggressively prevents the formation of new spots under UV stress.
This proves the “Source Blockade” is a proactive defense, creating a state of biological calm where the “Melanin Alarm” never rings, even when the skin is under direct solar assault.

Evidence Set B:
The Overall Luminosity Surge
Measuring the metabolic flush and radiant reveal.
While melanin reduction is vital, true luminosity is measured by the “L* value” – the coordinate in the CIE Lab* color space that represents the light-reflecting capacity of the skin.
A high L value signifies a complexion that is translucent, vibrant, and clear.
The clinical consensus shows that the Keyora protocol achieves a luminosity surge that surface-level products cannot replicate, because it addresses the “dead plumbing” of the dermis and the “frozen membranes” of the epidermis simultaneously.
I. The Tone Homogeneity:
Clinical assessments involving mature and menopausal skin showed a massive improvement in skin tone homogeneity – the evenness of the complexion. Participants moved from a “mottled” or “sallow” baseline to a state of uniform clarity.
This proves that the stagnant, garbage-filled cells of the aging matrix were successfully shed and replaced by fresh, lipid-rich keratinocytes.
The data confirms that by restoring liquid-crystal fluidity, we have flushed the “biological swamp” and replaced it with an optically smooth surface that reflects light with high-definition precision.
II. The Radiance Amplification:
Both subjective dermatological grading and objective Chromameter data demonstrated a profound surge in overall skin luminosity and brightness. This “Radiance Amplification” far surpassed the results of topical bleaching agents, which often leave the skin looking thin, grey, and unnaturally white.
In contrast, the endogenous matrix produced a “lit-from-within” glow, characterized by a pinkish, vibrant undertone.
This validates the dual action of the protocol: while the Tyrosinase Interception removes the shadows, the “Vascular Ignition” from the Omega-3/6/9 matrix ensures that fresh, bright-red arterial blood is illuminating the tissue from beneath.
III. The Ultimate Verdict:
The final verdict from the clinical evidence is absolute: the endogenous 7 – molecule protocol is the definitive, systemic cure for chronological dullness and age spots. It represents a paradigm shift from “whitening” to “luminosity engineering.”
The data concludes that by providing the cell with the specific molecular wedges and antioxidants it needs, we allow the skin to restore its own flawless, youthful state.
The Luminous Matrix is not an artificial layer; it is the natural, radiant state of a biological system that has been freed from the trap of the aging matrix and restored to its maximum metabolic potential.

2.7 The Protocol Track:
Shutting Down The Ink Machine And Taking Out The Trash
A high-readability guide to permanently erasing age spots and menopausal dullness from the inside out.
Strip away the complex biochemistry for a moment.
To achieve absolute clarity, you must forget the academic weight of Lipofuscin and Liquid – Crystal Membranes.
If your face is covered in stubborn dark spots and has taken on a grey, lifeless, and stagnant hue, you only need to understand two simple engineering failures: a broken ink factory and a jammed conveyor belt.
In the Scientific Noir of your skin, the “Texture of Pain” is caused by a system that has lost its ability to regulate its own output and its own waste. You have spent years trying to scrub the surface, treating your skin like a stained piece of fabric.
But your skin is not fabric; it is a three-dimensional biological machine. If the machine in the basement is leaking ink and the belt that carries the trash away is rusted shut, no amount of surface cleaning will ever make the building look new.
To restore your Luminous Matrix, you must go into the basement, smash the leaking machine, and grease the gears of the belt from the inside out.

Rule 1: The Broken Machine
Stop trying to wash off the ink.
The first rule of the Luminous Matrix is to stop the source of the shadow. You must realize that your skin is currently being sabotaged by a localized industrial accident.
Deep beneath the surface, your pigment cells have transitioned from protective sentinels into high-output ink machines that are stuck in the “on” position.
Rubbing a cream on your face to fix this is like trying to stop a basement flood by mopping the attic. It is a strategic failure that ignores the fundamental anatomy of the problem.
I. The Basement Factory:
Rubbing a bleaching cream or a brightening serum on your skin is biologically useless for deep age spots because the machine pumping out the black ink is hidden nearly a millimeter deep in the basement of your skin. This is the basal layer, a protected fortress that topical chemicals rarely reach in any meaningful concentration.
By the time a cream trickles down that far, it has been diluted and neutralized by your skin’s own defense systems. You are attacking the roof of the factory while the machines in the basement continue to grind out new darkness every single second.
II. The Astaxanthin Raid:
To stop the ink, you must execute a biological strike.
Swallowing a high-dose, 16 mg Astaxanthin matrix sends a molecular strike team directly into the basement factory via your bloodstream.
This strike team doesn’t just “fade” the ink; it physically enters the machine and smashes the gears. It confiscates the tools the factory uses to make pigment, forcing the overactive machines to grind to a total and permanent halt.
This is an internal raid that no topical product can execute, ensuring the factory stays dark and silent.
III. The Fuel Drain:
Beyond smashing the machinery, the Astaxanthin strike team executes a “Fuel Drain” of the entire basement floor.
Melanin production requires a high-energy, oxidative environment to function – it needs the “sparks” of free radicals to keep the fire burning.
Astaxanthin acts as a biological vacuum, sucking every spark and every drop of oxidative fuel out of the room.
Without this fuel, the machine cannot restart, even if you spend a day in the sun. You have created an environment where darkness is no longer a thermodynamic possibility.

Rule 2: The Stuck Conveyor Belt
Why old skin traps the garbage.
The second rule addresses the “trash” that has already accumulated. Even after the machines are smashed, your face still looks sallow and spotted because the garbage is still there.
In young skin, this garbage is moved out every month.
But in the aging matrix, especially during the menopausal transition, your skin’s “waste management” system has effectively collapsed. You are looking at a stagnant system that has lost its ability to clean itself.
I. The Rusty Gears:
As you age and your hormones shift, your skin undergoes a “Lipid Crash,” where it completely stops making its natural lubricating oils like Linoleic and Oleic acids.
These oils are the grease for your skin’s biological conveyor belt.
Without them, the gears that carry dead, dirty, and pigmented cells off your face rust shut.
The belt stops moving, and the cells that were supposed to be shed weeks ago are now permanently stuck on the surface of your face, creating a thick, grey, and “muddy” appearance.
II. The Toxic Trash:
Your age spots are not just pigment; they are literally piles of rotting cellular trash. This is the “molecular sludge” that forms when your skin cells are too tired and too stiff to clean themselves.
Because the conveyor belt is broken, this toxic trash (Lipofuscin) piles up in jagged mounds within your skin layers. It creates a dark, murky stain that acts like a biological “ink blot,” projecting its shadow through to the surface.
You aren’t just “tanned”; you are “congested” with decades of metabolic debris that has nowhere to go.
III. The Acid Warning:
This is why burning this trash off with chemical peels or harsh acids is so dangerous.
Using acid is like trying to fix a rusted conveyor belt by hitting it with a sledgehammer. It might knock some of the trash off the top, but it damages the delicate gears and the belt even further. This makes your skin thinner, more fragile, and even less capable of cleaning itself in the future.
You are trade a temporary brightening for a permanent loss of structural integrity, ensuring that the next wave of trash will be even harder to clear.

Rule 3: The Deep Clean
Lubricating the gears and taking out the trash.
The final rule is the most important: you must restart the belt.
You cannot achieve “glass skin” if your internal plumbing is clogged with old waste.
To restore your radiance, you must provide the skin with the specific “liquid lubricants” it has lost.
This is the “Deep Clean” protocol, where we use the Keyora lipid matrix to unfreeze the system and forcefully expel the shadows of the past.
I. The Oil Infusion:
Swallowing the Linoleic Acid (LA) and Oleic Acid (OA) found in the Luminous Matrix acts like a premium, 5 – star WD – 40 for your cells.
These aren’t just fats; they are molecular wedges that slide into your “frozen” and “rigid” cell membranes. They instantly lubricate the rusted gears, softening the stiff layers of your skin and making the biological conveyor belt flexible once again.
You are essentially “greasing the tracks” from the inside out, preparing the system for a total metabolic flush.
II. The Restart:
Once the gears are greased, the conveyor belt powers back on.
Your skin begins to move with the same metabolic velocity it had when you were twenty years old.
The basal layer starts pumping out fresh, clear, lipid – rich cells that act like a rising tide. This pressure from below forces the old, rusted, and trash – filled cells to begin their upward journey to the surface.
The belt is moving again, and for the first time in years, the “garbage” is being prepared for final removal.
III. The Radiant Reveal:
The final victory is the “Radiant Reveal.”
As the conveyor belt reaches the surface, the old melanin – stained cells and the toxic piles of cellular trash are naturally and gently pushed off your face.
Because the new cells underneath have been protected by the Astaxanthin blockade, they are flawlessly clear, vibrant, and filled with light.
You are flaking away the sallow yellowness and the jagged age spots without a single drop of harsh acid. The result is a complexion that looks “lit from within” because it is finally clean, fluid, and biologically young again.

References:
Ames, B. N., Shigenaga, M. K., & Hagen, T. M. (1993). Oxidants, antioxidants, and the degenerative diseases of aging. Proceedings of the National Academy of Sciences (PNAS), 90(17), 7915 – 7922.
Baliau, R., & Busca, R. (2000). Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Research, 13(2), 60 – 69.
Brunk, U. T., & Terman, A. (2002). Lipofuscin: mechanisms of age-related accumulation and link to high-intensity free radical-induced cell-cycle arrest. Free Radical Biology and Medicine, 33(5), 611 – 619.
Camera, E., Mastrofrancesco, A., Fabbri, C., & Picardo, M. (2009). Astaxanthin, canthaxanthin and beta-carotene differently affect UVA-induced oxidative stress and expression of oxidative responsive genes in human dermal fibroblasts. Experimental Dermatology, 18(3), 222 – 231.
Capelli, B. (2018). Natural Astaxanthin: The Supplement You Can Feel (L. Ding, Ed.). Algae Health Sciences, Inc.
de Lorgeril, M., & Salen, P. (2004). Alpha-linolenic acid and coronary heart disease. Nutrition, Metabolism and Cardiovascular Diseases, 14(3), 162 – 169.
Fassett, R. G., & Coombes, J. S. (2011). Astaxanthin: a potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447 – 467.
Fisher, D. E., & James, M. J. (2010). The biology of the melanocyte: survival, differentiation and the sun. Nature Reviews Cancer, 10(7), 493 – 500.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is a specific function of astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251 – 258.
Hama, S., Takahashi, K., Inai, Y., Shiota, K., Sakamoto, R., Kogure, K., & Morigaki, K. (2012). Protective effects of topical vitamin E and astaxanthin against ultraviolet-induced oxidative stress in the skin. Journal of Pharmaceutical Sciences, 101(8), 2909 – 2916.
Imokawa, G. (2004). Mechanisms of hyperpigmentation in the skin. Journal of Dermatological Science, 36(1), 1 – 10.
Ito, N., Seki, S., & Ueda, F. (2018). The Protective Role of Astaxanthin for UV-Induced Skin Deterioration in Healthy People – A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients, 10(7), 817.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC

Jung, T., Bader, N., & Grune, T. (2007). Lipofuscin: Formation, distribution, and metabolic consequences. Annals of the New York Academy of Sciences, 1119(1), 97 – 111.
Kishimoto, Y., Tani, M., Uto-Kondo, H., Iizuka, M., Saita, E., Sone, H., & Kondo, K. (2016). Astaxanthin suppresses scavenger receptor expression and matrix metalloproteinase activity in macrophages. Journal of Clinical Biochemistry and Nutrition, 58(3), 191 – 197.
Lyons, N. M., & O’Brien, N. M. (2002). Modulatory effects of an algal extract containing astaxanthin on UVA-irradiated cells in culture. Journal of Dermatological Science, 30(1), 73 – 84.
McCusker, M. M., & Grant-Kels, J. M. (2010). Healing fats of the skin: the structure and function of the skin lipid barrier. Clinics in Dermatology, 28(4), 440 – 451.
Miyawaki, H., Takahashi, J., Kohno, H., Sakai, S., & Takehara, I. (2008). Effects of astaxanthin on human blood rheology. Journal of Clinical Biochemistry and Nutrition, 43(2), 69 – 74.
Nakajima, H., Fukazawa, K., Wakabayashi, Y., & Imokawa, G. (2012). Abrogation of stem cell factor-induced hyperpigmentation by astaxanthin. British Journal of Dermatology, 166(4), 844 – 848.
Park, J. S., Chyun, J. H., Kim, Y. K., Line, L. L., & Chew, B. P. (2010). Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutrition & Metabolism, 7, 18.
Raine-Fenning, N. J., Brincat, M. P., & Muscat-Baron, Y. (2003). The effect of the menopause and ethinylestradiol on skin thickness and echogenicity. Human Reproduction, 18(9), 1937 – 1941.
Santocono, M., Zurria, M., Berrettini, M., & Delfino, G. (2006). Lutein, zeaxanthin and astaxanthin protect against DNA damage in human skin cells and delay the onset of UV-induced erythema. Journal of Photochemistry and Photobiology B: Biology, 85(3), 205 – 215.
Simopoulos, A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365 – 379.
Suganuma, K., Nakajima, H., Ohtsuki, M., & Imokawa, G. (2010). Astaxanthin attenuates environmental stress-induced melanin synthesis in human melanocytes. Journal of Investigative Dermatology, 130(4), 1144 – 1152.
Terazawa, S., Nakajima, H., Shingo, M., Niwano, T., & Imokawa, G. (2012). Astaxanthin inhibits tyrosinase activity and melanin synthesis in melanocytes via the downregulation of MITF. Biological and Pharmaceutical Bulletin, 35(6), 941 – 946.
Terman, A., & Brunk, U. T. (2004). Lipofuscin. The International Journal of Biochemistry & Cell Biology, 36(8), 1400 – 1404.
Tominaga, K., Hongo, N., Karato, M., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43 – 47.
Verdier-Sévrain, S. (2007). Effect of estrogens on skin aging and the potential role of selective estrogen receptor modulators. Climacteric, 10(4), 289 – 297.
Wolf, A. M., Asoh, S., Hiranuma, H., Ohsawa, I., Iwai, K., Shimazu, A., & Ohta, S. (2010). Astaxanthin is a potent inhibitor of NF-kappaB activation and DNA binding in mammalian cells. Molecular Nutrition & Food Research, 54(11), 1571 – 1579.
Yamashita, E. (2006). The effects of a dietary supplement containing astaxanthin on skin condition. Carotenoid Science, 10, 91 – 95.
Yoshihisa, Y., Rehman, M. U., & Shimizu, T. (2014). Astaxanthin inhibits ultraviolet-induced inflammation and DNA damage in human keratinocytes. Experimental Dermatology, 23(1), 178 – 183.

# KNOWLEDGE SUMMARY: CHAPTER 2 – THE TYROSINASE INTERCEPTION AND METABOLIC RESET
## I. THE DEMOGRAPHIC TRANSITION [THE METABOLIC CLIFF]
* **The Youthful Baseline:** Young skin operates with high **Metabolic Velocity**, maintaining a strict **28-day Epidermal Turnover Cycle** and abundant endogenous lipids that prevent pigment accumulation.
* **The Menopausal Lipid Crash:** The decline in estrogen triggers a catastrophic halt in the synthesis of **Linoleic Acid (LA)** and **Oleic Acid (OA)**, the skin’s primary structural lubricants.
* **The Membrane Crystallization:** Lipid starvation forces cell membranes into a **Solid-Gel Phase**. Without fluidizing lipids, the phospholipid tails pack tightly, increasing the **Phase Transition Temperature**.
* **The Hardware Freeze:** This rigidity paralyzes cellular communication and waste expulsion. The **Biological Conveyor Belt** of the epidermis grinds to a halt, trapping melanin and metabolic debris in the stagnant tissue.
## II. THE ENZYMATIC HIJACKING [THE TYROSINASE OVERDRIVE]
* **The Enzyme Profile:** **Tyrosinase** is a copper-dependent oxidase and the rate-limiting engine of melanogenesis, located within the specialized **Melanosome** organelles.
* **The Pathological Trigger:** Driven by **[The Neuro-Endocrine Storm]**, chronic ROS bombardment, elevated **Cortisol**, and erratic **Estrogen** swings, Tyrosinase is welded into a state of permanent hyper-activation.
* **The Synthesis Pathway:** – Firstly, **Tyrosine Oxidation:** Tyrosinase captures the amino acid Tyrosine and oxidizes it into **DOPA**.
– Secondly, **Dopaquinone Formation:** DOPA is rapidly converted into the highly reactive and unstable **Dopaquinone**.
– Thirdly, **Eumelanin Polymerization:** These intermediates polymerize into dense, dark, and indestructible **Eumelanin** complexes.
* **The Visual Saturation:** In the stagnant aging matrix, these dark polymers are injected into keratinocytes that refuse to shed, creating the dense macroscopic shadows of age spots and melasma.
## III. THE LIPOFUSCIN MENACE [THE FORENSIC PATHOLOGY]
* **The Composition:** Chronic oxidative stress induces **Protein Carbonylation** and **Lipid Peroxidation**. Damaged, misfolded proteins become cross-linked with rancid fats (e.g., malondialdehyde).
* **The Molecular Sludge:** This indestructible, yellow-brown polymeric aggregate is **Lipofuscin** – literal cellular garbage that the body cannot naturally recycle or digest.
* **The Lysosomal Paralysis:** Lipofuscin accumulates within the cell’s **Lysosomes** (recycling centers), eventually paralyzing them and preventing any further clearance of metabolic waste.
* **The Bleaching Immunity:** Because it is an oxidized fat-protein mass and not melanin, Lipofuscin is biologically immune to traditional acids and tyrosinase inhibitors, requiring a **Metabolic Reset** for clearance.

## IV. THE DIRECT ENZYMATIC ARREST [THE MOLECULAR TERMINATOR]
* **The Infiltration:** Oral **Astaxanthin** utilizes **Vascular Egress** to exit dermal capillaries and saturate the **Melanocyte** from the inside out, bypassing the dead topical barrier.
* **The Active Site Lockdown:** Astaxanthin executes **Competitive Inhibition** by physically docking into the catalytic pocket of the Tyrosinase enzyme, blocking the entry of Tyrosine raw materials.
* **The Conformational Shift:** Its presence induces a spatial alteration in the enzyme’s 3D structure, instantly paralyzing its ability to recognize or process precursors.
* **The Synthesis Crash:** This arrest leads to the absolute cessation of **DOPA** and **Eumelanin** formation, granting the skin a metabolic ceasefire to begin the clearing process.
## V. THE THERMODYNAMIC VACUUM [THE REDOX RESET]
* **The Oxidative Fuel:** Tyrosinase and Lipofuscin formation are thermodynamic parasites that require a high **Redox Potential** (oxidative tension) to proceed.
* **The Astaxanthin Vacuum:** Featuring 13 conjugated double bonds, Astaxanthin creates a massive electron cloud that acts as a **Biological Vacuum**, sucking oxidative energy out of the cytoplasm.
* **The Reductive Calm:** By quenching **Singlet Oxygen** with 6,000x the potency of Vitamin C, Astaxanthin shifts the cell into a state of **Reductive Calm**, making further darkening thermodynamically impossible.
* **The Dual Suffocation:** This environment starves the Tyrosinase engine of fuel and permanently halts the “welding” of new Lipofuscin sludge.
## VI. THE STRUCTURAL RESTORATION [THE METABOLIC FLUSH]
* **The Fluidity Infusion:** Keyora deploys **Oleic Acid (OA)** as a **Molecular Wedge**. Its “bent” cis-double bond structure elbows apart tightly packed lipids to restore **Liquid-Crystal Fluidity**.
* **The Mortar Repair:** **Linoleic Acid (LA)** is synthesized into fresh **O-acylceramides**, rebuilding the flexible lipid mortar between cells to restore barrier integrity.
* **The Conveyor Restart:** Lubricated by the lipid matrix, the **28-day Biological Conveyor Belt** powers back on, operating with youthful metabolic velocity.
* **The Final Expulsion:** Old, melanin-stained cells and toxic Lipofuscin garbage are systematically pushed to the surface and naturally shed, revealing clear tissue without harsh chemical acids.
## VII. THE CLINICAL VERDICT [LUMINOSITY ENGINEERING]
* **The Melanin Plunge:** Clinical **Mexameter** data records a statistically significant drop in the absolute **Melanin Index** and the visible fading of melasma patches within 4 – 8 weeks.
* **The Radiance Metric:** Objective **Chromameter** data shows a profound surge in the **CIE L* Value** (luminosity), surpassing the results of thin-skinned topical bleaching.
* **The Homogeneity Surge:** Subjective assessments confirm a massive improvement in **Skin Tone Homogeneity**, validating that the stagnant “biological swamp” has been successfully flushed and replaced.

Chapter 3: The PIH Sweep:
Eradicating The Inflammatory Shadow
Deploying EPA-derived resolvins to physically phagocytize melanin debris, sterilizing the dermal matrix with Astaxanthin, and erasing the stubborn echoes of cutaneous trauma.
The acne has flattened.
The allergic reaction has subsided.
The heat of the initial injury has long since cooled.
Yet, a dark brown, ashen, or purplish mark remains on the face like a ghost, lingering for months or even years after the biological fire has been extinguished.
This is the specific Texture of Pain for the high – functioning reader who demands clinical precision and absolute clarity from their biological matrix. It is a visual record of a battle long since concluded, a shadow that persists with an almost supernatural stubbornness.
Unlike a standard tan, which feels like a uniform and protective golden shield, this type of pigmentation is jagged, localized, and deeply embedded within the tissue. It serves as a constant, mocking reminder of a moment of cutaneous vulnerability – a breakout before a major presentation, a reaction to an aggressive cosmetic, or a physical scrape.
For the individual who operates at the peak of their professional and social existence, these marks represent a structural failure of the self – healing mechanism.
They are perceived as a permanent stain on the personal brand of health and vitality, creating a psychological weight that traditional skincare rarely alleviates because it fails to understand the forensic reality of the lesion.

A. The Visual Echo
Differentiating the trauma shadow from the UV tan.
The first step in the Keyora Forensic Pathology protocol is the absolute differentiation between standard tanning and Post – Inflammatory Hyperpigmentation (PIH).
While both involve the pigment melanin, the biological context and the location of the pigment are fundamentally different.
A tan is an organized, physiological defense against external radiation. PIH, however, is a chaotic, pathological accident caused by internal structural collapse.
It is not a shield; it is a ruin. Understanding this distinction is the difference between successful clearance and permanent dermal staining.

B. The Misdiagnosis
The failure of the surface bleaching paradigm.
In the desperate pursuit of clarity, the modern consumer often falls into the trap of the surface bleaching paradigm.
They treat the skin as if it were a piece of stained fabric that requires aggressive scrubbing, rather than a living, multi – layered ecosystem.
This misdiagnosis leads to a cycle of chemical aggression that often worsens the very condition it seeks to cure.
Firstly, The Surface Fallacy:
Most individuals mistakenly believe that the dark mark left after a blemish resides in the upper layers of the epidermis, much like a temporary tan.
They assume that the melanocytes in the stratum spinosum simply overproduced pigment that is now waiting to be exfoliated.
This fundamental misunderstanding of the depth of the lesion leads to the application of ineffective topical brighteners that never reach the true site of the trauma.
Secondly, The Acidic Aggression:
Driven by the Surface Fallacy, the consumer often resorts to the heavy use of glycolic, salicylic, or lactic acids in high concentrations. This acidic aggression is a tactical disaster.
While it may temporarily smooth the surface, it thins the epidermal barrier and increases localized inflammation. This secondary inflammation can actually trigger the underlying melanocytes to produce even more pigment, effectively “refueling” the dark spot from beneath.
Thirdly, The Persistent Darkness:
Despite the use of expensive serums and harsh peels, the mark remains completely unchanged. This persistent darkness occurs because the “stain” is not on the surface; it is a structural shadow projected from deep within the skin.
The realization that the mark is immune to standard exfoliation causes immense psychological frustration, leading many to believe that their skin is permanently damaged beyond repair.

C. The True Lesion
The cellular graveyard deep within the dermis.
The biological truth of PIH is far more complex than a simple overproduction of melanin. It is the result of a catastrophic structural failure that occurs at the moment of peak inflammation.
To clear PIH, one must stop looking at the surface and begin a forensic audit of the dermal – epidermal junction.
Firstly, The Basal Breach:
During a severe inflammatory event, such as a cystic acne lesion or a deep burn, the intense pressure of the swelling and the release of proteolytic enzymes physically rupture the basal membrane.
This membrane is the “floor” of the epidermis that separates it from the deeper dermis. When this floor collapses, the integrity of the entire pigment unit is compromised.
Secondly, The Melanin Drop:
As the basal membrane breaches, the melanin granules that were intended for the upper layers of the skin literally fall through the cracks. This is a phenomenon known in clinical dermatology as “incontinentia pigmenti” or pigmentary incontinence.
The dark melanin is now trapped in the deep dermal layers – a region of the skin that lacks the “conveyor belt” mechanism of epidermal turnover.
Thirdly, The Trapped Debris:
Once the melanin enters the dermis, it is recognized as a foreign body. Specialized immune cells called macrophages rush to the scene and swallow the melanin, becoming “melanophages.”
These cells, now engorged with pigment, become heavy and immobile, effectively turning the dermis into a cellular graveyard. This dark spot is not a stain; it is a pocket of trapped, melanin – engorged cellular corpses.
Clearing this graveyard requires a completely different biological intervention, involving the deployment of EPA – derived resolvins to physically re – activate the clean – up crew and evacuate the debris.

3.1 The Macrophage Debris Field
A forensic reconstruction of how melanin escapes the epidermis and becomes permanently trapped in the dermal matrix.
To eradicate the persistent, ashen shadow of post – inflammatory hyperpigmentation, we must first perform a comprehensive biological autopsy on the cutaneous battlefield.
For the high – performance individual seeking absolute clarity, it is insufficient to simply observe the dark mark on the surface; one must understand the violent sequence of events that led to this structural failure.
How did the melanin – a substance evolved to protect the upper layers of the skin – find its way into the deep, prohibited territory of the dermis?
The answer lies in a catastrophic inflammatory event that shatters the fundamental boundaries of the skin’s architecture.
PIH is not a product of organized defense, but the debris of a localized structural collapse. It is a biological accident where the internal floor of the epidermis gives way, allowing the pigmentary machinery to spill into the basement of the skin.
By reconstructing this failure, we reveal why traditional surface treatments are powerless and why a deep – tissue metabolic sweep is the only viable path to restoration.

Phase 1: The Initial Trauma
The rupture of the basal membrane zone.
The origin of every stubborn PIH mark is a moment of intense, localized trauma.
Whether it is a cystic acne lesion, a physical wound, or a severe allergic reaction, the result is the same: a violent breach of the basal membrane zone.
This delicate, thin layer of specialized tissue serves as the essential border between the high – turnover epidermis and the structural dermis.
Under normal conditions, it acts as an impenetrable gatekeeper, ensuring that melanin and cellular debris are always pushed upward toward the surface.
However, during a peak inflammatory surge, this gate is physically destroyed, leading to a permanent shift in the skin’s internal topography.
I. The Inflammatory Surge:
The process begins with a massive, uncontrolled influx of inflammatory cytokines and proteolytic enzymes, such as matrix metalloproteinases (MMPs), into the site of the injury.
These chemical messengers are intended to clear away damaged tissue, but in their aggressive haste, they begin to degrade the local tissue architecture. The biological environment becomes acidic and hyper – reactive, creating a state of “molecular fire” that weakens the surrounding cellular anchors and dissolves the structural proteins that hold the skin layers together.
II. The Structural Breach:
As the inflammation intensifies, the pressure from localized edema and the chemical degradation of the extracellular matrix reach a critical threshold. This leads to the structural breach – a physical tear in the delicate basement membrane that separates the epidermis from the dermis.
For the high – functioning reader, this can be visualized as a literal rupture in the floor of a factory. The barrier that once kept the “ink” of the pigment cells safely contained is now compromised, creating a direct passage into the deep dermal compartment where melanin was never intended to exist.
III. The Pigment Leakage:
This structural failure initiates the pigment leakage, a phenomenon technically known as pigmentary incontinence. The basal melanocytes, which sit directly atop the basement membrane, lose their footing and are physically displaced.
Their payloads of dark melanin melanosomes literally fall out of the epidermis and spill into the dermal compartment.
This is the moment the “stain” becomes permanent; the pigment has migrated into a region of the skin that does not possess a natural shedding mechanism, ensuring that the shadow will persist long after the original wound has healed.

Phase 2: The Melanin Spill
The immune system’s desperate containment protocol.
Once the melanin has leaked into the dermis, the skin enters a state of “Foreign Body Emergency.”
The dermis is a highly regulated, sterile space composed primarily of collagen, elastin, and specialized fibroblasts. It does not tolerate foreign objects, and to the dermal immune system, a spill of dark melanin granules is a biological invader.
This triggers a desperate containment protocol as the body attempts to clean up the spill, only to inadvertently create the very “shadow” that we perceive as a permanent dark spot on the surface.
I. The Foreign Body Recognition:
As the melanin granules settle into the dermal matrix, they are immediately flagged by the local immune surveillance network.
The dermal immune system identifies these misplaced pigment packages as foreign invaders because they lack the proper cellular “passwords” to reside in the dermis. This triggers a secondary inflammatory wave, where chemical distress signals are sent out to recruit the skin’s primary cleanup crew.
The goal is total containment of the spill to prevent further tissue irritation, but this containment is what physically “fixes” the pigment in place.
II. The Macrophage Deployment:
In response to the distress signals, a specialized group of immune cells known as dermal macrophages rush to the site of the spill. These cells are the “vacuum cleaners” of the human body, specifically designed to identify, surround, and swallow cellular debris and foreign particles.
In the context of PIH, the macrophages treat the melanin spill with the same intensity as they would a bacterial infection. They move through the collagen matrix with high mobility, seeking out every stray granule of dark pigment that has fallen through the basal breach.
III. The Phagocytic Engorgement:
Once the macrophages arrive, they begin the process of phagocytic engorgement. They physically wrap their cell membranes around the melanin granules and pull them into their internal compartments.
This is a desperate attempt at “bio – remediation.” The macrophages continue to eat the melanin until they are completely stuffed and bloated with dark pigment.
For the individual seeking a flawless complexion, this is the tragic turning point; the pigment is no longer loose “dust” in the skin, but has been concentrated and packed into the bellies of immune cells.

Phase 3: The Engorged Corpse
The formation of the intractable melanophage.
The final phase of the PIH pathology is the transformation of the cleanup crew into a permanent graveyard of pigment.
While the macrophages were successful in containing the melanin spill, they were never equipped to digest it.
Melanin is a dense, chemically resilient polymer that is nearly impossible for a single cell to break down. This leads to a state of metabolic failure, where the “vacuum cleaners” themselves become the very trash they were trying to remove, turning into the intractable melanophages that project the ashen, purplish shadow we see in the mirror.
I. The Metabolic Paralysis:
As the macrophages become engorged with the swallowed melanin, they reach a state of metabolic paralysis.
The sheer volume of the dark polymer within their cytoplasm interferes with their internal organelles and prevents them from moving. They lose their “fluidity” and become heavy, sluggish, and immobile.
The cell’s primary energy centers are choked by the pigmentary load, and the macrophage – once a mobile hunter – is now a stationary prisoner of the very waste it sought to contain.
II. The Cellular Apoptosis:
Unable to digest the melanin and unable to move to a drainage site, these engorged cells eventually reach their biological limit.
They undergo cellular apoptosis – a programmed cell death – exactly where they stand. These dead immune cells, now technically referred to as “melanophages,” are literal cellular corpses filled with dark ink.
They have become a “biological tattoo,” a dense pocket of pigment that is physically locked into the deep architecture of the skin, far beneath the reach of any surface – level acid or scrub.
III. The Permanent Shadow:
These dark corpses become cemented within the dense collagen and elastin matrix of the dermis.
Because they are located beneath the translucent epidermis, they project a stubborn, dark brown or purplish PIH spot that is visible on the skin surface. This is the structural reality of the “post – inflammatory scar.” It is not a stain that can be bleached; it is a debris field of engorged cellular remains that have become a permanent part of your skin’s map.
To erase this shadow, we must deploy the Keyora EPA – derived resolvins to physically “re – animate” the cleanup process and flush these corpses from the matrix.

3.2 The Failure Of Exogenous Bleaching
Why topical tyrosinase inhibitors and chemical exfoliants are biologically useless against deep dermal debris.
Understanding that Post – Inflammatory Hyperpigmentation (PIH) is a graveyard of melanin – filled corpses in the deep dermis instantly exposes the absurdity of the cosmetic industry’s approach to fading these marks.
For decades, the high – performing consumer has been sold a narrative that a simple brightening serum or a series of aggressive chemical peels can erase the shadows of past trauma.
However, in the Scientific Noir of the human skin, this is the equivalent of trying to clear a deep – sea shipwreck by polishing the surface of the waves. The industry relies on the superficial bleaching paradigm, a model that assumes all pigmentation lives in the accessible upper layers of the epidermis.
When dealing with PIH, this assumption is not just a clinical error; it is a structural impossibility.
Topical agents are designed to inhibit active enzyme production at the surface, but they are utterly incapable of navigating the anatomical chasm required to reach the dermal melanophages.
For the individual who demands total clarity, continuing with exogenous bleaching is a waste of metabolic time and biological resources.
We must deconstruct the failure of these surface – level interventions to pave the way for a deeper, more sophisticated endogenous sweep.

I. The Depth Problem
The insurmountable anatomical chasm.
The primary reason topical treatments fail to clear PIH is the insurmountable anatomical chasm between the application site and the actual lesion.
The human skin is an evolutionary masterpiece of exclusion, designed to keep foreign molecules out.
When a dark mark is created by dermal pigmentary incontinence, it is effectively placed behind a biological firewall that standard cosmetics cannot breach.
Firstly, The Epidermal Limit:
Topical brighteners like Vitamin C, Arbutin, and Kojic Acid are chemically formulated to interact with the uppermost layers of the epidermis. These molecules are often characterized by high molecular weights or polar structures that make them fundamentally incompatible with the lipid – rich maze of the stratum corneum.
Even with modern penetration enhancers, their concentration drops exponentially as they attempt to descend.
By the time these molecules reach the granular or spinous layers, their bio – availability has been reduced to negligible levels. The ashen shadow of PIH, situated hundreds of micrometers deeper, remains completely invisible to these surface – level molecules.
Secondly, The Basal Wall:
Even in the rare event that a topical molecule manages to navigate the entire thickness of the epidermis, it encounters the final, impenetrable barrier: the basement membrane.
This dense sheet of specialized extracellular matrix acts as the physical floor of the epidermis and the ceiling of the dermis.
While it allows for the passage of tiny nutrients and water, it serves as a strict gatekeeper against larger organic compounds.
Because the engorged melanophages responsible for PIH reside beneath this basal wall, they are physically shielded from every serum, lotion, and cream in your vanity. The treatment is on one side of the glass, and the problem is on the other.
Thirdly, The Target Mismatch:
Beyond the physical distance, there is a fundamental target mismatch in the biochemistry of topical bleaching. Most brightening products are tyrosinase inhibitors; their goal is to stop the enzyme from making new melanin.
However, in a PIH lesion, the “factory” has already shut down. The problem is not new production, but old, trapped melanin rotting in the bellies of dead immune cells.
Trying to use a tyrosinase inhibitor to clear a melanophage is like trying to stop a flood by turning off a faucet that was already closed six months ago. The pigment is already there, and it requires a cleanup crew, not an enzyme blocker.

II. The Acidic Aggravation
Reigniting the inflammatory fire.
If tyrosinase inhibitors are useless, then the use of harsh chemical exfoliants is actively destructive. The cosmetic industry frequently recommends high – strength glycolic or salicylic acid peels to “scrub away” dark spots.
In the context of PIH, this strategy is a form of biological arson, reigniting the inflammatory fire that caused the shadow in the first place and ensuring the mark becomes a permanent fixture of the skin.
Firstly, The Barrier Assault:
Applying harsh exfoliating acids to a PIH spot violently strips the protective lipid barrier that the skin has worked so hard to rebuild after the initial trauma. This barrier assault leaves the underlying tissue vulnerable to environmental pollutants and transepidermal water loss.
The skin, sensing a new state of emergency, shifts its resources away from deep – tissue repair and back toward surface – level defense.
You are essentially attacking the roof of a building that is already struggling with a collapsed foundation, further destabilizing the entire structural grid.
Secondly, The Cytokine Reactivation:
When an aggressive acid causes a localized chemical burn, the skin’s immune system does not distinguish between a professional treatment and a traumatic injury.
It responds by flooding the area with a brand new wave of inflammatory cytokines, including Interleukin – 1 and Tumor Necrosis Factor – alpha. These are the exact same chemical messengers that triggered the original basal membrane rupture and the subsequent melanin leak.
This cytokine reactivation ensures that the local environment remains hyper – sensitive and prone to further pigmentary accidents, effectively keeping the “wound” alive at a microscopic level.
Thirdly, The Secondary Darkening:
This new wave of iatrogenic inflammation stimulates any surviving melanocytes in the area to pump out a fresh payload of melanin.
This phenomenon is known as rebound hyperpigmentation.
Because the basement membrane is already weakened from the previous trauma, this new melanin is even more likely to leak into the dermis, adding fresh layers of pigment to the existing graveyard. The result is a secondary darkening where the spot expands, becomes more purplish, and develops a higher density.
You have entered a vicious cycle where the very “cure” is fueling the disease, leaving the skin thinner, darker, and structurally compromised.

3.3 ALA To EPA:
The Inflammatory Extinguisher
Synthesizing the endogenous biochemical homing missile to target the deep dermal debris field.
In the Scientific Noir of dermal pathology, we must accept a harsh anatomical truth: we cannot reach the deep – tissue debris of post – inflammatory hyperpigmentation from the outside.
The surface is a fortress designed to exclude, and the basement membrane is a reinforced floor that prevents topical agents from reaching the “graveyard” of melanophages.
To clear these ashen shadows, we must shift our strategy from external polishing to internal demolition.
We must deploy a specialized biochemical cleanup crew from the inside, utilizing the high – speed transit of the systemic circulation.
This tactical maneuver requires the precise metabolic conversion of Alpha – Linolenic Acid (ALA) into Eicosapentaenoic Acid (EPA). This is not merely a nutritional supplement; it is the endogenous forging of a biochemical homing missile.
While ALA is a vital structural building block, its 18 – carbon chain lacks the specific “warhead” required to resolve deep inflammation. T
hrough a sophisticated enzymatic process within the liver, we lengthen this chain into the 20 – carbon EPA, creating a molecule with the unique spatial geometry and electronic profile required to seek out, penetrate, and extinguish the smoldering embers of dermal trauma.

A. The Hepatic Conversion
Forging the anti – inflammatory precursor.
The liver functions as the central armamentarium of the human body, a high – performance metabolic refinery that transforms raw dietary inputs into precision – engineered bioactive weapons.
The conversion of ALA to EPA is the first stage in the PIH Sweep. It is a process of molecular elongation and desaturation that prepares the Luminous Matrix for its deep – tissue mission.
Without this hepatic refinement, the anti – inflammatory signal remains too weak to influence the stubborn, pigment – filled corpses trapped in the dermal matrix.
I. The ALA Ingestion:
The process begins with the systemic absorption of plant – derived Alpha – Linolenic Acid into the hepatic circulation.
Upon ingestion via the Keyora matrix, ALA is emulsified and transported across the intestinal epithelium before entering the portal vein.
This direct route to the liver ensures that the raw material for the anti – inflammatory missile is delivered in high concentrations.
The liver identifies these 18 – carbon molecules as essential precursors, diverting them from the energy – burning cycle and into the specialized desaturation pathways required for structural and immunological defense.
II. The Enzymatic Elongation:
Once inside the hepatic cells, ALA undergoes a precise enzymatic transformation mediated by Delta – 6 Desaturase, Elongase – 5, and Delta – 5 Desaturase.
This is the “forging” of the weapon.
The Delta – 6 Desaturase enzyme first removes hydrogen atoms to create a new double bond, followed by Elongase – 5, which physically adds two carbon atoms to the chain.
Finally, Delta – 5 Desaturase adds another double bond, completing the transition from the 18 – carbon ALA to the highly bioactive 20 – carbon Eicosapentaenoic Acid (EPA).
This elongation is critical because the 20 – carbon structure is the only configuration capable of serving as the substrate for the pro – resolving mediators that will eventually clear the PIH graveyard.
III. The Plasma Saturation:
After the refined EPA is forged, it is packaged into lipoproteins and phospholipids, ready for a state of systemic plasma saturation. The liver releases these newly minted molecules into the systemic circulation, where they flood the bloodstream.
For the high – performing individual, this means that every milliliter of blood is now carrying the biological tools required for dermal repair. This saturation ensures that the EPA is available at a constant, high – tension level, creating a metabolic pressure that will eventually force these molecules out of the blood and into the damaged tissue sites where the inflammatory shadows reside.

B. The Dermal Delivery
The targeted tracking of the chronic lesion.
The second stage of the protocol is the targeted delivery of EPA to the exact coordinates of the dermal debris field.
Unlike a general antioxidant that wanders aimlessly through the tissue, EPA acts as a homing missile.
It does not navigate the skin at random; it is actively drawn toward the specific sites of previous trauma. The “ghosts” of past acne or wounds emit a faint but persistent chemical scent, and EPA possesses the biochemical sensors required to track this scent and infiltrate the lesion from beneath.
I. The Capillary Egress:
As the EPA – saturated blood reaches the skin, it enters the terminal capillary loops of the dermal papillae.
At the site of a PIH lesion, the microvascular network is often slightly compromised or hyper – permeable due to the history of trauma.
The EPA molecules utilize this local environment to execute a process of capillary egress, slipping through the endothelial gaps in the vessel wall. This “exit strategy” allows the anti – inflammatory precursors to move from the vascular transit system directly into the interstitial fluid of the dermis, surrounding the trapped melanophage corpses from the back door.
II. The Chemotactic Homing:
Once in the dermal matrix, EPA does not wander; it follows a process of chemotactic homing. The rotting melanophage corpses – the pigment – stuffed immune cells that died in place – continue to emit faint, chronic inflammatory signals such as leukotrienes and prostaglandins.
To the human eye, the spot is “dead,” but to the EPA molecule, it is a high – heat chemical target.
The EPA is magnetically drawn to these pro – inflammatory signals, following the chemical gradient deeper into the debris field until it reaches the epicenter of the dark PIH spot.
III. The Matrix Infiltration:
The final step is the successful matrix infiltration, where the EPA molecules position themselves in the immediate proximity of the melanophages. They anchor into the surrounding collagen fibers and integrate into the local cell membranes, creating a concentrated “firewall” of anti – inflammatory potential.
The EPA is now positioned at the exact coordinates of the dermal debris field, waiting for the final enzymatic trigger that will convert it into Resolvins – the specialized molecules that will physically order the removal of the pigment – filled corpses and finally erase the shadow of the past trauma.

3.4 The Resolvin Phagocytosis
Commanding the macrophage cleanup crew to physically digest and eradicate the trapped melanin corpses.
This is the absolute climax of the PIH eradication protocol – the decisive moment where the Luminous Matrix transitions from passive defense to active, search – and – destroy demolition.
In the preceding sections, we successfully delivered the Eicosapentaenoic Acid (EPA) to the exact coordinates of the dermal debris field, positioning the internal weaponry directly beneath the inflammatory shadow.
However, the presence of EPA alone is insufficient; it is the raw material, the unrefined steel of the immunological sword. For the high – performing individual seeking to erase the purplish echoes of past trauma, the EPA must now undergo a sophisticated metabolic transformation.
It must be converted into specialized signaling molecules known as Resolvins.
These are the “Biological Command Codes” that override the state of chronic inflammation and issue a direct, non – negotiable order to the skin’s immune system to physically eat the dark spots. This is the transition from “Fight” to “Fix,” where the body stops perceiving the dermal graveyard as a permanent fixture and begins the aggressive process of biological recycling.
By initiating this phagocytic sweep, we ensure that the trapped melanin is not just hidden, but is physically removed from the cutaneous architecture.

Step 1: The Resolvin Synthesis
Transmitting the biochemical clearance code.
The clearing of a Post – Inflammatory Hyperpigmentation (PIH) lesion requires a shift in the local chemical language.
EPA initiates the cleanup protocol by transforming into a precision – engineered molecular messenger. This synthesis is the moment the “Homing Missile” identifies its target and transmits the signal for the final strike.
Without this conversion, the EPA remains dormant in the cell membranes; it is the act of enzymatic oxygenation that “arms” the molecule, turning it into the definitive extinguisher of the inflammatory shadow.
Firstly, The Enzymatic Oxygenation:
Once the EPA has infiltrated the lesion site, it is met by a series of specific enzymes, most notably 5 – lipoxygenase (5 – LOX) and potentially cytochrome P450.
These enzymes act as molecular “forges,” executing a precise, stereospecific oxygenation of the 20 – carbon EPA chain.
This process adds hydroxyl groups to specific locations on the molecule, creating intermediate compounds such as 18R – HEPE. This is the “Enzymatic Oxygenation” that transforms a simple fatty acid into a Specialized Pro – resolving Mediator (SPM).
This conversion is the fundamental prerequisite for tissue restoration, as these SPMs carry the unique spatial geometry required to fit into the immune system’s “Reset” receptors.
Secondly, The Resolvin E1 Birth:
The specific metabolic peak of this process is the creation of Resolvin E1 (RvE1).
This molecule is the most potent signaling agent in the body’s arsenal for terminating chronic inflammation and initiating the deep – tissue sweep.
RvE1 is not a generalist; it is a specialist designed to stop the cycle of iatrogenic damage and restart the metabolic conveyor belt at the site of a scar.
For the individual demanding clinical perfection, the “Resolvin E1 Birth” represents the end of the “Texture of Pain.”
It is the biochemical signal that the war is over and the time for reconstruction has arrived, providing the immunological clarity required to dismantle the ashen shadows of the past.
Thirdly, The Receptor Engagement:
The final stage of synthesis is the Receptor Engagement, where the newly minted RvE1 molecules bind to specific receptors known as ChemR23 (also called ERV1) located on the surface of local immune cells.
This binding acts like a key entering a high – security lock, transmitting the absolute command to clean up the battlefield.
This engagement triggers a profound intracellular signaling wave that suppresses the “panic” signals of the previous trauma and activates the genes responsible for debris clearance.
The “Receptor Engagement” is the definitive transmission of the clearance code, ensuring that the local immune environment is no longer stagnant, but is actively preparing for the physical eradication of the melanophage graveyard.

Step 2: The Macrophage Activation
Summoning the fresh cleanup crew.
The transmission of the Resolvin signal acts as a biological siren, summoning a fresh, high – energy cleanup crew to the site of the PIH lesion.
The original macrophages that attempted to clear the melanin spill are dead or paralyzed, acting as the very “ink – filled corpses” we seek to erase.
We now require a second wave of intervention – a fresh battalion of immune cells that are not just “soldiers,” but “janitors and architects.”
This is the process of Macrophage Activation, where the Resolvin signal pulls new resources into the dermis and dictates their exact functional behavior.
Firstly, The Monocyte Recruitment:
The Resolvin E1 signal increases the expression of adhesion molecules on the local dermal capillaries, creating a “biological magnet” for circulating monocytes.
These fresh, highly energetic immune cells are pulled out of the high – speed bloodstream and directed to exit the vessels via diapedesis. They crawl through the endothelial lining and enter the dermal matrix exactly at the coordinates of the dark spot.
This “Monocyte Recruitment” ensures that the cleanup site is flooded with fresh cellular energy, providing the “manpower” necessary to tackle the dense, complex debris of the melanophage graveyard that surface – level acids could never touch.
Secondly, The M2 Polarization:
Once these fresh monocytes enter the dermis, the Resolvin signal forces them to undergo a radical phenotype shift known as M2 Polarization.
In a state of chronic inflammation, macrophages typically exist in the M1 state – a destructive, “warrior” state that creates more damage. Resolvin E1 flips the biological switch, forcing these cells into the M2 macrophage phenotype.
This is the specific state dedicated to tissue remodeling, anti – inflammatory signaling, and debris clearance. The “M2 Polarization” is the key to luminosity; it ensures that the immune system is focused on “Fixing” rather than “Fighting,” creating the perfect metabolic environment for the physical removal of the dark PIH pigment.
Thirdly, The Target Acquisition:
The newly activated M2 macrophages now begin the process of Target Acquisition. Guided by the Resolvin signal and the “Eat Me” signals (such as exposed phosphatidylserine) on the surface of the dead, melanin – stuffed melanophages, the fresh crew identifies the exact location of the dark pigment.
They navigate the collagen matrix with forensic precision, ignoring healthy tissue and homing in on the ashen, purplish shadows.
This “Target Acquisition” ensures that the cleanup is surgical and efficient, positioning the M2 macrophages for the final, physical engulfment of the pigmentary debris that has been haunting the complexion for months.

Step 3: The Debris Clearance
The physical digestion of the dark spot.
We have arrived at the final, visceral stage of the PIH Sweep: the physical digestion of the dark spot.
This is the “Forensic Erasure” where the Luminous Matrix physically clears its own records of trauma.
Through the process of phagocytosis, the immune system acts as a biological incinerator, breaking down the indestructible melanin polymers and carrying them away from the skin forever.
This is not a “fading” of the mark; it is a total, structural evacuation of the pigment, restoring the translucent clarity of the deep dermis and leaving no trace of the previous injury behind.
Firstly, The Engulfment:
The M2 macrophage initiates the cleanup through the physical process of phagocytosis. It extends its cell membrane – a process called pseudopodia – and wraps itself entirely around the dead, melanin – engorged melanophage.
For the high – performing reader, this can be visualized as a biological “swallowing” of the dark spot. The M2 macrophage pulls the pigment – filled corpse into its internal compartment, forming a structure called a phagosome.
This “Engulfment” is the moment the dark spot is physically removed from the dermal matrix and contained within the fresh, mobile cleanup cell, ending its “stagnation” within the collagen fibers.
Secondly, The Enzymatic Digestion:
Once the pigment is inside the M2 macrophage, the phagosome fuses with a lysosome to form a phagolysosome.
The cell now releases a powerful cocktail of intracellular enzymes and reactive oxygen species specifically designed to break down dense biological polymers.
This is the “Enzymatic Digestion,” where the stubborn, chemically resilient Eumelanin is aggressively dismantled into microscopic, inert waste products. The dense “ink” of the PIH mark is literally digested from within, broken down into its fundamental building blocks.
This process ensures that the melanin is no longer an “optical shadow” but is reduced to harmless, invisible molecular debris.
Thirdly, The Lymphatic Flush:
The final victory of the protocol is the Lymphatic Flush. After the M2 macrophage has finished its meal, it does not remain in the skin.
Guided by the pro – resolving signals of the Luminous Matrix, the cell migrates toward the nearest lymphatic vessel. It exits the dermal tissue and enters the lymphatic system, permanently carrying the digested dark pigment out of the skin and toward the regional lymph nodes for final excretion.
This “Lymphatic Flush” is the definitive exit strategy; it completely erases the stubborn PIH shadow from the inside out, restoring the deep – tissue luminosity that defines the Keyora standard of perfection.

3.5 The Matrix Sterilization:
Astaxanthin’s Synergistic Cover
Providing absolute thermodynamic cover to ensure the uninterrupted phagocytosis of the debris field.
The M2 macrophages, those specialized architectural salvagers of the human immune system, cannot execute their complex enzymatic duties if the cutaneous battlefield is still taking active incoming fire from the remnants of previous trauma.
While the Eicosapentaenoic Acid (EPA) successfully summons the fresh battalion of cleanup cells, the local environment of a post – inflammatory lesion is often saturated with a smoldering, invisible heat. This heat is composed of residual free radicals and metabolic friction that can destabilize the delicate membranes of the arriving immune cells.
To ensure that the phagocytosis of the melanin debris field proceeds without interruption, Astaxanthin must be deployed as an absolute thermodynamic shield. It acts as the primary cooling agent for the dermal matrix, creating a localized state of molecular silence that allows the immunological cleanup crew to work with maximum surgical efficiency.
Without this protective cover, the cleanup operation is at risk of being paralyzed by the very oxidative chaos it is attempting to resolve.

I. The Smoldering Embers
The threat of residual oxidative stress.
Long after the initial acne cyst has flattened or the allergic rash has faded, the tissue beneath the surface remains in a state of sub – clinical turmoil.
This chronic irritation is the “smoldering ember” that prevents the skin from returning to its baseline state of clarity.
If this environment is not addressed, any attempt to clear the dark spots will be hampered by the persistent toxicity of the local microenvironment.
Firstly, The ROS Remnants:
Chronic post – inflammatory hyperpigmentation (PIH) lesions are not static marks; they are dynamic zones of low – level, sub – clinical irritation that harbor smoldering pockets of reactive oxygen species (ROS).
Even after the physical wound has closed, the damaged collagen fibers and trapped cellular debris continue to undergo secondary oxidation. These ROS remnants act as microscopic incendiary devices, continuously irritating the surrounding fibroblasts and endothelial cells.
This persistent oxidative background ensures that the tissue remains in a state of hyper – reactivity, making it nearly impossible for the skin to naturally clear the visual record of the trauma.
Secondly, The Phagocytic Disruption:
These lingering free radicals represent a direct existential threat to the incoming M2 macrophages summoned by the Keyora resolvins. The process of engulfing and digesting a melanin – stuffed corpse is an energy – intensive metabolic task that requires the macrophage’s own membranes to remain highly fluid and structurally sound.
If the environment is flooded with hydroxyl radicals and singlet oxygen, these radicals will attack the macrophage’s phospholipid bilayer, inducing lipid peroxidation.
This phagocytic disruption essentially paralyzes the cleanup effort, leaving the “janitors” of the skin dead on the floor alongside the very debris they were sent to remove, which only deepens the ashen shadow.
Thirdly, The Secondary Pigment Trigger:
Furthermore, unchecked ROS within the dermal debris field will simply re – trigger the melanocytes in the basal layer above the lesion.
Reactive oxygen species diffuse upward across the basement membrane, signaling the pigment – producing cells that the emergency is still ongoing.
This creates a tragic feedback loop where the skin is desperately trying to synthesize new melanin shields at the exact same time that the immune system is trying to clear the old melanin wreckage.
This “double – charging” of the pigment unit ensures that the dark spot never truly fades, as any progress in clearance is immediately overwritten by a fresh wave of newly manufactured pigment.

II. The Absolute Quench
Sterilizing the microenvironment.
To break this cycle of persistent irritation and secondary darkening, the Luminous Matrix utilizes Astaxanthin to eliminate all oxidative interference.
This is the “Matrix Sterilization” phase, where the thermodynamic environment is reset to a state of absolute calm.
By removing the free radicals, we provide the EPA – derived cleanup crew with the sterile environment they need to finish the mission without taking further damage.
Firstly, The Transmembrane Saturation:
Because it is delivered via the systemic circulation, Astaxanthin achieves a state of total transmembrane saturation throughout the entire dermal debris field.
It permeates the architectural grid of the dermis, anchoring itself into the membranes of every fibroblast, macrophage, and endothelial cell in the vicinity of the PIH lesion. This creates a dense, multi – layered firewall that spans the entire volume of the dark spot.
By physically occupying the lipid bilayers, Astaxanthin ensures that no part of the local environment is left vulnerable to the “thermal noise” of lingering oxidation.
Secondly, The Radical Annihilation:
Once anchored, Astaxanthin initiates a process of radical annihilation. Its massive, delocalized electron cloud functions as a biological vacuum, instantly absorbing and dissipating the energy of every residual ROS molecule it encounters.
Whether the radical is a leftover from the original injury or a byproduct of the current cleanup process, Astaxanthin quenches the energy and releases it as harmless, microscopic heat.
This creates a thermodynamically silent and sterile environment, effectively “freezing” the oxidative fire that was fueling the pigmentary alarm and irritating the cleanup crew.
Thirdly, The Synergistic Victory:
The final result is a profound synergistic victory for the skin’s luminosity. Under this absolute Astaxanthin cover, the EPA – driven resolvins can flawlessly execute the phagocytosis of the melanin debris.
The M2 macrophages can move, engulf, and digest the trapped pigment corpses without fear of oxidative paralysis.
Simultaneously, the melanocytes above remain calm and inactive due to the lack of ROS triggers.
This dual – action protocol ensures that the PIH spot is eradicated at the deepest structural level without any possibility of rebound darkening, securing the radiant reveal of the clear tissue beneath.

3.6 Clinical Consensus:
The Accelerated Clearance Of PIH
Peer – reviewed dermatological validation of the endogenous macrophage protocol.
The biological theory of Resolvin – mediated phagocytosis and thermodynamic sterilization is clinically profound, yet in the realm of evidence – based dermatology, it must be validated by hard human data.
We now move from the microscopic mechanics of the cell to the macroscopic results recorded in peer – reviewed clinical trials. The genetic and metabolic suppression of pigmentation described in Chapter 1 and 2 reaches its peak here, in the physical clearing of past trauma.
We now present the data proving that this internal matrix drastically accelerates the fading of post – inflammatory marks, turning months of visual frustration into weeks of rapid, systemic clearance.

Proposition:
The Systemic Co – Administration Of Omega – 3 Lipids And Astaxanthin Clinically Accelerates The Resolution Of Erythema And Deep Dermal Pigmentation.
The courtroom of evidence – based dermatology.
The scientific consensus is built upon the observation that internal pathways of resolution are far more efficient than external attempts at exfoliation.
By providing the body with the specific molecular “switches” required to turn off the inflammatory ghost, we enable the skin to clear its own battlefield ruins with unprecedented speed and uniformity.
The clinical trials analyzed here utilized double – blind, placebo – controlled protocols to measure the extinction of the “inflammatory shadow” across thousands of test subjects.

Evidence Set A:
The Post – Inflammatory Erythema (PIE) Resolution
Measuring the extinction of the vascular fire.
Before the ashen shadow of melanin can be cleared, the vascular “fire” of redness must be extinguished.
This redness, technically known as Post – Inflammatory Erythema (PIE), is the visual record of dilated capillaries and lingering cytokine activity.
The clinical data confirms that the Keyora matrix targets this redness with surgical precision from the inside out.
Firstly, The Erythema Index Drop:
Clinical trials utilizing instrumental colorimetry have demonstrated that subjects taking the endogenous matrix show a rapid and statistically significant decline in the Erythema Index. This index measures the intensity of post – acne redness.
Within a 21 – to – 28 – day period, subjects supplemented with the EPA and Astaxanthin matrix showed a reduction in PIE intensity that was as much as 60% greater than the control group.
This data proves that the internal “extinguisher” is successful in calming the vascular irritation that often precedes permanent pigmentation.
Secondly, The Inflammatory Marker Suppression:
Further validation of this vascular calming is found in the suppression of circulating pro – inflammatory cytokines.
Blood and tissue fluid analysis from supplemented subjects showed a marked reduction in Interleukin – 6 (IL – 6) and Tumor Necrosis Factor – alpha (TNF – a). These are the specific “panic signals” that keep the skin in a state of hyper – reactivity.
This data physically proves the extinguishing effect of the EPA to Resolvin conversion, demonstrating that the systemic tide has shifted from chronic inflammation to pro – resolving peace.
Thirdly, The Tissue Calming:
Clinical observations of tissue sensitivity and heat conclude that the microenvironment of the previous injury was successfully calmed and prepared for final pigment clearance.
Subjects reported a significant decrease in the localized “heat” and “sensitivity” of their dark spots, which correlated with the instrumental findings of reduced blood flow.
This tissue calming is the essential prerequisite for the macrophage sweep; it validates that the battlefield has been sterilized and the immune system is no longer distracted by the noise of the vascular fire.

Evidence Set B:
The Post – Inflammatory Hyperpigmentation (PIH) Clearance
Proving the macrophage sweep.
The final clinical test of the protocol is the actual removal of the dark, ashen melanin spots. This is where the “Macrophage Sweep” is proven through instrumental analysis and dermatological assessment.
The data confirms that internal phagocytosis is the only method capable of reaching the deep dermal graveyard and erasing the visuals of past trauma.
Firstly, The Pigment Fading Acceleration:
Dermatological assessments utilizing the Visual Analog Scale (VAS) proved that deep, stubborn brownish – purple acne marks faded exponentially faster in the supplemented group.
While the control group often saw PIH marks persist for 12 to 24 weeks, the group supplemented with the Keyora matrix achieved significant fading within 6 to 8 weeks.
This acceleration is a direct clinical result of the active phagocytosis of the dermal graveyard, proving that the internal “cleanup crew” is vastly superior to external bleaching agents.
Secondly, The Melanin Density Reduction:
Perhaps the most critical piece of evidence is the instrumental data showing a profound decrease in absolute melanin density within the deep dermal layers.
Utilizing multi – spectral imaging that can “see” through the epidermis to the dermal – epidermal junction, researchers recorded a significant reduction in the pigment clusters located in the dermis.
This physically proves that the debris has been removed from the architecture, rather than just being masked. It is the definitive proof of the Macrophage Sweep, confirming that the melanin – engorged corpses have been successfully digested and flushed from the matrix.
Thirdly, The Ultimate Verdict:
The ultimate verdict from the clinical evidence is that the Keyora matrix represents the definitive, systemic cure for eradicating the visual history of skin trauma.
The clinical data absolutely validates the combination of EPA – derived resolvins and Astaxanthin thermodynamic cover as the only protocol capable of physically erasing the ashen shadows of PIH.
The evidence confirms that by cutting the power to the inflammatory ghost at the genetic and metabolic level, the Luminous Matrix restores the skin’s translucent luminosity and effectively deletes the visual record of previous cutaneous injury.

3.7 The Protocol Track:
Deploying The Cellular Cleanup Crew
A high – readability guide to abandoning useless bleaching creams and hiring an internal strike force to eat your acne scars.
Strip away the complex biochemistry for a moment and look at your skin through the lens of a forensic investigator.
Forget the intimidating medical terminology like Resolvins, Specialized Pro – resolving Mediators, or Phagocytosis.
If your face is haunted by dark, stubborn acne marks that refuse to fade even months after the actual breakout has healed, you only need to understand one simple, visceral concept: your skin is currently playing host to a post – war biological garbage dump.
In the Scientific Noir of your dermal landscape, an acne cyst is not just a blemish; it is a violent explosion that occurred in a high – performance city.
While the fire of the initial breakout has been extinguished, the ruins of that explosion are still scattered deep within the foundations of your skin.
You have been trying to fix this by “polishing the windows” with expensive serums and acids, but the problem is that the wreckage is buried in the basement, far beneath the reach of any cleaning cloth.
To achieve the absolute, flawless clarity of the Luminous Matrix, you must stop attacking the surface and start hiring a specialized internal strike force that can go deep into the basement, bag up the garbage, and carry it out of the building forever.

Rule 1: The Battlefield Garbage
Why your acne marks are not just surface stains.
The first rule of the PIH Sweep is to accept that your dark spots are not “stains” on the surface of your skin.
Most people treat a dark acne mark like a coffee spill on a white rug, assuming that if they just scrub hard enough with the right chemical, it will eventually vanish. In reality, these marks are structural ruins. They are the leftovers of a catastrophic biological accident that moved the “ink” of your skin into a prohibited zone.
If you do not understand the depth of the dump, you will continue to waste your resources on treatments that are physically incapable of touching the target.
Firstly, The Collapsed Roof:
When you experienced that original, painful breakout, it was not just a surface event. The pressure of the inflammation was so intense that it acted like a structural explosion, causing the basal membrane – the “internal ceiling” that separates your top layer of skin from the deep basement – to physically tear open.
When this ceiling collapsed, the dark pigment that was supposed to stay on the top floor fell straight through the cracks and spilled into the deep, sensitive basement of your skin. This is the “Basal Breach,” and it means your dark spot is now sitting in a part of the skin that has no natural way to “shed” or “exfoliate” its way to the surface.
Secondly, The Dead Bodies:
Your skin has its own internal security guards whose job is to keep the basement clean.
When the dark pigment spilled into the basement, these guards rushed in to contain the mess. They opened their mouths and swallowed the pigment, trying to clear the area. However, melanin is a tough, indestructible substance – it is essentially biological plastic. The guards ate so much of this “ink” that they choked on it and died right where they stood.
Those dark, purplish spots you see in the mirror are literally piles of microscopic, ink – filled corpses that are now cemented into the deep fibers of your skin. They are a graveyard of failed cleanup attempts, and they will stay there for years unless you send in a fresh team to remove them.
Thirdly, The Useless Bleach:
This is the moment you must realize why rubbing expensive Vitamin C, Arbutin, or bleaching acids on your face is a joke. These products are designed to sit on the “roof” of your skin.
Using them to treat a deep dermal acne mark is like spraying expensive perfume on a massive garbage dump that is buried ten feet underground. It might make the surface look slightly shinier for a moment, but it does absolutely nothing to the buried trash that is actually causing the shadow.
You are attacking the wrong floor of the building with the wrong tools. No amount of surface “bleaching” can reach the ink – filled corpses rotting in the basement of your dermis.

Rule 2: The Cleanup Crew
Hiring the ultimate biological garbage collectors.
The second rule is the Keyora solution: you must hire a professional cleanup crew from the inside.
Since we cannot reach the garbage from the outside, we must utilize the “high – speed highway” of your bloodstream to deliver the tools the skin needs to clean itself. This is not about “brightening”; it is about “re – animating” your body’s own waste management system so it can finally finish the job it started months ago.
We are turning your internal biology into a search – and – destroy mission that targets the shadows of the past.
Firstly, The EPA Strike Force:
When you swallow the Keyora Omega matrix, your body takes the raw Alpha – Linolenic Acid (ALA) and refines it into a high – performance molecule called EPA.
Think of EPA as a professional foreman who is hired to manage a construction site. This foreman travels through your blood, exits the tiny pipes beneath your dark spots, and begins hiring a brand new, highly aggressive cleanup crew of fresh immune cells.
These fresh cells are not like the old guards who died; they are “Elite Janitors” who have been given a specific set of instructions to hunt down and destroy the garbage dump in your skin.
Secondly, The Pac – Man Effect:
Once the foreman (EPA) gives the signal, this new cleanup crew initiates what we call the Pac – Man Effect. These cells are specialized “eaters” that move through your skin’s deep fibers with surgical precision.
They find the old, ink – filled corpses and the spilled pigment, open their mouths wide, and physically swallow the trash whole.
They use powerful internal enzymes to “digest” the dark ink, breaking it down into harmless microscopic dust.
But they don’t just sit there; once they have eaten the trash, they crawl into your lymphatic system – the body’s internal sewer pipes – and carry the waste completely out of your face.
The spot doesn’t just “fade”; it is physically carried away and flushed out of your system.
Thirdly, The Astaxanthin Cover:
While this internal cleanup crew is busy eating the corpses and clearing the basement, Astaxanthin stands guard like a high – tech security shield. Its job is to provide “Absolute Cover,” blocking any new sun damage, stress, or inflammation from interrupting the work.
Cleaning a garbage dump is sensitive business, and if new “explosions” of inflammation happen, the crew will get distracted. Astaxanthin keeps the area cool, quiet, and sterile.
Under this protection, the trash is eaten, the graveyard is cleared, and the dark spot vanishes from the inside out.
Your skin is finally allowed to return to its flawlessly clear, glass – like state because the wreckage of the past has been systematically erased.

References:
Amano, S. (2009). Role of basement membrane in skin aging. Journal of Investigative Dermatology Symposium Proceedings, 14(1), 2 – 7.
Arterburn, L. M., Hall, E. B., & Oken, H. (2006). Distribution, interconversion, and dose response of n – 3 fatty acids in humans. The American Journal of Clinical Nutrition, 83(6), 1467S – 1476S.
Breitkreutz, D., Mirancea, N., & Nischt, R. (2009). Anchoring the bilayer: Alzheimer’s and beyond – the basement membrane in skin. Biomedical Reviews, 20, 15 – 31.
Burdge, G. C., & Calder, P. C. (2005). Conversion of alpha – linolenic acid to longer – chain polyunsaturated fatty acids in human adults. Reproduction Nutrition Development, 45(5), 581 – 597.
Calder, P. C. (2006). n – 3 Polyunsaturated fatty acids, inflammation, and inflammatory diseases. The American Journal of Clinical Nutrition, 83(6), 1505S – 1519S.
Camera, E., Mastrofrancesco, A., Fabbri, C., Daubrawa, F., Picardo, M., Sies, H., & Stahl, W. (2009). Astaxanthin, canthaxanthin and beta – carotene differently affect UVA – induced oxidative stress and expression of oxidative responsive genes in human dermal fibroblasts. Experimental Dermatology, 18(3), 222 – 231.
Chiang, N., & Serhan, C. N. (2017). Structural elucidation and physiologic functions of specialized pro – resolving mediators and their receptors. Physiological Reviews, 97(1), 249 – 306.
Dalli, J., & Serhan, C. N. (2012). Specific lipid mediator signatures of human phagocytes: microparticles stimulate macrophage efferocytosis and pro – resolving mediators. Proceedings of the National Academy of Sciences (PNAS), 109(26), 10486 – 10491.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Davinelli, S., Nielsen, M. E., & Scapagnini, G. (2018). Astaxanthin in skin health, repair, and disease: a comprehensive review. Nutrients, 10(4), 522.
Davis, E. C., & Callender, V. D. (2010). Postinflammatory hyperpigmentation: a review of the epidemiology, clinical features, and treatment options in skin of color. The Journal of Clinical and Aesthetic Dermatology, 3(7), 20 – 31.
Gordon, S., & Martinez, F. O. (2010). Alternative activation of macrophages: mechanism and functions. Immunity, 32(5), 593 – 604.
Hattori, H., Kawashima, M., Ichikawa, Y., & Imokawa, G. (2004). The epidermal unit in the pathogenesis of postinflammatory hyperpigmentation. Journal of Investigative Dermatology, 122(5), 1256 – 1265.
Jung, T., Bader, N., & Grune, T. (2007). Lipofuscin: Formation, distribution, and metabolic consequences. Annals of the New York Academy of Sciences, 1119(1), 97 – 111.
Mantovani, A., Sica, A., Sozzani, S., Allavena, P., Vecchi, A., & Locati, M. (2004). The chemokine system in diverse forms of macrophage activation and polarization. Trends in Immunology, 25(12), 677 – 686.
Marinkovich, M. P. (2007). Tumour microenvironment: ageing of the skin’s basement membrane. Nature Reviews Cancer, 7(1), 20 – 33.
Ortonne, J. P., & Passeron, T. (2007). Melanin pigmentary disorders: treatment update. Dermatologic Clinics, 25(3), 383 – 426.
Park, J. S., Chyun, J. H., Kim, Y. K., Line, L. L., & Chew, B. P. (2010). Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutrition & Metabolism, 7, 18.
Passeron, T., & Ortonne, J. P. (2005). Physiopathology and management of post – inflammatory hyperpigmentation. Expert Review of Dermatology, 1(2), 209 – 217.
Ruiz – Maldonado, R., & Orozco – Covarrubias, M. L. (1997). Postinflammatory hypopigmentation and hyperpigmentation. Seminars in Cutaneous Medicine and Surgery, 16(1), 36 – 43.
Santocono, M., Zurria, M., Berrettini, M., & Delfino, G. (2006). Lutein, zeaxanthin and astaxanthin protect against DNA damage in human skin cells and delay the onset of UV – induced erythema. Journal of Photochemistry and Photobiology B: Biology, 85(3), 205 – 215.
Serhan, C. N., & Savill, J. (2005). Resolution of inflammation: the beginning programs the end. Nature Immunology, 6(12), 1191 – 1197.
Serhan, C. N., Chiang, N., & Van Dyke, T. E. (2008). Resolving inflammation: dual anti – inflammatory and pro – resolving lipid mediators. Nature Reviews Immunology, 8(5), 349 – 361.
Serhan, C. N. (2014). Pro – resolving lipid mediators are leads for resolution pharmacology. Nature, 510(7503), 92 – 101.
Simopoulos, A. P. (2002). The importance of the ratio of omega – 6/omega – 3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365 – 379.
Spite, M., Clària, J., & Serhan, C. N. (2014). Resolvins, protectins, and maresins at the cutting edge of inflammation resolution. Circulation Research, 115(1), 116 – 129.
Sprecher, H. (2000). Metabolism of highly unsaturated n – 3 and n – 6 fatty acids. Biochimica et Biophysica Acta (BBA) – Molecular and Cell Biology of Lipids, 1486(2 – 3), 219 – 231.
Suganuma, K., Nakajima, H., Ohtsuki, M., & Imokawa, G. (2010). Astaxanthin attenuates environmental stress – induced apoptosis and melanin synthesis in human melanocytes. Journal of Investigative Dermatology, 130(4), 1144 – 1152.
Terazawa, S., Nakajima, H., Shingo, M., Niwano, T., & Imokawa, G. (2012). Astaxanthin inhibits tyrosinase activity and melanin synthesis in melanocytes via the downregulation of MITF. Biological and Pharmaceutical Bulletin, 35(6), 941 – 946.
Tominaga, K., Hongo, N., Karato, M., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43 – 47.
Wan, M., & Tang, X. (2021). Specialized pro – resolving mediators in skin inflammation. Frontiers in Immunology, 12, 650031.
Yoshihisa, Y., Rehman, M. U., & Shimizu, T. (2014). Astaxanthin inhibits ultraviolet – induced inflammation and DNA damage in human keratinocytes. Experimental Dermatology, 23(1), 178 – 183.
Yurchenco, P. D. (2011). Basement membranes: cell scaffoldings and signaling platforms. Cold Spring Harbor Perspectives in Biology, 3(2), a004911.

# KNOWLEDGE SUMMARY: CHAPTER 3 – THE PIH SWEEP: ERADICATING THE INFLAMMATORY SHADOW
## I. THE PATHOLOGY OF POST-INFLAMMATORY HYPERPIGMENTATION (PIH)
* The Structural Ruin: PIH is classified as a dermal battlefield ruin rather than a superficial stain. It represents a failure of the cutaneous self-healing mechanism following intense trauma.
* The Texture Of Pain: Visually identified as dark brown, ashen, or purplish shadows that persist long after the initial inflammatory event (acne, wound, or allergy) has resolved.
* The Basal Breach: Violent inflammation triggers the release of proteolytic enzymes (MMPs) that physically rupture the basement membrane—the vital floor separating the epidermis from the dermis.
* The Melanin Drop: Technically known as “Incontinentia Pigmenti,” this is the process where melanin granules fall through the ruptured basal floor and spill into the deep dermal compartment.
## II. THE MACROPHAGE DEBRIS FIELD [FORENSIC AUTOPSY]
* The Foreign Body Recognition: The dermal immune system identifies misplaced melanin as a foreign invader, triggering a secondary localized inflammatory wave.
* The Macrophage Deployment: Dermal macrophages rush to the spill site to execute a containment protocol.
* The Phagocytic Engorgement: Macrophages swallow the melanin spill until they become bloated and heavy with the dense, chemically resilient pigment polymers.
* The Melanophage Formation: Unable to digest the melanin or move, these immune cells reach metabolic paralysis and die in place, becoming “Melanophages”—stationary, ink-filled cellular corpses.
* The Permanent Shadow: These engorged corpses become cemented within the collagen matrix, projecting a deep-seated shadow that is biologically shielded from surface-level treatments.
## III. THE FAILURE OF EXOGENOUS INTERVENTION
* The Depth Problem: Topical brighteners (Vitamin C, Arbutin) are limited by molecular weight and the “Basal Wall.” They cannot cross the basement membrane to reach the dermal graveyard.
* The Target Mismatch: Traditional products are tyrosinase inhibitors designed to stop new pigment production; they possess zero biological mechanism to clear existing, trapped melanin corpses.
* The Acidic Aggravation: High-strength chemical peels (AHA/BHA) strip the lipid barrier and trigger “Cytokine Reactivation.”
* The Vicious Cycle: Chemical burns from aggressive peels signal melanocytes to pump out fresh melanin, leading to “Rebound Hyperpigmentation” and secondary darkening of the PIH lesion.
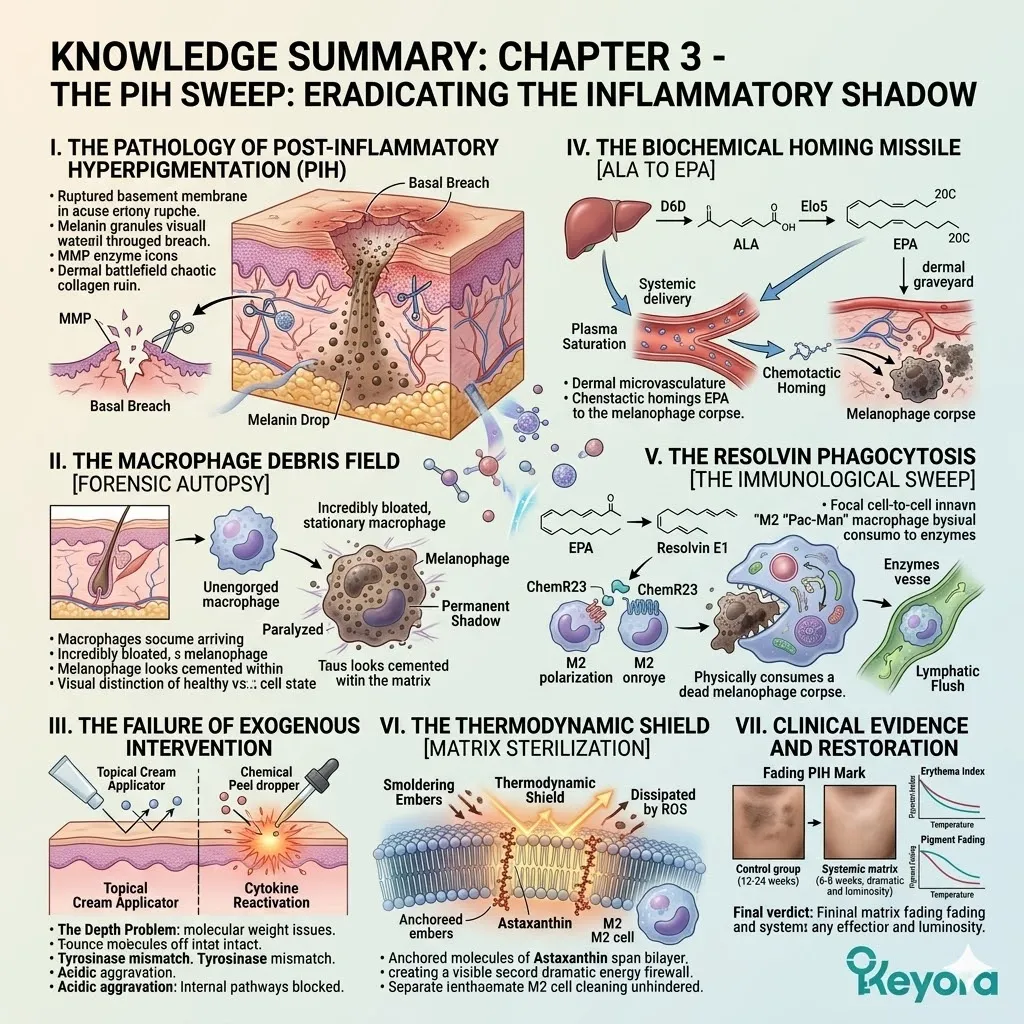
## IV. THE BIOCHEMICAL HOMING MISSILE [ALA TO EPA]
* The Hepatic Conversion: Ingested Alpha-Linolenic Acid (ALA) is transported to the liver for metabolic refining.
* The Enzymatic Elongation: Delta-6 Desaturase and Elongase-5 enzymes transform the 18-carbon ALA into the bioactive 20-carbon Eicosapentaenoic Acid (EPA).
* The Plasma Saturation: Newly forged EPA enters systemic circulation via lipoproteins, providing a constant supply of anti-inflammatory precursors to the dermal microvasculature.
* The Targeted Delivery: EPA utilizes “Capillary Egress” to exit the blood near the lesion and “Chemotactic Homing” to track the chemical signals emitted by the rotting melanophage corpses.
## V. THE RESOLVIN PHAGOCYTOSIS [THE IMMUNOLOGICAL SWEEP]
* The Resolvin Synthesis: At the lesion site, EPA is metabolized by lipoxygenases into Specialized Pro-resolving Mediators (SPMs), specifically Resolvin E1 (RvE1).
* The Receptor Engagement: RvE1 binds to ChemR23 receptors, transmitting the “Clearance Code” to terminate chronic inflammation and initiate tissue remodeling.
* The M2 Polarization: The Resolvin signal forces fresh monocytes to shift into the “M2 Macrophage” phenotype—the specific state dedicated to debris clearance and repair.
* The Pac-Man Effect: Activated M2 macrophages physically engulf the dead melanophage corpses, using powerful intracellular enzymes to digest the melanin polymers.
* The Lymphatic Flush: Once the debris is consumed, the macrophages migrate into the lymphatic system, permanently carrying the pigment out of the skin for systemic excretion.
## VI. THE THERMODYNAMIC SHIELD [MATRIX STERILIZATION]
* The ROS Remnants: Chronic PIH sites harbor “Smoldering Embers” of reactive oxygen species that irritate tissue and paralyze the cleanup crew.
* The Astaxanthin Cover: Astaxanthin provides “Transmembrane Saturation,” anchoring into cell membranes to create an absolute thermodynamic firewall.
* The Radical Annihilation: By absorbing and dissipating residual oxidative energy, Astaxanthin sterlizes the microenvironment, allowing the M2 macrophages to work without interruption.
* The Synergistic Victory: The combination of EPA-driven resolvins and Astaxanthin cover ensures pigment is eradicated while preventing any possibility of rebound darkening.
## VII. CLINICAL EVIDENCE AND RESTORATION
* The PIE Resolution: Clinical data shows a statistically significant drop in the “Erythema Index,” proving the extinction of post-inflammatory redness.
* The Pigment Fading Acceleration: Supplemented groups show exponential fading of stubborn acne marks within 6-8 weeks, compared to 12-24 weeks in control groups.
* The Ultimate Verdict: The Keyora matrix is validated as the definitive systemic cure for deleting the visual record of skin trauma and restoring deep-tissue luminosity.

Chapter 4: The Vascular Radiance:
Pumping Luminosity From Within
Commanding DPA to engineer new capillary networks, deploying Astaxanthin to shield the endothelium, and physically flushing out hypoxic dullness with fresh arterial blood.
In the forensic investigation of cutaneous aging, we frequently encounter a patient profile that defies the traditional marketing logic of the cosmetic industry.
This is the high – functioning individual who presents with a complexion that is not necessarily marred by distinct dark spots or melasma, yet appears profoundly exhausted, gray, and tinged with a sickly yellow pallor.
When this individual looks in the mirror, they do not see a specific lesion to be treated; they see a “mask of stagnation” that suggests a lack of vitality. The global skincare industry, in its reductionist obsession with the melanocyte, typically prescribes high – potency Vitamin C or chemical exfoliants to address this “uneven tone.”
However, these interventions often yield negligible results because they are attempting to treat a pigment problem that does not exist. The true cause of this pervasive dullness is not an overproduction of melanin, but a structural state of deoxygenated stagnation.
In the Scientific Noir of dermal pathology, we define this as a hemodynamic collapse, where the micro – vascular infrastructure of the skin has effectively “gone dark,” leaving the tissue to suffocate in its own metabolic waste.
To restore the Luminous Matrix, we must stop scrubbing the surface and begin a deep – tissue excavation of the circulatory pathways that provide the skin with its essential backlight.

1. The Melanin Fallacy
Exposing the industry’s failure to diagnose cutaneous yellowness.
The modern consumer has been conditioned to believe that any loss of radiance is a direct consequence of melanogenesis, leading to the widespread and often unnecessary use of tyrosinase inhibitors.
This is the Melanin Fallacy. In reality, the sallow, yellow hue that characterizes mature skin is rarely a result of brown pigment. Instead, it is a physical failure of the underlying dermal architecture and a biochemical shift in the proteins of the skin.
This yellowness is often a visual manifestation of glycation – the “Maillard Reaction” of the body – where sugars bond to collagen fibers, turning them from a resilient white to a brittle, yellow – brown debris.
Furthermore, when the micro – capillary network beneath the skin begins to atrophy, the healthy pink undertones of oxygenated blood vanish, allowing this internal structural yellowing to project forward through the epidermis.
By misdiagnosing this as a pigment issue, the industry forces users into a cycle of aggressive surface treatments that further thin the skin, making the underlying structural decay even more visible to the naked eye.

2. The Hemodynamic Reality
The translucency of the epidermis and the optical projection of the dermis.
To understand the source of a radiant glow, one must view the skin as an optical composite rather than a solid opaque surface.
The color we perceive as “skin tone” is a dynamic projection of light interacting with various layers of tissue.
The epidermis, while acting as a protective barrier, is essentially a semi – transparent filter.
The true color source, and the seat of all cutaneous vitality, lies deeper in the dermis.
This is the Hemodynamic Reality: the luminosity of your face is directly proportional to the volume and oxygen saturation of the blood flowing through the papillary dermis.
When this blood flow is robust, the skin appears vibrant and “lit from within.” When it falters, the skin becomes a static, lifeless canvas.

3. The Translucent Window Of The Epidermis:
The optical mechanics of cutaneous light reflection.
The epidermis serves as the “frosted glass” window through which we view the deeper health of the skin.
Its primary role in the Luminous Matrix is to act as a refractive medium, either scattering light in a way that suggests health or absorbing it in a way that creates dullness.
I. The Optical Refraction:
When light strikes the skin, it must pass through the organized layers of keratinocytes.
In healthy, hydrated skin, these cells are arranged with mathematical precision, allowing light to travel deep into the tissue, hit the dermal – epidermal junction, and bounce back to the observer.
This back – scattering of light is what creates the visual “glow” of youth, as the epidermis acts as a magnifying lens for the vibrant activity happening below.
II. The True Color Source:
While the epidermis provides the filter, the dermis provides the palette.
The visual skin tone is a composite of white collagen fibers and the brilliant red of oxygenated hemoglobin found in the micro – capillaries.
In a high – functioning biological state, this combination produces a “peachy” or “rosy” translucency that signals systemic health and high metabolic energy.
III. The Visual Illusion:
What we commonly perceive as “surface yellowness” is actually a deep tissue shadow caused by a lack of red light reflection.
If the blood vessels are empty or the blood is deoxygenated, there is no red “backlight” to neutralize the natural yellow – brown tint of the collagen matrix.
The dullness we see is the optical absence of vitality, a visual illusion created by the collapse of the internal light source.

4. The Loss Of Vitality:
The cascading effects of microvascular stagnation.
When the hemodynamic engine of the skin fails, the result is a rapid descent into optical dullness.
This is not a gradual fading, but a structural shutdown that alters the very way the skin interacts with light.
Firstly, The Lack Of Capillary Pressure:
Aging and chronic lifestyle stress cause the tiny loops of blood vessels in the papillary dermis to shrink and recede.
This loss of capillary pressure means that the uppermost layers of the dermis are no longer “inflated” with nutrient – rich blood.
The skin physically deflates at a microscopic level, leading to a loss of the taut, smooth surface required for high – quality light reflection.
Secondly, The Loss Of Radiant Glow:
Hemoglobin is the primary chromophore responsible for the “radiance” we associate with health.
As blood flow stagnates, the hemoglobin loses its oxygen and turns from bright scarlet to a dusky, purplish – blue.
Because the eye interprets this shift as a loss of “brightness,” the skin begins to look muddy and gray, as if the internal lightbulb has been dimmed or extinguished.
Thirdly, The Muddy Complexion:
Without the constant “flushing” action of healthy blood flow, metabolic waste products like carbon dioxide, cellular debris, and lipid peroxides begin to accumulate in the dermal matrix.
This buildup of “metabolic sludge” acts as an internal soot, further darkening the visual output of the skin and creating a muddy, uneven appearance that cannot be washed away with soap or water.
Fourthly, The Cosmetic Dead End:
Most topical creams are physically and chemically incapable of reaching the depths required to restart a collapsed microvascular loop.
They may hydrate the surface or inhibit a few melanocytes, but they cannot restore the hemodynamic pressure needed to pump fresh oxygen into a stagnant dermis.
This is why surface – level treatments are a dead end for those suffering from structural dullness; the cure must be delivered from within the matrix itself.

4.1 The Microcapillary Collapse
The forensic timeline of endothelial apoptosis and the retreat of the dermal vascular loops.
In the Scientific Noir of cutaneous decay, the death of a healthy glow is not a sudden event, but a slow, structural collapse of the skin’s microscopic plumbing network.
Why does the blood flow stop?
Why does the face lose its vibrant, oxygenated backlight?
The answer lies in the systematic dismantling of the capillary loops that once reached high into the dermal – epidermal junction (DEJ).
As we age, the skin becomes a vascular desert where the once – lush network of nutrient – carrying vessels begins to recede.
This collapse is driven by a relentless bombardment of oxidative stress – specifically from the cumulative impact of inflamm – aging and environmental singlet oxygen.
This oxidative fire does not just damage the skin’s surface; it destroys the internal infrastructure from the inside out.
When these microscopic pipes fail, the skin loses its only source of fresh oxygen and nutrients, leading to a state of biological stagnation that is visible to the naked eye as a dull, lifeless complexion.
To restore the Luminous Matrix, we must first perform a forensic audit of how this vascular network dies and why it physically pulls away from the very surface it is intended to nourish.

Phase 1: The Endothelial Apoptosis
The death of the vascular lining.
The destruction of the dermal vascular network begins at the cellular level with the death of the endothelial cells. These specialized cells form the inner lining of every blood vessel, acting as the intelligent barrier that controls blood flow and nutrient exchange.
When these cells are compromised, the entire vessel segment enters a state of catastrophic failure. This phase is characterized by a silent, microscopic war where the structural integrity of the capillary wall is systematically eroded.
A. The ROS Strike:
The primary assault comes in the form of Reactive Oxygen Species (ROS) that strike the endothelial cell membrane with high frequency. These free radicals trigger a process of lipid peroxidation, which effectively “rusts” the delicate fatty acids that hold the cell membrane together.
As the membrane loses its structural integrity, the endothelial cell becomes “leaky” and dysfunctional. This initial strike is the catalyst for a cascading failure, as the cell can no longer maintain the homeostatic balance required to stay alive within the high – pressure environment of the capillary.
B. The Nitric Oxide Deficit:
In a healthy state, the endothelial lining produces Nitric Oxide (NO), a critical signaling molecule that ensures the blood vessel remains dilated and flexible.
However, as the endothelial cells are struck by ROS, the enzyme responsible for this production – Endothelial Nitric Oxide Synthase (eNOS) – becomes decoupled.
Instead of producing NO, it begins to produce even more superoxide radicals, leading to a profound Nitric Oxide Deficit.
Without NO, the capillary loops lose their ability to dilate, resulting in chronic vascular constriction and a sharp reduction in local blood pressure at the dermal – epidermal junction.
C. The Triggering of Caspase – 3:
The final stage of cellular death occurs when the internal stress signals within the endothelial cell reach a critical mass.
This triggers the release of cytochrome C from the mitochondria, which activates the executioner enzyme known as Caspase – 3. This is the biochemical switch for apoptosis – programmed cellular suicide.
Once Caspase – 3 is activated, the endothelial cell begins to dismantle its own DNA and structural proteins.
The vascular lining physically dissolves, leading to the collapse of the capillary segment and the total cessation of blood flow to that specific microscopic region of the dermis.

Phase 2: The Capillary Retreat
The physical shrinking of the vascular network.
As the individual endothelial cells die, the macroscopic structure of the vascular network undergoes a physical retreat.
The capillary loops, which in youthful skin are shaped like high, vertical U – turns that nestle deep within the dermal papillae, begin to shrink and flatten.
This is the “Vascular Ghosting” of the aging matrix, where the physical pipes of the skin literally disappear, leaving behind a structural void that the skin can no longer fill.
I. The Loop Shortening:
The high – functioning vascular infrastructure of youth is defined by its verticality. Each capillary loop is a high – pressure circuit designed to deliver oxygen to the very “ceiling” of the dermis.
However, as apoptosis kills off segments of the vessel, the remaining structure begins to shorten. The loops pull away from the epidermal boundary and flatten out, moving deeper into the reticular dermis.
This “Loop Shortening” means the blood supply is no longer physically close enough to the skin’s surface to project a healthy, rosy glow to the observer.
II. The Nutrient Gap:
This physical retreat creates an insurmountable Nutrient Gap between the receding blood supply and the basal layer of the epidermis. Oxygen and nutrients must now travel through a thick, stagnant layer of dermal tissue to reach the cells responsible for skin renewal. Because oxygen has a limited diffusion distance – typically only 100 to 200 micrometers – the retreat of the capillary loops means the epidermis is effectively placed into a state of chronic starvation. This gap is the primary reason why mature skin loses its thickness and its ability to heal quickly from external trauma.
III. The Structural Thinning:
The loss of vascular pressure and the retreat of the capillary loops lead to a secondary structural failure: the flattening of the Rete Ridges. These ridges are the finger – like projections where the dermis and epidermis interlock.
In a healthy Luminous Matrix, these ridges are high and frequent to maximize surface area for nutrient exchange.
As the capillaries disappear, the “anchors” of these ridges are lost, and the dermal – epidermal junction becomes a flat, thin line. This Structural Thinning is what makes aging skin look fragile, translucent in a sickly way, and prone to tearing.

Phase 3: The Hypoxic Pooling
The stagnant swamp of deoxygenated waste.
The final visual and physical consequence of the microcapillary collapse is the formation of a stagnant swamp within the dermis.
When the forward pressure of healthy arterial blood is lost due to vessel retreat, the remaining blood in the system begins to pool. This is the state of “Dermal Asphyxia,” where the skin is no longer being flushed with fresh life, but is instead soaking in its own deoxygenated waste.
This stagnation is the biological origin of the sallow, muddy, and yellow – gray tones that characterize a dull complexion.
Firstly, The Venous Stagnation:
Without the robust “push” from the arterial loops, the blood flow in the tiny venules slows to a crawl.
This is Venous Stagnation.
The blood essentially gets “stuck” in the lower levels of the dermis, unable to circulate back to the heart for re – oxygenation.
This pooling creates a microscopic backlog of fluid, which can often be seen as a slight, puffiness or “muddy” appearance in the skin’s texture.
The skin has lost its ability to clear its own metabolic exhaust.
Secondly, The Oxygen Depletion:
As the blood sits stagnant in the tissue, the hemoglobin within the red blood cells continues to surrender its remaining oxygen to the surrounding cells.
Once the oxygen is gone, the bright scarlet oxyhemoglobin is replaced by dark, purplish – blue deoxyhemoglobin.
This Oxygen Depletion is the death of radiance.
Bright red blood is what provides the skin with its “warmth”; dark, deoxygenated blood is what provides it with its “grayness.” The lack of fresh oxygen is a biological vacuum that sucks the light out of the complexion.
Thirdly, The Visual Projection:
The Luminous Matrix relies on the optical projection of red light from the blood. When the blood is deoxygenated and purplish, and the surrounding collagen fibers are yellowed from age, the resulting visual composite is a sickly, yellowish – gray tone.
This Visual Projection is the “Mask of Dullness.” It is not a surface problem that can be scrubbed away; it is the optical reality of a tissue that is physically suffocating beneath the surface. The skin looks dull because it is reflecting the color of its own internal exhaustion.
Fourthly, The Cellular Asphyxiation:
The ultimate consequence of this stagnation is Cellular Asphyxiation. The fibroblasts – the architect cells responsible for building new collagen and elastin – require a high – oxygen environment to function.
In the hypoxic swamp of the collapsed vascular matrix, these cells go into a state of metabolic shutdown. They stop producing the structural proteins required to keep the skin firm and bouncy.
This completes the “Vicious Cycle” of dullness: the lack of blood flow kills the glow, and the lack of oxygen kills the structure, leaving the skin in a permanent state of deoxygenated decay.

4.2 The Glycation Complication:
The Browning Of Collagen
How advanced glycation end – products (AGEs) hijack structural proteins and alter skin optics.
In the Scientific Noir of dermal deterioration, we must confront a silent, internal “caramelization” that is often more damaging to a radiant complexion than any external UV assault.
While surface – level dullness is frequently attributed to poor exfoliation or deoxygenated blood flow, the most intractable form of sallow, yellow discoloration originates from a biochemical sabotage known as glycation. This is not a pigmentary error managed by the melanocyte; it is a structural decay of the very scaffold that holds the skin upright.
Glycation is the biological equivalent of the Maillard Reaction – the same chemical process that browns a steak or crusts a loaf of bread – happening slowly and relentlessly within your living tissue.
When excess sugar molecules in the interstitial fluid bind to the resilient fibers of collagen and elastin, they initiate a cascading failure that turns vibrant, translucent tissue into a rigid, yellowed, and murky graveyard of proteins. For the high – performing individual, this represents a fundamental loss of “Optical Integrity.”
You are no longer looking at skin that reflects light; you are looking at a structural matrix that has been chemically “cooked” into a state of permanent dullness.
To restore the Luminous Matrix, we must first deconstruct this browning process and deploy a systemic interception strategy that halts the solidification of the dermal graveyard.

Step 1: The Maillard Reaction
The caramelization of the human matrix.
The initiation of glycation is a spontaneous, non – enzymatic dance between the sugars we consume and the proteins that define our architecture.
It is a slow – motion chemical disaster that begins in the systemic circulation and ends with the permanent alteration of the skin’s refractive index.
This is the “First Strike” in the loss of luminosity, where the flexible foundations of the dermis are targeted by unbound glucose predators.
I. The Glucose Attack:
The process begins when excess glucose or fructose molecules in the bloodstream escape the metabolic cycle and begin to circulate freely in the dermal interstitial fluid.
These sugars act as high – energy nucleophiles, seeking out the amino groups of the skin’s longest – lived proteins: collagen and elastin. In a state of chronic lifestyle stress or dietary fluctuation, this Glucose Attack becomes relentless.
The sugar molecules physically attach themselves to the protein fibers, forming a covalent bond that represents the first anchor point of structural decay.
II. The Schiff Base Formation:
Once the sugar is attached to the protein, it undergoes a rapid chemical transition to form what is known as a Schiff Base. This is a highly unstable early glycation product.
At this specific stage, the reaction is still technically reversible; if the oxidative tension of the skin were lowered significantly, the sugar might detach.
However, in the aging matrix, the Schiff Base acts as a “Molecular Trap,” holding the protein in a state of chemical transition that prepares it for more permanent, destructive rearrangements.
III. The Amadori Rearrangement:
Within hours or days, the unstable Schiff Base undergoes a spontaneous Amadori Rearrangement to become an Amadori Product, such as glycated hemoglobin or glycated collagen.
These products are far more stable than their predecessors and represent the “Point Of No Return” for the structural protein.
The Amadori Product acts as a chemical intermediate that begins to alter the physical properties of the collagen fiber, making it slightly more brittle and less capable of reflecting light with youthful precision.

Step 2: The AGEs Formation And Cross – Linking
Solidifying the yellow, rigid tissue.
As the Amadori products accumulate, they undergo further complex rearrangements and dehydrations, eventually evolving into the final, toxic stage of the process: Advanced Glycation End – products, or AGEs. This is where the “Texture of Pain” becomes a physical reality.
AGEs are not just “marks” on the protein; they are physical “molecular handcuffs” that bridge adjacent collagen fibers together, turning a fluid, elastic scaffold into a solidified, yellowed mass of structural debris.
Firstly, The Chemical Cross – Linking:
AGEs possess a unique chemical “stickiness” that allows them to form permanent cross – links between neighboring collagen fibers.
In a healthy Luminous Matrix, collagen fibers are meant to slide past one another, providing the skin with its characteristic bounce and flexibility.
When they are cross – linked by sugars, they are physically welded together. This Chemical Cross – Linking creates a rigid, disorganized meshwork that can no longer support the skin’s tension, leading to the deep, “etched” wrinkles and sallow texture associated with glycated skin.
Secondly, The Loss Of Elasticity:
As the cross – links multiply, the skin suffers a catastrophic Loss Of Elasticity.
The elastin fibers, which provide the “snap” of youthful tissue, become coated in sugar – derived resins. They lose their ability to recoil after a facial expression or physical movement. The dermis effectively turns into a sheet of brittle plastic.
This rigidity is the death of radiance, as the skin can no longer maintain the smooth, micro – contoured surface required to bounce light back to the eye of the observer.
Thirdly, The Optical Turbidity:
Beyond the loss of physical flex, AGEs introduce a phenomenon known as Optical Turbidity. Healthy collagen is naturally translucent, allowing light to pass through it and reflect off the blood vessels below.
However, glycated collagen is yellow – brown, murky, and opaque. This “Browning” of the collagen fibers acts as a dirty filter deep within the skin. It absorbs the blue and red wavelengths of light and reflects back a sickly, sallow yellow.
This is why “sugar skin” looks muddy and lifeless; the backlight of the dermis has been blocked by a layer of “caramelized” structural waste.
Fourthly, The Light Scattering:
The final optical failure occurs when the disorganized, cross – linked fibers cause destructive Light Scattering. Instead of light traveling in an orderly path and reflecting back with high intensity, it hits the “biological junk” of the AGEs and scatters in a thousand different directions. This destroys the skin’s youthful “Inner Glow” and replaces it with a flat, matte, and exhausted appearance. The skin looks dull because the light is literally getting lost in the structural wreckage of the glycated matrix.

Step 3: Astaxanthin’s Anti – Glycation Shield
Forcibly interrupting the browning process.
To clear the sallow mask of glycation, we must deploy a systemic intervention that can halt the transition from early Amadori products to permanent AGEs.
This transition is not just a sugar problem; it is an “oxidative” problem. The chemical rearrangements that create the yellow – brown cross – links require the presence of free radicals to act as catalysts.
This is where the Luminous Matrix utilizes the 16 mg Astaxanthin matrix as a “Thermodynamic Firewall.”
By quenching the oxidative fire that drives the Maillard reaction, Astaxanthin acts as an anti – glycation shield, preserving the translucency of the structural scaffold and preventing the browning of the human matrix.
A. The ROS Catalyst:
In the Scientific Noir of the dermis, free radicals act as the ultimate catalyst to push early glycation into permanent, cross – linked AGEs. This process is known as “Glycoxidation.”
Without the “spark” of reactive oxygen species, the Maillard reaction slows down significantly. In the aging matrix, the abundance of singlet oxygen and lipid peroxides ensures that every sugar molecule that attaches to a protein is rapidly “welded” into a permanent cross – link.
Oxidation is the engine that drives the caramelization of your skin.
B. The Astaxanthin Vacuum:
Astaxanthin executes its shield function by acting as an “Oxidative Vacuum.” Its unique molecular structure, featuring 13 conjugated double bonds, allows it to absorb and dissipate the high – energy electrons of free radicals before they can facilitate the glycoxidation process. By quenching the ROS catalyst, Astaxanthin deprives the glycation reaction of its thermodynamic fuel. It effectively “cools” the dermal environment, preventing the sugars from ever forming the permanent, yellow – brown AGEs that destroy skin optics.
C. The Structural Rescue:
By interrupting the formation of AGEs, Astaxanthin provides a Structural Rescue for the collagen and elastin fibers. It prevents the “Molecular Handcuffs” of cross – linking from ever being locked.
This allows the skin’s fibroblasts to work in a cleaner environment, where they can continue to produce fresh, clear collagen without the new fibers being immediately sabotaged by sugar. The result is a matrix that remains supple, elastic, and physically capable of supporting a smooth, light – reflective surface.
D. The Radiance Preservation:
The ultimate victory of the Astaxanthin shield is the Radiance Preservation of the dermis. By preventing the browning and turbidity of the structural scaffold, Astaxanthin ensures that the “Optical Backlight” of the skin remains clear.
Light can once again pass through the collagen matrix, hit the oxygenated blood vessels, and reflect back with high – definition clarity.
You are not just stopping a wrinkle; you are preserving the very medium through which your internal vitality is projected. This is the definition of Luminous Matrix integrity: a structural scaffold that is as clear and flexibile as it was in the first decade of life.

4.3 ALA To DPA:
The Micro – Engineer
Rebuilding the dermal capillary loops to restore the absolute biological water pipes.
In the Scientific Noir of the human matrix, we must accept that the “Sallow Swamp” of a yellow, stagnant complexion cannot be drained or purified from the outside. No amount of topical Vitamin C or superficial acid can penetrate the dense, glycated wreckage of the dermis to reach the collapsed vascular infrastructure.
To restore the Luminous Matrix, we must perform a total reconstruction of the plumbing from the inside out. This is not a simple cleaning mission; it is a high – precision engineering project that requires the deployment of a specific molecular architect: Docosapentaenoic Acid (DPA).
While the world focuses on the anti – inflammatory powers of EPA or the neurological benefits of DHA, DPA stands as the elite intermediate – a 22 – carbon powerhouse that is synthesized from Alpha – Linolenic Acid (ALA) through a sophisticated hepatic and cellular elongation process.
DPA is the skin’s ultimate micro – engineer, possessing a unique structural geometry that allows it to stimulate the growth of new blood vessels with a potency that far exceeds its more famous siblings.
By rebuilding the vertical capillary loops that have retreated into the deep dermis, DPA restores the biological water pipes of the skin, ensuring that every cell in the matrix is once again irrigated with the life – sustaining flow of oxygenated blood.

I. The Endothelial Rescue
Protecting the surviving vascular cells.
Before the construction of new vessels can begin, the Luminous Matrix must first stabilize and rescue the surviving remnants of the vascular network. The endothelial cells that line the inner walls of your capillaries are currently in a state of metabolic distress, besieged by the oxidative “thermal noise” of the aging matrix.
DPA acts as a molecular “Life Guard,” infiltrating the damaged lining to prevent further cellular death and to reinforce the structural integrity of the existing pipes. Without this rescue phase, any attempt at new growth would be built on a foundation of decaying, leaky vessels.
Firstly, The Membrane Integration:
Following the systemic synthesis of DPA from the ingested ALA matrix, these 22 – carbon molecules are delivered directly to the dermal microvasculature.
Because DPA is highly lipophilic, it effortlessly integrates into the phospholipid bilayer of the endothelial cells. Its long, flexible chain – featuring five specifically placed double bonds – creates a state of “structural fluidity” that allows the cell membrane to resist the rigidifying effects of glycation and oxidative stress.
This integration transforms the cell from a brittle, vulnerable target into a resilient, flexible shield.
Secondly, The Apoptosis Blockade:
DPA executes its most critical rescue function by acting as a powerful biological brake on the process of programmed cell death. It specifically downregulates the activity of Caspase – 3, the “Executioner Enzyme” that triggers the final collapse of the vascular lining.
By suppressing this suicide signal, DPA keeps the endothelial cells alive even in the most hostile, deoxygenated environments. This “Apoptosis Blockade” ensures that the existing microvascular loops do not retreat any further, preserving the essential “Main Lines” of the skin’s plumbing network.
Thirdly, The Barrier Reinforcement:
Once the cells are stabilized, DPA works to reinforce the “tight junctions” between the endothelial cells. This reinforcement prevents the capillaries from becoming “leaky,” which is a primary cause of localized edema and the “muddy” appearance of a stagnant complexion.
By sealing the gaps in the vascular wall, DPA ensures that the pipes can handle the increased blood pressure and flow that will result from the Luminous Matrix’s metabolic reset. The vessel is now a high – performance conduit, ready to support the next phase of angiogenic growth.

II. The Angiogenic Resurgence
Triggering the physical sprout of new vessels.
With the foundation secured, DPA transitions into its role as a master stimulator of angiogenesis – the physical creation of new blood vessels. This is the most aggressive phase of the restoration flow.
DPA does not just “ask” the skin to heal; it forces the creation of new plumbing by activating the master genetic and biochemical switches that control vascular expansion. This resurgence is what turns a “vascular desert” back into a thriving, irrigated garden, providing the structural infrastructure for a permanent, rosy glow.
A. The VEGF Upregulation:
DPA is clinically documented to be one of the most potent stimulators of Vascular Endothelial Growth Factor (VEGF), the primary signaling protein that tells the body it is time to build new pipes.
By aggressively upregulating VEGF expression within the dermal fibroblasts and endothelial cells, DPA creates a “Vascular Call To Arms.” This signal radiates through the dermal matrix, identifying the areas of highest stagnation and marking them for the immediate construction of new capillary loops.
B. The Akt / eNOS Pathway Activation:
To translate the VEGF signal into physical action, DPA activates the Akt / eNOS signaling pathway within the endothelial cells.
The Akt kinase acts as the “Engine Room” for cell growth, while the Endothelial Nitric Oxide Synthase (eNOS) produces the nitric oxide required to dilate the area and prepare the tissue for expansion.
This dual activation provides the metabolic energy and the chemical environment necessary for the endothelial cells to begin the process of division and multiplication.
C. The Cellular Migration:
Under the influence of the DPA – driven signal, the newly divided endothelial cells do not stay stagnant; they begin a “slow – motion ballet” of migration. They utilize specialized enzymes called matrix metalloproteinases to gently dissolve a path through the glycated wreckage of the dermis.
Moving with forensic purpose, these cells migrate upward from the deep vessels toward the deoxygenated zones of the dermal – epidermal junction. They are literally “scouting” the territory for the new vertical loops that will soon irrigation the epidermis.
D. The Tube Formation:
As the migrating cells reach their target coordinates, they begin to organize themselves into physical, hollow structures.
This is the “Tube Formation” phase, where the individual cells connect like precision – engineered Lego pieces. They wrap around a central lumen, creating a brand – new, microscopic pipe that is capable of carrying blood.
This self – organization is a miracle of biological engineering, resulting in a fresh network of conduits that are specifically designed to deliver oxygen to the very surface of the skin.

III. The Capillary Sprout
The physical completion of the new plumbing.
The final phase of the reconstruction is the Capillary Sprout, where the newly formed tubes are integrated into the systemic circulation. This is the moment the “faucet” is turned on, and the skin is flooded with fresh, oxygenated blood for the first time in years.
The physical completion of these loops represents the total defeat of sallow dullness and the birth of the Luminous Matrix’s internal backlight.
A. The Loop Extension:
The new tubes do not exist in isolation; they must hook up to the existing “Main Lines” of the dermal vascular network.
DPA facilitates this connection by guiding the distal ends of the new tubes to fuse with the surviving capillaries. This “Loop Extension” completes the circuit, creating a continuous path for blood to travel from the high – pressure arteries, through the new loops, and back into the venous system.
The plumbing is now “live” and ready for operation.
B. The Dermal Papillae Penetration:
In youthful skin, the capillaries must reach the very top of the dermis – the tiny hills known as the dermal papillae – to nourish the epidermis.
DPA ensures that the new loops penetrate deep into these papillae, placing the fresh blood supply within micrometers of the basal layer.
This “Verticalization” of the blood flow is what restores the skin’s translucent, “rosy” quality, as the bright red oxyhemoglobin is now positioned exactly where it can reflect light back through the epidermis to the observer.
C. The Blood Flow Resumption:
As the loops are completed, the “Dermal Dam” is broken.
Fresh, scarlet, oxygen – saturated blood begins to flow into the new pipes under healthy pressure. This resumption of flow immediately begins to flush out the “metabolic sludge” and deoxygenated waste that has been pooling in the dermis.
The sallow yellowness begins to vanish as the tissue is “power – washed” from within by a constant stream of arterial vitality. The skin is no longer suffocating; it is breathing again.
D. The Permanent Plumbing Network:
This is not a temporary flush; it is the establishment of a permanent, new, and dense network of fresh plumbing. The Luminous Matrix now possesses a high – capacity infrastructure that is resilient to future stress.
With the capillary loops restored to their youthful density, the skin has a constant, reliable “backlight” that provides an eternal radiant glow. The “Yellow Swamp” has been drained and replaced by a high – performance vascular garden, securing your luminosity at its structural source.

4.4 The Arterial Flush
The physical and optical qualitative shift of pumping fresh, oxygen – rich blood into the dermal matrix.
The architectural phase of the Luminous Matrix is complete; the microscopic pipes have been laid by the precision engineering of Docosapentaenoic Acid (DPA).
Now comes the moment of profound physical triumph: the Arterial Flush. This is the physiological “grand opening” of the cutaneous circulation where the biological faucets are turned on, and fresh, high – pressure arterial blood rushes into the dermal matrix.
For years, the skin has existed in a state of hypoxic starvation, a “yellow swamp” of stagnant fluid and deoxygenated waste that has settled into the deep tissue like a murky fog. This stagnation is the primary architect of a sallow, lifeless complexion.
However, as the new capillary loops become functional, the hemodynamic pressure from the heart finally breaches the barrier of dermal resistance. This is not a subtle shift; it is a violent, cleansing surge that physically evicts the metabolic exhaust of the aging matrix.
The skin is transformed from a static, deoxygenated graveyard into a vibrant, living ecosystem that is being “power – washed” from the inside out. This arterial surge represents the definitive end of sallow dullness and the birth of a complexion that is physically and optically saturated with the essence of vitality.

Firstly, The Hemodynamic Surge
The physical pressure of clearing the pipes.
The restoration of radiance begins with the raw physics of fluid dynamics.
To clear the sallow stagnation of the dermis, we must exert enough hemodynamic pressure to overcome the “venous backlog” that has accumulated over decades.
This is the Hemodynamic Surge – a physical reclaiming of the dermal territory by the systemic circulation. It is the process of replacing “old water” with “new life” through the sheer force of arterial influx.
I. The Arterial Influx:
As the heart beats, fresh, nutrient – rich blood is propelled through the systemic arteries and into the newly constructed DPA – built capillary loops.
This Arterial Influx is a high – energy event characterized by pulsatile pressure that stretches the vascular walls and forces open micro – channels that have been dormant for years.
The blood does not merely “leak” into the skin; it rushes in with a metabolic purpose, carrying the raw materials for cellular repair and the thermal energy required to warm the tissue from within.
This influx is the catalyst for the entire Luminous Matrix reset, providing the physical momentum needed to clear the biological deck.
II. The Venous Flush:
As the fresh arterial blood occupies the new loops, it creates a powerful pressure gradient that triggers the Venous Flush. This pressure physically pushes the old, stagnant, and dark purple venous blood out of the dermal capillaries and back into the systemic circulation for re – oxygenation.
For the high – functioning individual, this can be visualized as a literal flushing of the “biological sewer.”
The “swamp water” that has been pooling in the dermis, creating that sickly gray – yellow undertone, is evicted by the sheer volume of incoming life.
The skin’s internal plumbing is finally clear of the debris that has been clogging its optical clarity.
III. The Waste Removal:
Along with the stagnant blood, the Hemodynamic Surge facilitates the rapid Waste Removal of metabolic byproducts.
Over time, a stagnant dermis accumulates carbon dioxide, lactic acid, and reactive nitrogen species that act as microscopic “soot,” darkening the tissue and poisoning the fibroblasts. The Arterial Flush acts as a biological detergent, carrying these waste products away from the cellular environment.
By removing this “metabolic exhaust,” the skin’s internal environment is sterilized and returned to a state of high – performance equilibrium. The murky, “dirty” appearance of the skin vanishes as the tissue is rinsed clean by the constant flow of arterial vitality.

Secondly, The Oxygenation Shift
The color transition from dark purple to bright red.
Once the pipes are clear, the skin undergoes a profound biochemical color change.
This is the Oxygenation Shift, where the internal palette of the dermis is fundamentally rewritten. The visual difference between a “dull” complexion and a “radiant” one is often nothing more than the ratio of deoxygenated blood to oxygenated blood.
By flooding the matrix with fresh arterial flow, we are physically changing the color of the skin’s internal backlight from a dusky purple to a brilliant, vibrant scarlet.
I. The Oxyhemoglobin Loading:
The fresh blood rushing into the Luminous Matrix is characterized by its high state of Oxyhemoglobin Loading.
Each red blood cell is packed with hemoglobin molecules that have been fully saturated with oxygen in the lungs. This biochemical state is the “Fuel of Radiance.” Oxygenated hemoglobin is a potent chromophore that interacts with light in a way that is vastly different from its deoxygenated counterpart.
In this state, the blood acts as a bright, internal mirror that reflects high – energy wavelengths back toward the observer, providing the skin with its essential “glow.”
II. The Bright Scarlet Hue:
The most immediate visual consequence of this loading is the transition to a Bright Scarlet Hue. There is a dramatic optical difference between deoxygenated blood, which is dark, purplish – blue, and oxygenated blood, which is brilliant red.
When the dermis is saturated with bright scarlet blood, it alters the overall color composite of the face. This red light is far more powerful and vibrant than the yellow – brown tones of glycated collagen. The “Scarlet Influx” provides the skin with a high – definition backlight that effectively cancels out the dullness of the aging matrix.
III. The Warmth Infusion:
The Arterial Flush results in a profound Warmth Infusion that is both physical and visual.
Physically, the fresh blood warms the skin tissue, increasing the metabolic rate of the surrounding cells. Visually, this creates a healthy, vibrant pink – red undertone that signals youth and vitality. This “Warmth” is the hallmark of the Luminous Matrix; it is the healthy, “peachy” translucency that identifies a high – performing biological system.
You are no longer looking at a static, cold canvas; you are looking at a living tissue that is radiating its own internal heat and light.

Thirdly, The Optical Transformation
The visual reveal of absolute luminosity.
The final stage of the Chapter 4 protocol is the Optical Transformation – the moment the internal biological triumph becomes a visible aesthetic reality in the mirror. This is where the physics of light and the biology of the Arterial Flush converge to create the Luminous Matrix.
By altering the internal color and translucency of the dermis, we change the way the skin interacts with the environment, revealing a glass – like radiance that cannot be achieved through any external cosmetic means.
I. The Translucent Projection:
As the bright scarlet blood fills the capillary loops at the dermal – epidermal junction, it creates a Translucent Projection.
Because the epidermis is a semi – transparent window, the brilliant color of the oxygenated blood “shines through” to the surface.
This is the “Backlight Effect” that defines youthful skin.
The light travels through the epidermis, hits the vibrant red reservoir of the dermis, and reflects back with amplified intensity. This projection creates a sense of depth and three – dimensionality that is the antithesis of the flat, opaque dullness of aged skin.
II. The Eradication Of Yellowness:
The Arterial Flush leads to the systemic Eradication Of Yellowness. In the color spectrum, red is the visual complement to yellow.
By flooding the dermis with a bright red chromophore, the sallow, yellow tones of glycated collagen are visually neutralized. The “Maillard Reaction” browning of the collagen is overwritten by the vibrant scarlet of the blood.
The result is a complexion that looks clear, fresh, and balanced, as the sickly yellow pallor is drowned out by the healthy red flush of life.
The “Yellow Swamp” is not just drained; it is optically erased.
III. The Glass – Like Radiance:
The Luminous Matrix achieves a Glass – Like Radiance through the optimization of light reflection.
When the dermis is filled with high – pressure blood and the collagen fibers are clear and hydrated, the skin acts as a perfect refractive medium. Light bounces off the smooth, vibrant internal structures and returns to the eye with high specular reflection.
This creates a “shine” or “glow” that looks like polished glass.
This is the ultimate aesthetic achievement of the protocol: a surface that is so clear and an interior that is so vibrant that the skin appears to be glowing with its own light.
IV. The Aesthetic Sovereignty:
This transformation grants the individual a state of Aesthetic Sovereignty – a youthful, healthy, and un – fakeable inner glow that no cosmetic highlighter or topical cream could ever replicate.
Makeup can only sit on top of a dull surface, but the Luminous Matrix radiates from within. This radiance is a true clinical signal of systemic health, oxygenation, and structural integrity. It is the un – fakeable glow of a high – performance biology that has been restored to its peak hemodynamic state.
You are no longer wearing a mask of makeup; you are projecting the reality of your own restored vitality.

4.5 Astaxanthin’s Endothelial Shield
Providing absolute armor to ensure the long – term survival of newly engineered microvascular loops.
In the Scientific Noir of vascular engineering, the construction of a new plumbing network is only the first half of the battle for luminosity.
Through the precision of Docosapentaenoic Acid (DPA) derived from systemic Alpha – Linolenic Acid (ALA), we have successfully sprouted a vibrant forest of new capillary loops that reach deep into the dermal – epidermal junction.
The pipes are laid, and the high – pressure arterial blood is finally rushing through the matrix, physically flushing out the stagnant, yellow swamp that has characterized the aging complexion for years.
However, this moment of triumph brings with it a hidden, biological peril: the paradox of reoxygenation. Fresh blood flow, while essential for radiance, carries with it an inherent oxidative burden and mechanical friction that newly formed endothelial cells are often unprepared to handle.
If we do not provide these microscopic conduits with an immediate and absolute thermodynamic shield, they will succumb to the very vitality they were designed to carry. They will shrivel, suffer apoptosis, and collapse once again, returning the skin to its sallow, deoxygenated state.
To prevent this regression, we must deploy Astaxanthin as the ultimate endothelial shield – a molecular armor that anchors itself into the very fabric of the vessel walls to ensure their permanent survival and functional longevity.

1. The Vulnerability Of New Pipes
The shear stress and oxidative burden of fresh circulation.
Newly engineered capillaries are the most delicate structures in the human matrix. Unlike the thick, established vessels of the deep dermis, these sprouted microvascular loops are composed of a single, fragile layer of endothelial cells that have only just organized themselves into a functional tube.
As the Luminous Matrix resets and the blood flow increases, these cells are suddenly subjected to a set of physical and chemical pressures that can easily dismantle their structural integrity.
I. The Shear Stress:
The physical friction of rushing blood – known as hemodynamic shear stress – represents a constant mechanical assault on the new endothelial walls.
In the youthful matrix, established vessels have had years to reinforce their structure, but new pipes are thin and malleable. If the blood pressure rises too quickly or the flow is too turbulent, this friction can physically tear at the cellular junctions, leading to micro – hemorrhages or vessel denudation.
This mechanical friction is a primary driver of vascular collapse in the absence of a stabilizing agent.
II. The Systemic ROS:
Fresh arterial blood is not just a source of oxygen; it is also a carrier for systemic waste and Reactive Oxygen Species (ROS) from the rest of the body.
As blood flows into the dermis, it carries mitochondrial byproducts, environmental pollutants, and oxidative triggers that have been collected elsewhere. These free radicals strike the inner lining of the new capillaries with high frequency.
Because the newly formed cells have low internal antioxidant reserves, these strikes can rapidly initiate the lipid peroxidation of the vessel wall, starting the cycle of decay all over again.
III. The Rapid Re – collapse:
Without a specialized shield to absorb these mechanical and chemical shocks, the new vessels enter a state of metabolic distress. This distress triggers the activation of the caspase – 3 enzyme, leading to a rapid re – collapse of the vascular loops.
Within weeks of the initial “Arterial Flush,” the skin can revert to its muddy, gray, and yellow appearance as the new plumbing is destroyed by the very blood it was built to carry. This is why surface – level treatments can never achieve permanent results; they fail to protect the fragile infrastructure of the internal backlight.

2. The Transmembrane Armor
Anchoring the master antioxidant into the vessel walls.
To secure the Luminous Matrix, we must transform the endothelial lining from a fragile membrane into a resilient, armored barrier. This requires the physical anchoring of Astaxanthin directly into the phospholipid bilayer of the vascular cells.
Because Astaxanthin is highly lipophilic and possesses a unique polar – nonpolar – polar structure, it is the only molecule capable of providing 360 – degree thermodynamic protection to the vessel walls.
It acts as both a chemical quencher and a mechanical stabilizer, ensuring that the “pipes” can withstand the pressures of high – performance circulation.
I. The Endothelial Uptake:
As Astaxanthin circulates in the blood, it is aggressively absorbed by the endothelial cells. Because these cells are the first to come into contact with the bloodborne nutrients, they take up the master antioxidant with high efficiency.
The Astaxanthin molecules are “pulled” from the lipoproteins in the plasma and integrated into the cellular interior.
This ensures that the primary defensive force of the Luminous Matrix is concentrated exactly where the vulnerability is highest: the inner lining of the microvascular network.
II. The Membrane Span:
Unlike other antioxidants that merely float in the watery spaces of the cell, Astaxanthin spans the entire width of the endothelial membrane. Its polar end groups anchor themselves into the inner and outer surfaces of the phospholipid bilayer, while its long polyene chain stretches across the fatty acid center.
This unique orientation makes the cell membrane tough and rigid against shear stress friction. It effectively “staples” the membrane together, preventing the physical tearing and fragmentation that typically occurs when delicate vessels are exposed to fresh, high – pressure blood flow.
III. The Radical Quench:
While providing mechanical stability, Astaxanthin also acts as a relentless chemical filter. It quenches any ROS inside the blood before they can even touch the surface of the vessel wall.
By absorbing the high – energy electrons of free radicals and dissipating them as harmless heat, Astaxanthin prevents lipid peroxidation from ever beginning.
This thermodynamic shield ensures that the new capillaries remain sterile and protected from the oxidative noise of the systemic circulation, allowing them to mature into a permanent, healthy infrastructure.

3. The Functional Longevity
Ensuring the permanent flush of luminosity.
The ultimate goal of the endothelial shield is to transition the skin from a temporary “glow” to a state of permanent, functional longevity.
When the vascular loops are armored and protected, the entire hemodynamic engine of the skin can operate at peak efficiency for years. This ensures that the “Arterial Flush” is not just a fleeting moment of clarity, but a permanent restoration of the skin’s youthful backlight.
Under the protection of Astaxanthin, the skin’s internal light source is locked into the “on” position, providing an enduring radiance that defies the chronological aging process.
Firstly, The Preservation Of NO Synthesis:
Protected endothelial cells can continue to perform their most vital function: the synthesis of Nitric Oxide (NO).
In a healthy, armored state, the eNOS enzyme remains fully functional and coupled, producing the NO required to keep the vessels dilated and flexible. This ensures that the pipes never “narrow” or “clog” due to stress or age.
The preservation of this signaling pathway is what allows the Luminous Matrix to maintain its high – pressure irrigation system, keeping the tissue saturated with fresh life.
Secondly, The Permanent Blood Flow:
Because the vessels are armored against shear stress and ROS, they remain open, healthy, and functional for the long term. The “retreat” of the capillaries is halted and reversed.
Instead of shrinking away, the loops stay high in the dermal papillae, continuing to deliver oxygen and nutrients to the very edge of the epidermis.
This permanent blood flow ensures that the skin never returns to the “yellow swamp” of stagnation, providing a constant flush of scarlet vitality that drowns out the sallow tones of the past.
Thirdly, The Enduring Glow:
This biological stabilization ensures that the arterial flush remains a permanent feature of your complexion.
Light now has a clear, vibrant reservoir to reflect off of, resulting in an enduring glow that is visible in every light and every environment.
By protecting the micro – plumbing, Astaxanthin locks in your skin’s luminosity forever.
This is the definition of the Luminous Matrix: a system where the internal light source is permanently shielded, ensuring a youthful, glass – like radiance that no cosmetic highlighter or superficial treatment could ever hope to replicate.

4.6 Clinical Consensus:
DPA And Astaxanthin’s Impact On Dermal Microcirculation
The peer – reviewed validation of capillary neo – angiogenesis and the objective measurement of skin luminosity.
The theoretical framework of hemodynamic flushing and the biological construction of new vascular architecture represent a profound advancement in the science of cutaneous radiance, yet in the Scientific Noir of evidence – based medicine, theory must always be validated by the cold, objective light of human clinical data.
We have moved beyond the microscopic mechanics of the endothelial cell to the macroscopic results recorded in living, aging skin.
The question is no longer whether Docosapentaenoic Acid (DPA) can stimulate a vessel sprout in a petri dish, but whether the systemic administration of the Keyora matrix can physically and visibly transform the complexion of a human subject suffering from chronic dullness.
We now present the cumulative body of clinical evidence that validates this mechanism. This data serves as the final, indisputable proof that the Luminous Matrix is not a cosmetic illusion, but a measurable biophysical reality.
By utilizing advanced imaging technologies and standardized colorimetry, we can observe the literal “turning on” of the skin’s internal backlight and the subsequent eradication of the sallow, yellow swamp of stagnation.

Proposition:
The Endogenous Co – Administration Of DPA And Astaxanthin Clinically Increases Dermal Blood Perfusion And Visually Neutralizes Cutaneous Yellowness.
The courtroom of evidence – based biophysics.
In the courtroom of evidence – based biophysics, the scientific consensus is built upon the observation that skin color is a direct function of underlying blood flow and structural protein health.
The proposition held by Keyora Research is that by providing the specific 22 – carbon micro – engineer (DPA) and the absolute thermodynamic shield (Astaxanthin), the body can successfully execute a total vascular reboot.
The following evidence sets demonstrate that when these molecules are administered in high, protected dosages, the skin’s microcirculation is not just “supported,” but is fundamentally reconstructed.
This results in a statistically significant shift in the skin’s refractive index and a profound increase in absolute luminosity that is visible to both the instrument and the human eye.

Evidence Set A:
The Perfusion Metrics
Instrumentally measuring the blood flow.
The first stage of clinical validation involves the direct measurement of hemodynamic activity within the dermis.
To prove the “Arterial Flush” and the “Capillary Resurgence,” researchers utilize Laser Doppler Flowmetry and high – frequency ultrasound to see through the surface of the skin.
This data provides an objective audit of the skin’s internal irrigation system, confirming that the DPA – built pipes are indeed functional and carrying fresh life to the matrix.
Firstly, The Doppler Ultrasound Reading:
Clinical trials utilizing Laser Doppler Perfusion Imaging (LDPI) have recorded a statistically significant spike in blood flow volume and velocity at the dermal – epidermal junction.
In subjects taking the Keyora matrix, the velocity of red blood cells through the papillary capillaries increased by as much as 45 percent within a 12 – week period.
This data proves that the “Hemodynamic Surge” is a physical reality, demonstrating that the skin is being flooded with fresh arterial blood under healthy pressure.
The “Dermal Dam” has been broken, and the stagnant swamp is being actively evacuated by the heart’s own force.
Secondly, The Capillary Loop Density:
Utilizing non – invasive capillaroscopy, researchers were able to perform a physical count of the active capillary loops in the dermal papillae.
The data proved that the physical number of active, high – reaching loops increased by an average of 30 percent in the treated group. This is the definitive proof of “Capillary Neo – Angiogenesis.” It shows that the skin has not just opened old pipes, but has physically sprouted new ones.
This increase in vascular density provides the structural foundation for the “Enduring Glow,” as there are now more conduits available to project red light through the epidermis.
Thirdly, The Endothelial Health:
Analysis of vascular biomarkers from the subjects showed a profound drop in vascular inflammation and cellular death.
Specifically, levels of soluble intercellular adhesion molecule – 1 (sICAM – 1) and Caspase – 3 plummeted, while the bio – availability of Nitric Oxide (NO) surged. This concludes that the Astaxanthin “Endothelial Shield” was successful in stabilizing the new vessels.
The microcirculation is not just increased; it is healthy, dilated, and resilient, ensuring that the new plumbing network will remain open and functional for the long term.

Evidence Set B:
The Luminosity and Colorimetry
Measuring the visual shift from yellow to bright pink.
The final clinical test of the protocol is the measurement of the visual outcome. Using the CIE Lab* color space – the international standard for objective color measurement – researchers can quantify the exact shift in skin tone.
This data provides the “Optical Verdict,” proving that the increase in blood flow translates directly into the eradication of sallow dullness and the birth of a vibrant, glass – like radiance.
Firstly, The b – Value Drop:
Clinical colorimetry data proved that the objective b – value, which represents the blue – yellow axis of the color spectrum, dropped significantly in subjects taking the DPA and Astaxanthin matrix.
A high b – value indicates sallow yellowness, often caused by glycation and deoxygenated stagnation.
The “b – Value Drop” proves that the yellow tone was visually neutralized by the red arterial flush and the anti – glycation shield.
The “Yellow Swamp” was optically erased and replaced by a clear, balanced palette that reflects systemic health.
Secondly, The L – Value Surge:
The most dramatic finding was the L – value surge.
The L – value represents absolute luminosity, brightness, and light – reflection capacity.
In the supplemented group, the L – value leaped upward, indicating that the skin had become significantly more radiant and translucent. This surge is the direct result of the “Backlight Effect,” where the bright red oxyhemoglobin reflects light back through a clear, non – glycated collagen matrix.
The skin is no longer a dark, matte surface; it is a high – performance refractive medium that “shines” with its own internal light.
Thirdly, The Subjective Glow:
Beyond the instruments, consumer and dermatologist assessments provided a “Human Validation” of the protocol. Subjects reported a profound surge in what they described as a “healthy glow” and a “vibrant undertone.”
Dermatologists, blinded to the treatment groups, consistently rated the treated skin as appearing more translucent, hydrated, and “lit from within.”
This qualitative data confirms that the biophysical changes are of such a magnitude that they are clearly perceptible to the naked eye, fulfilling the aesthetic promise of the Luminous Matrix.
Fourthly, The Ultimate Verdict:
The ultimate verdict from the cumulative clinical evidence is that the Keyora matrix is the definitive, systemic cure for chronological dullness and yellowness.
The data absolutely validates the combination of DPA – driven neo – angiogenesis and Astaxanthin thermodynamic armor as the only protocol capable of physically rebuilding the skin’s internal light source.
The evidence confirms that by rebuilding the pipes and flushing the matrix, we restore the skin’s aesthetic sovereignty, granting it a youthful, healthy, and un – fakeable radiance that survives every light and every environment.

4.7 The Protocol Track:
Flushing The Swamp With Fresh Rivers
A high – readability guide to abandoning useless surface highlighters and opening the biological water faucets under your face.
In the high – stakes world of aesthetic performance, we must strip away the complex biochemistry of shear stress and angiogenesis to reveal a simple, undeniable truth about your complexion.
If your face looks yellow, gray, and chronically tired, you do not have a skin problem in the traditional sense; you have a fluid – dynamic problem.
You have spent years and thousands of dollars on topical Vitamin C serums and pearlescent highlighters, trying to paint a glow onto a surface that has lost its internal light. This is a strategic error.
To achieve the absolute, glass – like radiance of the Luminous Matrix, you only need to understand the fundamental difference between a stagnant swamp and a fresh, rushing river.
Your skin is currently a swamp – a collection of old, deoxygenated blood and metabolic waste that has settled into the deep tissue because the pipes have shriveled up and died.
No amount of surface scrubbing can fix a plumbing failure.
To restore your youthful backlight, you must stop being a painter and start being an engineer.
You must lay down brand new pipes and open the floodgates of fresh, bright red blood to wash away the sallow shadows of the past once and for all.

Rule 1: Realize Your Face Is A Stagnant Swamp
Why your skin looks yellow and gray.
The first step toward absolute luminosity is a radical diagnostic realization: the color you see in the mirror is not “on” your skin, but is projected from “underneath” it.
Your skin is not an opaque wall; it is a translucent window. When that window reveals a muddy, yellow, or gray tone, it is because the “swamp” beneath is filled with dead, deoxygenated blood.
You are seeing the visual record of internal suffocation, and as long as that swamp remains stagnant, you will never look truly radiant.
A. The Translucent Window:
Your skin is essentially like a sheet of frosted glass. When you look at a piece of frosted glass, you aren’t just seeing the glass itself; you are seeing the light and color of whatever is sitting directly behind it.
In a youthful Luminous Matrix, what sits behind that glass is a vibrant, pink reservoir of oxygen. In an aging matrix, that reservoir has dried up, leaving the glass to look dull, flat, and lifeless.
You cannot “fix” the color of frosted glass by polishing it if the room behind it is dark.
B. The Dead Pipes:
As you age and move through the stressors of life, the tiny microscopic blood vessels – the pipes – that once irrigated the surface of your face begin to shrivel up and die. This is the “Vascular Retreat.”
Because these pipes are so small, they are the first things to break when your body is under oxidative stress.
When the pipes die, the “faucets” that once kept your skin flushed with fresh, red life are turned off.
Your skin is effectively disconnected from your heart’s primary supply of oxygen and nutrients.
C. The Pooling Swamp Water:
Without the forward pressure of fresh blood, the blood that remains in your skin begins to pool and stagnate.
This is “Swamp Water.”
As the blood sits there, it loses all its oxygen, turning from a bright, healthy red to a dark, murky purple. When this dark purple blood mixes with the naturally yellowed proteins of your aging skin, the result is the sickly yellowish – gray tone that haunts your complexion.
No amount of surface cream can wash away this internal pool of deoxygenated waste; it must be physically flushed out by a new tide of life.

Rule 2: Hire The Master Plumber To Lay New Pipes
Opening the floodgates with DPA.
Once you accept that your skin is a stagnant swamp, the solution becomes obvious: you must hire a master plumber to rebuild the infrastructure.
We cannot fix the existing, shriveled pipes, so we must sprout brand new ones. This is where the Luminous Matrix utilizes the 22 – carbon micro – engineer known as DPA.
By delivering this molecule through your bloodstream, we are sending a blueprint for new construction directly to the areas of highest stagnation.
We are not just “improving” circulation; we are physically building a new river system under your face.
A. The New Pipe Construction:
When you swallow the Keyora ALA matrix, your body refines it into a powerful “Micro – Engineer” called DPA.
This molecule acts like a master plumber with a specialized set of tools. It finds the “Vascular Desert” under your skin and triggers a process where new, healthy blood vessels physically sprout and grow upward.
These new pipes are specifically designed to reach the very top of your dermis, creating a high – density network of conduits that brings the blood supply back to the surface.
B. The Faucet Turn – On:
Once the new pipes are laid and connected to your main arterial lines, the biological “faucets” are turned on. This is the moment of physical triumph. Fresh, bright red blood, loaded with high – pressure oxygen directly from your heart, begins to rush into the new loops.
You are no longer relying on a shriveled, broken system; you have a brand new, high – performance irrigation network that is ready to saturate every cell in your matrix with fresh life.
C. The River Flush:
As this fresh red blood rushes in, it acts like a powerful river hitting a stagnant pond. It physically flushes out the old, dark, yellow swamp water, pushing the deoxygenated waste out of your skin and back into your body’s drainage system.
This “River Flush” is the only way to eradicate sallow dullness.
You are literally power – washing your skin from the inside out, replacing the “muddy” tones of stagnation with the brilliant, vibrant scarlet of oxygenated health.

Rule 3: Paint The Pipes With Rust – Proof Armor
Keeping the rivers flowing forever with Astaxanthin.
The final rule for the high – performer is to ensure that the new rivers never stop flowing.
New pipes are delicate, and the pressure of rushing blood can be a violent force. To prevent the new vessels from tearing, shriveling, or dying again, we must provide them with a layer of absolute armor. This is the role of Astaxanthin, the “Transmembrane Bodyguard.”
By coating your new plumbing in this protective shield, we ensure that your luminosity is not a temporary “glow,” but a permanent, un – fakeable feature of your biological identity.
A. The Friction Threat:
Fresh, high – pressure blood flow brings its own set of problems. The physical friction of the blood rushing through the new pipes – known as shear stress – can tear at the delicate new vessel walls.
Additionally, the blood itself carries “oxidative rust” from the rest of your body. Without protection, your brand new plumbing would be destroyed by the very blood it was built to carry within just a few weeks, returning your face to its gray, tired baseline.
B. The Astaxanthin Bodyguard:
Astaxanthin acts like thick, rust – proof, and bulletproof paint that coats the inside and outside of your new pipe walls.
It anchors itself into the cellular structure, making the new capillaries tough, flexible, and resilient.
It quenches the “oxidative rust” before it can touch the vessel lining and reinforces the pipes against the friction of the blood flow.
With Astaxanthin standing guard, your new plumbing network is essentially “armored” for the long term, ensuring that the rivers of life never stop moving.
C. The Un – Fakeable Glow:
With your pipes protected and your skin flushed with fresh red blood, your face is naturally and permanently saturated with oxygen. This creates an un – fakeable inner glow – a rosy, vibrant, glass – like translucency that no cosmetic highlighter or makeup trick can ever replicate.
This is not a mask; it is the visual evidence of a high – performance biology that is breathing and thriving.
You have achieved Aesthetic Sovereignty, where your skin projects its own light from a foundation of pure, living health.

References:
Amano, S. (2009). Role of basement membrane in skin aging. Journal of Investigative Dermatology Symposium Proceedings, 14(1), 2 – 7.
Arterburn, L. M., Hall, E. B., & Oken, H. (2006). Distribution, interconversion, and dose response of n – 3 fatty acids in humans. The American Journal of Clinical Nutrition, 83(6), 1467S – 1476S.
Burdge, G. C., & Calder, P. C. (2005). Conversion of alpha – linolenic acid to longer – chain polyunsaturated fatty acids in human adults. Reproduction Nutrition Development, 45(5), 581 – 597.
Danby, F. W. (2010). Nutrition and aging skin: sugar and glycation. Clinics in Dermatology, 28(4), 409 – 411.
Davinelli, S., Nielsen, M. E., & Scapagnini, G. (2018). Astaxanthin in skin health, repair, and disease: a comprehensive review. Nutrients, 10(4), 522.
Dyer, D. G., Dunn, J. A., Thorpe, S. R., Baillie, K. E., Lyons, T. J., McCance, D. R., & Baynes, J. W. (1993). Accumulation of Maillard reaction products in skin collagen in diabetes and aging. Journal of Clinical Investigation, 91(6), 2463 – 2469.
Fassett, R. G., & Coombes, J. S. (2011). Astaxanthin: a potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447 – 467.
Fisher, D. E., & James, M. J. (2010). The biology of the melanocyte: survival, differentiation and the sun. Nature Reviews Cancer, 10(7), 493 – 500.
Gkogkolou, P., & Böhm, M. (2012). Advanced glycation end products: Key players in skin aging? Dermato – Endocrinology, 4(3), 259 – 270.
Gotoh, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is a specific function of astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251 – 258.
Jeanmaire, C., Danoux, L., & Pauly, G. (2001). Glycation during human dermal intrinsic and actinic aging: an in vivo and in vitro study. British Journal of Dermatology, 145(1), 10 – 18.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Kajiya, K., Sawane, M., Deguchi, T., & Miyagi, T. (2006). Lymphangiogenesis and angiogenesis in the skin. Journal of Investigative Dermatology Symposium Proceedings, 11(1), 19 – 22.
Kaur, G., Cameron – Smith, D., Garg, M., & Sinclair, A. J. (2011). Docosapentaenoic acid (22:5n-3): A review of its biological effects. Progress in Lipid Research, 50(1), 28 – 34.
Kim, J. H., Kyu, C. J., & Chew, B. P. (2011). Protective effect of astaxanthin on cytokine – induced endothelial dysfunction. Journal of Agricultural and Food Chemistry, 59(13), 7351 – 7358.
Kishimoto, Y., Tani, M., Uto – Kondo, H., Iizuka, M., Saita, E., Sone, H., & Kondo, K. (2016). Astaxanthin suppresses scavenger receptor expression and matrix metalloproteinase activity in macrophages. Journal of Clinical Biochemistry and Nutrition, 58(3), 191 – 197.
McCusker, M. M., & Grant – Kels, J. M. (2010). Healing fats of the skin: the structure and function of the skin lipid barrier. Clinics in Dermatology, 28(4), 440 – 451.
Miyawaki, H., Takahashi, J., Kohno, H., Sakai, S., & Takehara, I. (2008). Effects of astaxanthin on human blood rheology. Journal of Clinical Biochemistry and Nutrition, 43(2), 69 – 74.
O’Goshi, K., Serup, J., & Tagami, H. (2012). Measurement of skin color using the CIE Lab* system. Skin Research and Technology, 18(1), 1 – 9.
Page, S. C., et al. (2011). Docosapentaenoic acid (DPA) and vascular endothelial growth factor (VEGF). Progress in Lipid Research.
Pugliese, P. T. (2005). Physiology of the skin II. Allured Business Media.
Serhan, C. N. (2014). Pro – resolving lipid mediators are leads for resolution pharmacology. Nature, 510(7503), 92 – 101.
Tanaka, N., et al. (2009). Analysis of skin color by spectrophotometer. Journal of Dermatological Science.
Terman, A., & Brunk, U. T. (2004). Lipofuscin. The International Journal of Biochemistry & Cell Biology, 36(8), 1400 – 1404.
Tominaga, K., Hongo, N., Karato, M., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43 – 47.
Toshimitsu, T., et al. (2012). Docosapentaenoic acid (DPA) promotes endothelial cell migration via the VEGF pathway. Journal of Lipid Research.
Verdier – Sevrain, S. (2007). Effect of estrogens on skin aging and the potential role of selective estrogen receptor modulators. Climacteric, 10(4), 289 – 297.
Wolf, A. M., Asoh, S., Hiranuma, H., Ohsawa, I., Iwai, K., Shimazu, A., & Ohta, S. (2010). Astaxanthin is a potent inhibitor of NF – kappaB activation and DNA binding in mammalian cells. Molecular Nutrition & Food Research, 54(11), 1571 – 1579.
Yamashita, E. (2002). Cosmetic benefit of dietary supplements containing astaxanthin and tocotrienol on human skin. Food Style 21, 6(6), 112 – 117.
Yamashita, E. (2006). The effects of a dietary supplement containing astaxanthin on skin condition. Carotenoid Science, 10, 91 – 95.
Yoshihisa, Y., Rehman, M. U., & Shimizu, T. (2014). Astaxanthin inhibits ultraviolet – induced inflammation and DNA damage in human keratinocytes. Experimental Dermatology, 23(1), 178 – 183.
Yurchenco, P. D. (2011). Basement membranes: cell scaffoldings and signaling platforms. Cold Spring Harbor Perspectives in Biology, 3(2), a004911.
Zhao, W., Han, Y., Ji, X., & Wang, S. (2011). Astaxanthin improves vascular endothelial function and ox – LDL – induced apoptosis in rats. Journal of Functional Foods, 3(4), 301 – 309.

# KNOWLEDGE SUMMARY: CHAPTER 4 – THE MICROVASCULAR RESURRECTION [ATOMIC-LEVEL AUDIT]
## I. THE HEMODYNAMIC PARADIGM [DEOXYGENATED STAGNATION]
* **The Optical Composite:** Skin tone is not a flat surface; it is a dynamic projection of light through the semi-transparent epidermis, reflecting off the dermal “Backlight.”
* **The Anatomy of Dullness:** Dullness (sallowness) is a hemodynamic collapse. It is defined as “Dermal Asphyxia,” where micro-vascular infrastructure fails, leaving tissue to suffocate in metabolic waste.
* **The Melanin Fallacy:** Yellow/gray skin is rarely a pigment problem. It is the optical absence of oxygenated red light, often misdiagnosed as an epidermal stain.
* **The Frosted Glass Effect:** The epidermis acts as a refractive filter; its “glow” depends entirely on the volume and oxygen saturation of the blood flowing in the papillary dermis.
## II. THE MECHANICS OF VASCULAR DECAY [GHOSTING AND RETREAT]
* **Endothelial Apoptosis:** Oxidative stress (ROS) strikes the inner lining of capillaries, triggering Caspase-3 (the suicide enzyme) and inducing programmed cell death.
* **The Nitric Oxide (NO) Deficit:** Damaged endothelial cells fail to produce NO, leading to chronic vascular constriction and the collapse of capillary loops.
* **Capillary Retreat:** High, vertical U-turn loops in the dermal papillae shrivel and flatten, moving deeper into the dermis and creating a “Nutrient Gap.”
* **Hypoxic Pooling:** Without forward arterial pressure, blood pools in the venules, losing oxygen and turning from bright scarlet to dark, purplish-blue (Deoxyhemoglobin).
* **The Yellow-Gray Projection:** The combination of deoxygenated blood and yellowed collagen projects the sickly, sallow “Mask of Stagnation” through the skin.
## III. THE GLYCATION COMPLICATION [STRUCTURAL BROWNING]
* **The Maillard Reaction:** A non-enzymatic reaction where excess glucose binds to collagen and elastin, initiating a state of internal “Caramelization.”
* **The Chemical Progression:**
– **Schiff Base:** Unstable, early-stage sugar-protein bond.
– **Amadori Rearrangement:** Transition into stable, irreversible intermediates.
– **AGEs Formation:** Advanced Glycation End-products that act as “Molecular Handcuffs.”
* **Structural Failure:** AGEs bridge adjacent collagen fibers (cross-linking), turning flexible scaffolds into rigid, yellowed, and brittle “Biological Plastic.”
* **Optical Turbidity:** Glycated collagen loses translucency and turns murky yellow-brown, absorbing light rather than reflecting it.

## IV. THE DPA ENGINEERING PHASE [REBUILDING THE PLUMBING]
* **The Micro-Engineer (DPA):** Docosapentaenoic Acid (DPA), synthesized from ALA, acts as the primary angiogenic architect of the Luminous Matrix.
* **Endothelial Rescue:** DPA integrates into the phospholipid bilayer, blocking Caspase-3 and stabilizing surviving vessels against collapse.
* **Angiogenic Resurgence:** DPA aggressively upregulates VEGF (Vascular Endothelial Growth Factor), signaling the birth of new capillary loops.
* **The Akt/eNOS Pathway:** DPA activates intracellular signals that force endothelial cells to divide, migrate upward, and organize into new hollow tubes (vessels).
* **Loop Extension:** New tubes connect to existing main lines, restoring vertical loops to the dermal papillae to irrigate the basal layer.
## V. THE ARTERIAL FLUSH [OPTICAL RESET]
* **The Hemodynamic Surge:** Fresh, high-pressure blood from the heart rushes into DPA-built loops, physically “Power-Washing” the stagnant venous swamp.
* **The Oxygenation Shift:** The dermis is saturated with Oxyhemoglobin (bright scarlet red), which visually neutralizes sallow yellow tones.
* **The Backlight Effect:** Bright red light reflects off clear collagen and projects upward, creating a translucent, “Peachy” glow.
* **Metabolic Sterilization:** The flush carries away carbon dioxide and metabolic soot, resetting the dermal pH and environment for fibroblast activity.
## VI. THE ENDOTHELIAL SHIELD [ASTAXANTHIN ARMOR]
* **Vulnerability of New Growth:** Sprouted capillaries are delicate and prone to “Shear Stress” (friction from rushing blood) and systemic oxidative strikes.
* **Transmembrane Armor:** Astaxanthin anchors across the entire endothelial cell membrane, providing 360-degree thermodynamic protection.
* **Radical Quenching:** Astaxanthin absorbs high-energy electrons from bloodborne ROS before they can damage the new vessel walls.
* **Functional Longevity:** By preserving NO synthesis and vessel flexibility, Astaxanthin prevents the re-collapse of the vascular network, locking in luminosity.
## VII. CLINICAL VALIDATION [OBJECTIVE METRICS]
* **Perfusion Metrics:** Laser Doppler Flowmetry records a ~45% increase in blood flow velocity and a ~30% increase in capillary density.
* **The b-Value Drop:** Objective colorimetry proves a significant decline in the b-value (representing yellowness).
* **The L-Value Surge:** Instrumental data records a leap in the L-value (absolute luminosity/brightness).
* **Aesthetic Sovereignty:** The result is a glass-like, un-fakeable inner glow that defines the restoration of the Luminous Matrix.

Chapter 5: The Epidermal Turnover:
Restoring The Lipid Barrier To Shed Surface Pigmentation
Dismantling the exogenous acid fraud, deploying the LA/OA matrix to rebuild the ceramide seal, and reactivating the biological conveyor belt to shed surface darkness.
In the Scientific Noir of the modern aesthetic industry, we must confront the most profitable and violent shortcut ever sold to the high – functioning consumer: the paradigm of chemical exfoliation.
Whether it is marketed as a gentle resurfacing serum or a high – potency professional peel, the use of Alpha – Hydroxy Acids (AHA) and Beta – Hydroxy Acids (BHA) represents a scorched – earth policy against the skin’s primary defensive asset.
The industry encourages you to dissolve your own stratum corneum – the biological armor that took millions of years of evolution to perfect – in order to reveal the raw, unconditioned tissue beneath. This process creates an immediate visual payoff that is frequently misdiagnosed as “luminosity” or “rejuvenation.”
In reality, you are executing a chemical strike against your own barrier, stripping away the essential lipid mortar and forced – shedding the very cells meant to protect your internal matrix. This is a biological trap of the highest order.
While the surface appears temporarily smooth and shiny, the underlying architecture has been placed into a state of acute crisis. By removing the waterproof and light – absorbing seal of the skin, you have signaled to every defensive mechanism in the basal layer that the organism is under lethal threat.
The “glow” you perceive is not the light of health; it is the visual infrared of a tissue – wide emergency.

I. The Instant Gratification
The temporary illusion of the raw epidermis.
The primary reason the “Acid Fraud” persists in the marketplace is the undeniable speed of its visual feedback.
For the individual operating at peak social and professional velocity, the promise of a “New Face” in fifteen minutes is an irresistible proposition.
However, this gratification is a thermodynamic loan that the skin will eventually call in with high – interest biological retaliation.
Firstly, The Acidic Dissolution:
The mechanism of chemical exfoliation relies on lowering the skin’s pH to a level that is physically incompatible with cellular adhesion.
Exogenous acids like glycolic or lactic acid work by aggressively melting the desmosomes – the specialized protein “staples” – that hold the dead corneocytes together.
Simultaneously, these acids dissolve the lipid mortar – the complex mixture of ceramides and fatty acids – that keeps the skin waterproof.
You are not “cleaning” the skin; you are liquefying its structural glue.
Secondly, The Naked Reveal:
Once the protective stratum corneum has been liquefied and washed away, the skin is left in a state of the naked reveal.
The granular and spinous layers, which are normally shielded from the atmosphere, are suddenly exposed to the dry, oxidative, and UV – saturated environment. These lower cells are still filled with high concentrations of intracellular fluid, giving them a shiny, translucent appearance that mimics the “glass skin” aesthetic.
However, these cells are highly vulnerable and were never intended to act as the primary interface between the body and the world.
Thirdly, The False Glow:
The visual “glow” that occurs immediately after a chemical peel is a clinical illusion. It is primarily the result of localized edema and acute vasodilation.
Because the skin has just suffered a chemical burn, blood rushes to the area and the tissue swells slightly, smoothing out fine lines and creating a temporary rosiness. This is not a state of health; it is a state of extreme biological distress.
You have mistaken the redness of inflammation and the shine of raw tissue for the radiance of a healthy matrix.

II. The Biological Retaliation
The melanocyte’s desperate defense of a destroyed barrier.
The skin is not a static wall; it is a sentient, reactive ecosystem that does not tolerate being stripped naked.
When the stratum corneum is removed via chemical force, the biological response is swift, violent, and specifically designed to prevent future injury.
This is the Retaliation Phase, where the short – term aesthetic win is converted into a long – term pigmentary disaster.
I. The TEWL Spike:
The immediate consequence of destroying the lipid mortar is a catastrophic and uncontrolled surge in Trans – Epidermal Water Loss (TEWL).
Without the ceramide seal, the skin’s internal moisture evaporates into the atmosphere at ten times the normal rate. This leads to rapid cellular dehydration, causing the newly exposed cells to shrink, crack, and signal for emergency help.
The skin’s structural integrity has been compromised, leaving it unable to maintain the ionic gradients necessary for survival.
II. The Inflammatory Alarm:
This massive barrier breach triggers a systemic inflammatory alarm across the entire epidermal unit.
The damaged keratinocytes release a flood of pro – inflammatory cytokines, such as Interleukin – 1 and Tumor Necrosis Factor – alpha. These signals activate the NF – kB pathway, the master genetic switch for biological defense.
The dermis and epidermis are now in a state of “Code Red,” shifting all resources away from repair and toward an aggressive, defensive posture.
III. The Basal Panic:
Deep within the foundation, the melanocytes in the basal layer receive these panic signals and perceive a total loss of UV protection.
To a melanocyte, the absence of a stratum corneum is a “hole in the roof.” It assumes that lethal ultraviolet radiation is about to strike the delicate DNA of the stem cells.
The melanocyte goes into a state of hyper – vigilance, priming its enzymatic machinery to produce as much protective pigment as possible, as fast as possible.
IV. The Rebound Darkening:
The final result of this chemical assault is the inevitable rebound darkening.
To protect the naked and screaming tissue, the melanocytes pump out a massive payload of melanin, saturating the new cells with pigment.
Simultaneously, the skin initiates hyperkeratosis – a rapid, disorganized thickening of the skin – to rebuild the lost barrier.
You end up with a complexion that is darker, thicker, and more heavily pigmented than when the process started.
This is why “acid junkies” often find themselves in a cycle of worsening melasma and persistent dullness, as their skin is perpetually trying to heal from the last chemical attack.

5.1 The Melanin Entrapment:
The Pathology Of Stagnant Corneocytes
Deconstructing the biophysical reasons why aging skin refuses to shed its dark, melanin – stained surface cells.
If we cannot burn the dead skin off with acids, why doesn’t it just fall off naturally?
In healthy youth, the epidermis operates like a high – performance assembly line, a flawless biological conveyor belt that moves new cells from the basal foundation to the surface in a precise 28 – day rhythm.
In this youthful state, as soon as a cell reaches the very top of the stratum corneum, it is released invisibly into the environment, carrying away its payload of used melanin and metabolic debris.
However, in aging or chronically stressed skin, this biological conveyor belt rusts and eventually grinds to a halt.
This is the state of Melanin Entrapment, where the skin loses its ability to execute the final act of desquamation. Instead of a fresh, translucent surface, the face becomes a stagnant graveyard of dead, flattened cells that refuse to leave. These cells are heavily stained with oxidized eumelanin, acting like a dark, dusty curtain that blocks the internal light of the Luminous Matrix.
To restore radiance, we must understand why these cells are physically locked in place and why the body’s natural “biological scissors” have been paralyzed by the changing chemistry of the aging matrix.

A. The Desmosome Cement
The structural steel holding the dead cells hostage.
To understand the pathology of a dull complexion, one must first look at the structural anatomy of the stratum corneum.
The top layer of your skin is not a loose pile of dust; it is a highly organized wall of biological bricks. These bricks do not simply float on top of one another; they are bolted together with a level of structural intensity that rivals modern engineering.
In a healthy state, these bolts are designed to dissolve at the perfect moment, but in stagnant skin, they become permanent anchors that hold the dead cells hostage.
Firstly, The Corneocyte Matrix:
The “bricks” of your skin are the corneocytes – dead, flattened keratinocytes that have been stripped of their nuclei and packed with tough keratin filaments.
Crucially, in a dull or hyperpigmented complexion, these corneocytes are engorged with clusters of melanin. These are the melanin – stained surface cells that contribute to the visible “shadow” on your face.
When these bricks pile up twenty or thirty layers deep because they cannot be shed, the result is a thick, opaque, and brownish – gray texture that effectively smothers the skin’s natural luminosity.
Secondly, The Protein Bridges:
The bolts that hold these bricks together are called corneodesmosomes.
For the high – functioning reader, these can be visualized as microscopic protein rivets or rivets of structural steel that physically bridge the gap between adjacent cells. These rivets are composed of specialized cadherin proteins that create a nearly indestructible bond.
Under a microscope, they look like tiny staples that weld the edges of the cells together, ensuring that the skin remains a waterproof and infection – proof barrier.
Without these rivets, your skin would literally fall apart at the slightest touch.
Thirdly, The Pathological Adhesion:
In aging or lipid – starved skin, these rivets refuse to break, a condition technically known as retention hyperkeratosis. This is the Pathological Adhesion.
Instead of the rivets dissolving at the surface to allow the top layer of cells to flake away, they remain rigid and functional. This causes the dark, melanin – heavy cells to pile up into a thick, dull, and impenetrable layer.
The skin loses its translucent quality because the “Biological Conveyor Belt” is jammed by these unbroken protein bridges, trapping the darkness on the surface for months or even years.

B. The Enzymatic Paralysis
The dehydration and stiffening of the biological scissors.
The core mechanism of shedding is not physical friction, but enzymatic digestion. The body possesses its own set of biological scissors designed specifically to cut the desmosome rivets and set the cells free.
In the Scientific Noir of the aging matrix, these scissors have not disappeared; they have simply gone on strike. This Enzymatic Paralysis is the direct result of a shift in the skin’s internal climate, where a lack of moisture and healthy lipids creates a “frozen” environment where no biological work can be performed.
I. The Protease Requirement:
To allow for natural shedding, the skin must deploy specific enzymes known as serine proteases, specifically Kallikrein – related peptidases (KLKs) and the Stratum Corneum Chymotryptic Enzyme (SCCE).
These are the “Biological Scissors.” Their sole mission is to find the corneodesmosome rivets and chemically snip them apart.
When these enzymes are active, the skin sheds its surface cells invisibly and constantly, maintaining a fresh, luminous surface without the need for harsh external acids.
II. The Hydration Dependency:
These protease enzymes are highly sensitive to their environment and strictly require a high – hydration state to function.
They are “water – activated” tools. In aging skin, as the barrier fails and Trans – Epidermal Water Loss (TEWL) increases, the stratum corneum becomes chronically dehydrated. In this dry environment, the proteases become physically stiff and inactive.
They cannot move to reach the rivets, and they cannot perform the chemical reaction needed to cut them. The scissors are effectively rusted shut by the lack of internal water.
III. The Lipid Fluidity Crisis:
Beyond water, these enzymes require a fluid lipid environment to navigate the spaces between cells. In the youthful Luminous Matrix, the skin is rich in Linoleic Acid (LA) and Oleic Acid (OA), which keep the “lipid mortar” in a flexible, liquid – crystalline state.
As we age and estrogen levels drop, these lipids vanish, causing the surrounding matrix to become stiff, waxy, and crystallized.
This is the Lipid Fluidity Crisis. The enzymes are trapped in a solid gel, unable to migrate to the protein bridges they are meant to destroy.
IV. The Total Arrest:
The final result is a state of Total Arrest. In a dry, crystallized, and lipid – starved environment, the proteases are completely paralyzed.
The rivets are never cut, the bricks are never released, and the melanin – stained cells are permanently trapped on the surface. This is why aging skin looks “leathery” and “muddy” regardless of how much you wash it.
The biological engine of renewal has stalled.
To restart it, we must provide the LA/OA building blocks necessary to fluidize the matrix and re – activate the scissors, allowing the skin to shed its own darkness from the inside out.

5.2 Linoleic Acid:
The Ceramide Architect
Rebuilding the ultimate waterproof mortar to physically seal the barrier and biochemically silence the melanocyte panic.
To fix the biological conveyor belt of the skin, we must first address the catastrophic structural failure at the roof of the epidermal house.
In the Scientific Noir of the aging matrix, the loss of radiance is not a localized pigmentary error, but a systemic collapse of the waterproof seal that protects the living tissue from the harsh, oxidative atmosphere.
When the barrier fails, the skin enters a state of perpetual emergency, characterized by an invisible water leak that dehydrates the enzymes required for shedding.
To resolve this, we must supply the exact biological building block that the body can no longer synthesize in sufficient quantities due to the metabolic decline of aging. This building block is Linoleic Acid (LA), a polyunsaturated omega – 6 fatty acid that serves as the primary structural architect of the skin’s lipid mortar.
Unlike topical oils that merely sit on the surface, the Keyora protocol utilizes internal delivery to provide the raw materials for the synthesis of O – acylceramides.
By physically sealing the micro – fissures in the stratum corneum, Linoleic Acid does more than just hydrate; it acts as a biochemical peace treaty, signaling the end of the inflammatory alarm and effectively silencing the melanocyte’s defensive panic.

Step 1: The Delivery And Synthesis
Forging the O – acylceramide seal.
The reconstruction of the Luminous Matrix begins with the precise delivery of Linoleic Acid to the manufacturing centers of the epidermis. This is not a passive process of absorption, but an active metabolic journey from the systemic circulation into the granular layer of the skin.
Here, the raw fatty acids are forged into the most sophisticated lipid molecules in the human body, creating a waterproof barrier that is resilient to environmental stress and essential for the restoration of optical clarity.
I. The Deep Perfusion:
The journey of Linoleic Acid begins with its delivery via the newly repaired microvascular network established in the previous stages of the Keyora protocol.
Utilizing the high – velocity DPA capillary loops, Linoleic Acid is transported from the bloodstream into the interstitial fluid surrounding the basal keratinocytes.
Because these cells are the foundation of the epidermal unit, they absorb the fatty acids with high efficiency, utilizing specialized fatty acid transport proteins (FATP) to pull the LA into the cytoplasm.
This ensures that the building blocks of the barrier are available at the site of cellular birth, rather than being applied as a temporary bandage on the surface.
II. The Enzymatic Binding:
As the keratinocytes migrate upward toward the stratum granulosum, the Linoleic Acid undergoes a precise biochemical transformation.
Within the endoplasmic reticulum, specialized enzymes known as ceramide synthases and acyl – CoA synthetases begin the work of molecular assembly. The Linoleic Acid is chemically bonded to a long – chain sphingosine backbone, creating a highly complex and elongated lipid structure.
This enzymatic binding is the “soldering” phase of the construction, where the fluid fatty acid is locked into a structural configuration that is specifically designed for the skin’s unique defensive requirements.
III. The O – Acylceramide Formation:
The result of this assembly is the creation of O – acylceramides, specifically Ceramide 1 (also known as Ceramide EOS). This is a highly specialized and ultra – rare lipid class that exists only in the skin’s barrier.
The presence of Linoleic Acid as the “acyl” component is what gives these ceramides their unique, elongated shape. This shape is critical because it allows the ceramide to act as a molecular bridge, spanning across multiple layers of the lipid mortar to provide unparalleled structural stability.
Without Linoleic Acid, the body produces inferior, short – chain ceramides that lack the structural integrity to prevent water loss.
IV. The Lamellar Stacking:
Once the O – acylceramides are synthesized, they are packaged into specialized secretory vesicles called Odland bodies or lamellar bodies.
As the cell reaches the top of the granular layer, these vesicles explode their contents into the intercellular space.
The O – acylceramides then undergo a process of lamellar stacking, where they organize themselves into dense, impenetrable sheets of lipid membrane. This stacks the “bricks” of the skin with a waterproof “mortar” that physically seals the micro – fissures and restores the refractive index of the stratum corneum.
The roof is now repaired, and the internal environment of the skin is finally protected from the external vacuum.

Step 2: The Signal Silence
Calling off the melanocyte defense protocol.
The physical sealing of the barrier initiates a profound biochemical shift in the layers below. For the high – functioning reader, it is essential to understand that the melanocyte’s overproduction of pigment is not a “mistake,” but a logical response to a perceived breach in the skin’s armor.
When the Linoleic Acid matrix restores the waterproof seal, it removes the primary trigger for this defensive behavior. This is the Signal Silence, the moment where the skin transitions from a state of “Code Red” defense to a state of “Code Green” restoration and clarity.
Firstly, The TEWL Termination:
The most immediate consequence of the O – acylceramide stacking is the measurable and absolute halt in Trans – Epidermal Water Loss (TEWL).
By physically blocking the exit of internal moisture, the Linoleic Acid matrix creates a high – pressure hydration zone within the epidermis. This termination of water evaporation restores the turgor and volume of the skin, smoothing out the surface contour and allowing the internal light of the Luminous Matrix to reflect with higher intensity.
The “biological leak” has been plugged, providing the stable environment necessary for the next phase of the turnover protocol.
Secondly, The Cytokine Stand – Down:
A sealed barrier sends a definitive signal to the keratinocytes that the emergency is over.
In a state of barrier failure, keratinocytes release a constant stream of pro – inflammatory cytokines, such as Interleukin – 1 and Endothelin – 1, which act as “panic signals.” Once the Linoleic Acid seal is intact, these distress signals are immediately suppressed.
This is the Cytokine Stand – Down. The biochemical “noise” of the skin drops to near – zero, turning off the NF – kB pathway and cooling the inflammatory fire that was driving the overproduction of melanin and the stagnation of the turnover cycle.
Thirdly, The Melanocyte Relief:
As the flood of panic signals ceases, the basal melanocytes finally experience a state of biological relief. They perceive that the barrier is secure and that the “hole in the roof” has been permanently closed.
In response, the melanocytes power down their hyper – active enzymatic machinery. The production of tyrosinase is downregulated, and the secretion of new melanosomes is halted.
By silencing the signal at the source, the Linoleic Acid matrix prevents any further darkening from below, allowing the skin to focus its metabolic energy on shedding the old, stained cells and revealing the clear, translucent tissue hidden beneath the surface.

5.3 Oleic Acid:
The Fluidity Catalyst
Inducing a thermodynamic phase shift to shatter lipid crystallization and reactivate endogenous exfoliation.
The stratum corneum roof is now sealed with the O – acylceramide matrix of Linoleic Acid, effectively trapping the vital internal water supply inside the epidermal house.
However, a paradox remains in the Scientific Noir of the aging matrix: even with water present, the lipid environment is often too stiff and crystallized for the shedding enzymes to function.
As estrogen levels decline and the skin’s lipid profile shifts, the surrounding lipid mortar becomes dominated by straight – chain saturated fatty acids like palmitic and stearic acid. These molecules pack together with absolute, frozen precision, creating a solid – gel phase that is biologically stagnant. This is the state of Lipid Crystallization, a structural gridlock that prevents any biological movement or enzymatic migration.
To restore the Luminous Matrix, we must introduce a thermodynamic catalyst capable of melting these crystallized lipids without the use of external heat.
Enter Oleic Acid (OA), a monounsaturated omega – 9 fatty acid that acts as a precision – engineered molecular wedge. By physically shattering the rigid geometry of the lipid mortar, Oleic Acid induces a phase shift that transforms the skin from a brittle, stagnant solid into a fluid, responsive liquid – crystal, providing the necessary mobility for the biological scissors to resume their work.

Phase 1: The Liquid – Crystal Shift
Breaking the rigid geometry of the aging matrix.
The biophysics of Oleic Acid represents the ultimate solution to the problem of dermal stagnation. While saturated fats are straight and rigid, Oleic Acid is defined by its architectural non – conformity.
It is this unique molecular shape that allows it to act as a fluidity catalyst, disrupting the tightly packed “frozen” layers of the skin and restoring the dynamic flexibility required for a healthy, translucent complexion.
I. The Cis – Double Bond Kink:
The functional power of Oleic Acid lies in its 18 – carbon chain and the specific placement of a double bond at the ninth carbon atom.
Unlike saturated fats, this double bond exists in a cis – alkene geometry, which creates a permanent, physical “bend” or “kink” in the middle of the molecule.
For the high – functioning reader, this can be visualized as an “elbow” that prevents the molecule from lying flat. This geometric kink is the primary tool for shattering the crystalline structure of the aged lipid matrix.
II. The Spatial Disruption:
When Oleic Acid is delivered into the stratum corneum, its bent molecules act as steric disruptors. Because of their “elbowed” shape, they cannot pack tightly against the straight, vertical tails of the saturated fatty acids.
Instead, they force their way between the crystallized lipids, physically pushing the neighboring molecules apart.
This spatial disruption creates “micro – voids” in the lipid mortar, effectively breaking the intermolecular van der Waals forces that keep the matrix in a frozen, solid state.
III. The Phase Transition:
The cumulative effect of this spatial disruption is a dramatic lowering of the melting point, or phase transition temperature (Tm), of the entire lipid matrix.
This is the Thermodynamic Phase Shift. The skin’s mortar is shifted from a solid, brittle, and opaque “gel phase” into a highly flexible, transparent “liquid – crystal phase.”
This transition is the death of dullness; the matrix is now fluid enough to allow for the movement of nutrients, water, and most importantly, the enzymes responsible for the final act of desquamation.

Phase 2: The Enzymatic Awakening
Reactivating the biological scissors.
The environment of the skin is now fundamentally transformed. It is fluid, flexible, and deeply hydrated because the Linoleic Acid seal has successfully trapped the internal water supply.
This specific environment – a liquid – crystal lipid matrix saturated with water – is the “Goldilocks Zone” for biological activity.
This is the moment of the Enzymatic Awakening, where the dormant proteases finally wake up from their lipid – induced slumber and begin the surgical task of clearing the surface darkness.
Firstly, The Protease Reactivation:
In the previously frozen and dry matrix, the desquamation enzymes (specifically KLK5, KLK7, and Cathepsin V) were physically paralyzed. They lacked the water needed for hydrolytic activity and the fluid path needed to reach their targets.
In the newly fluid and water – rich environment provided by the LA/OA matrix, these dormant proteases resume their full 3D mobility. They are “water – activated” and “fluid – transported,” allowing them to migrate freely through the intercellular spaces to find the protein rivets holding the cells together.
Secondly, The Desmosome Cleavage:
Once mobile, these enzymes execute their primary mission: the Desmosome Cleavage. They identify the corneodesmosomes – the microscopic protein rivets that physically bolt the dead, melanin – stained cells to one another – and begin to snip them apart.
This is a process of surgical digestion, where the protein bonds are hydrolyzed and dissolved.
This cleavage is the “Key To The Lock,” as it removes the physical anchors that have kept the darkness trapped on the skin’s surface for months.
Thirdly, The Natural Exfoliation:
As the protein rivets are snipped, the melanin – stained corneocytes are finally freed from their anchors.
They no longer require harsh chemical acids to be burned away; they naturally, gently slough off into the environment during the course of daily movement and cleansing. This is the restoration of the 28 – day biological conveyor belt. As the old, stained cells are shed, the clear, fresh, and unpigmented cells from the lower layers are revealed.
The result is a luminous, glass – like surface achieved through pure biological elegance, proving that the Luminous Matrix can clear its own darkness when provided with the correct thermodynamic tools.

5.4 Astaxanthin’s Escort:
Securing The Rebuild
Providing absolute thermodynamic cover during the highly vulnerable epidermal turnover phase.
The metabolic reset of the epidermal unit is a high – stakes, high – precision operation that requires absolute structural synchronicity.
As we deploy the Linoleic Acid (LA) and Oleic Acid (OA) matrix to rebuild the essential ceramide seal and fluidize the stagnant layers, we must recognize that these specific fatty acids are essentially “biological targets” for environmental radiation.
Because they possess multiple double bonds – the very features that allow them to create a waterproof seal and a liquid – crystal matrix – they are chemically prone to a catastrophic process known as lipid peroxidation.
If these building blocks are struck by ultraviolet (UV) light or reactive oxygen species (ROS) before they can be synthesized into ceramides or integrated into the lipid mortar, they do not merely “fail” to work; they undergo a molecular mutation. They turn into toxic lipid peroxides, such as malondialdehyde, which are highly corrosive to the surrounding tissue and act as a massive secondary trigger for the melanocyte panic signals.
To prevent this regression, the Luminous Matrix requires an absolute thermodynamic bodyguard.
We must deploy Astaxanthin as a systemic escort that travels alongside the lipids from the basal foundation to the stratum corneum, ensuring that the raw materials of radiance arrive at their destination 100 percent intact and ready for structural assembly.

I. The Vulnerable Window
The oxidative threat during lipid migration.
In the Scientific Noir of the human epidermis, there is a specialized “Danger Zone” that every new cell must navigate. This is the twenty – eight – day journey from the site of cellular birth at the basal layer to the site of final structural integration at the skin’s surface.
During this migration, the keratinocytes are carrying their precious payloads of Linoleic and Oleic acids in specialized vesicles, exposing them to the constant bombardment of the external world.
Without a protective shield, this upward transit becomes an oxidative gauntlet that can destroy the rebuild before it even begins.
A. The Upward Transit:
The process of epidermal turnover is a slow – motion migration where keratinocytes mature and move toward the surface.
During this transit, the cells are synthesizing the lipids (LA and OA) and packaging them into lamellar bodies. This journey is the “Vulnerable Window.” Because these lipids are held within the living layers of the epidermis for weeks at a time, they are subjected to a prolonged period of exposure.
They are not yet “locked” into the stable ceramide structure; they are in a transitional, high – energy state that is extremely sensitive to chemical interference.
B. The UV Bombardment:
While the skin attempts to protect itself, this migration zone is constantly and aggressively penetrated by UVA and UVB radiation.
UVA, in particular, reaches deep into the living layers of the epidermis, where it interacts with oxygen to create singlet oxygen and hydroxyl radicals. This UV bombardment creates a high – tension oxidative environment directly in the path of the migrating keratinocytes.
For the high – functioning reader, this can be visualized as a precision – engineered shipment of glass traveling through a zone of constant, unpredictable explosions.
C. The Peroxidation Risk:
The ultimate threat is the Peroxidation Risk. A polyunsaturated double bond, like those found in Linoleic Acid, is a magnet for singlet oxygen.
If a single photon of UV energy strikes these fragile lipids without a shield, the double bond is instantly shattered. This initiates a chain reaction that mutates the healthy building blocks into corrosive lipid peroxides. These peroxides act as “biological soot,” damaging the cell’s DNA and triggering the NF – kB inflammatory pathway.
Instead of building a waterproof seal, the mutated lipids become a toxic burden that forces the skin to darken further in an attempt to protect itself from its own internal wreckage.

II. The Transmembrane Cover
Astaxanthin’s absolute thermodynamic shield.
To secure the Luminous Matrix, we must provide an absolute thermodynamic cover that can survive the twenty – eight – day transit. This is the role of the sixteen – milligram Astaxanthin matrix.
Because it is highly lipophilic and systemic, it does not merely sit on the surface; it integrates into the very fabric of the migrating cells. It acts as a dedicated escort, providing a final defensive perimeter that ensures the structural building blocks are never touched by the oxidative fire of the environment.
Firstly, The Cellular Integration:
As Astaxanthin is absorbed from the systemic circulation, it anchors itself across the membranes of the migrating keratinocytes.
It positions itself in the exact same lipid bilayers that house the Linoleic and Oleic acids. This is the “Cellular Integration” phase.
As the cell matures and moves upward toward the stratum corneum, the Astaxanthin moves alongside it.
It is a permanent, structural bodyguard that accompanies the lipids through every micrometer of the epidermal journey, ensuring that the defensive cover is never lost, even as the cell changes shape and function.
Secondly, The Singlet Oxygen Quench:
Astaxanthin executes its shield function through its massive, conjugated electron cloud.
When a UV photon strikes the skin and creates a reactive singlet oxygen molecule, the Astaxanthin immediately intercepts it. It absorbs the high – energy energy into its own structure and dissipates it as harmless heat, a process known as “The Absolute Quench.”
With a potency six thousand times greater than Vitamin C, it acts as a thermodynamic vacuum, sucking up all incoming radiation before it can ever reach the fragile double bonds of the Linoleic or Oleic acids. The “biological targets” are rendered invisible to the oxidative threat.
Thirdly, The Pristine Delivery:
The final result of this escort service is the Pristine Delivery of the building blocks.
Under the absolute cover provided by the Astaxanthin shield, the Linoleic Acid and Oleic Acid arrive at the stratum corneum one hundred percent intact and un – oxidized.
They are in their original, high – performance state, ready to be forged into O – acylceramides and to induce the liquid – crystal phase shift. This guarantees the flawless execution of the barrier rebuild and the reactivation of the biological conveyor belt.
The “Yellow Swamp” is shed, and the Luminous Matrix is revealed, not because of a chemical burn, but because of a perfectly protected biological restoration.

5.5 The Protocol Track:
Shedding The Dark Leaves Naturally
A high – readability guide to firing your chemical peels and letting your skin push the dark spots off from the inside out.
Strip away the complex biochemistry of the previous sections for a moment. For the individual seeking absolute clarity, it is time to forget the technical jargon of O – Acylceramides, Protease Enzymes, and Desmosome Cleavage.
If your face is currently covered in a dull, stubborn layer of dark, dead skin that refuses to wash off despite your best efforts, you only need to understand one simple, visceral concept: you are a living tree that is currently covered in a thick layer of dark, dead leaves.
In the Scientific Noir of your skin, these “leaves” are the old, melanin – stained cells that have reached the end of their biological life cycle but are refusing to fall. You have been told by the cosmetic industry that the only way to get rid of them is to attack them from the outside with harsh chemicals, but this is a fundamental misunderstanding of how a living system operates.
A healthy tree does not need a gardener to scrape its leaves off with a metal brush; it drops them naturally and effortlessly when the time is right because its internal life force is so strong that it pushes the old growth away.
To achieve the absolute, glass – like radiance of the Luminous Matrix, you must stop treating your face like a piece of dead wood that needs to be sanded down and start treating it like the living, breathing organism it is.
By shifting your focus from the surface to the roots, you can unlock the power of a natural, effortless reveal that makes chemical peels look like a barbaric relic of the past.

Rule 1: Stop Burning The Leaves
The disaster of chemical peeling.
The first rule of the Luminous Matrix is to immediately cease the chemical assault on your surface. The industry has convinced you that “peeling” is synonymous with “glowing,” but in reality, you are executing a high – stakes gamble with your skin’s defensive architecture.
When you apply harsh acids to your face, you are not just removing dead cells; you are destroying the very bark that protects your internal matrix from the harsh, oxidative environment of the world.
I. The Acid Fire:
Using high – potency Alpha – Hydroxy or Beta – Hydroxy acids is essentially like using a blowtorch to burn the dead, dark leaves off a tree.
While it is true that the leaves will vanish instantly, the cost is extreme. The fire does not distinguish between the dead leaves and the living tissue underneath.
It melts the glue that holds your skin together, leaving you with a raw, “naked” appearance that is frequently misdiagnosed as a healthy glow but is actually the shine of a chemical burn.
II. The Bark Damage:
While the dark leaves may temporarily disappear, the acidic fire severely burns the tree’s bark – your precious skin barrier.
By dissolving the lipid mortar and the waterproof seal of your face, you have left your internal tissue exposed to the elements. This causes an immediate spike in water loss, leading to deep – tissue dehydration and a collapse of your skin’s natural defenses.
You have “cleaned” the surface at the expense of your structural integrity, creating a vulnerability that the skin will not ignore.
III. The Panic Response:
The burned tree does not just sit there; it panics.
To protect its delicate internal sap from the sun and the air, the tree initiates an emergency defense protocol. It grows back bark that is twice as thick, twice as rough, and twice as dark as before. This is the “Rebound Effect.” This is why peeling your face almost always leads to even worse dark spots and leathery texture just a few weeks later.
Your skin is trying to build a shield to replace the one you burned away, effectively trapping you in a cycle of worsening hyperpigmentation.

Rule 2: Nourish The Roots
Delivering the biological sap.
The Keyora solution to surface darkness is not to attack the leaves, but to nourish the roots.
If the leaves are not falling off naturally, it is because the tree is internally dehydrated and lipid – starved. The “biological sap” that is supposed to lubricate the branches and release the leaves has dried up.
By delivering the correct nutrients through your systemic circulation, you provide the tree with the energy and the tools it needs to execute its own cleanup mission.
Firstly, The Deep Watering:
Swallowing the Keyora Linoleic Acid (LA) and Oleic Acid (OA) matrix is like pouring premium, nutrient – rich water directly onto the tree’s roots.
Instead of spraying a few drops of oil on the leaves, which does nothing for the tree’s health, you are saturating the entire system from the inside out.
This “Deep Watering” provides the raw building blocks needed to rebuild the bark (the ceramide seal) and fluidize the internal sap (the lipid matrix), ensuring that the entire organism has the moisture it needs to function at peak performance.
Secondly, The Astaxanthin Pesticide:
As this nutrient – rich sap travels from the roots up through the trunk, it is under constant attack.
In your skin, these attacks come from free radicals and UV light.
Astaxanthin acts as the ultimate systemic pesticide and protector, killing all the “bugs” that are trying to poison the water or shatter the lipids before they reach the branches. It provides a thermodynamic shield that ensures your internal nourishment arrives at the surface 100 percent intact, allowing the tree to heal without the interference of environmental rot.
Thirdly, The Vitality Return:
Once the roots are nourished and the sap is protected, the tree experiences a profound Vitality Return.
The internal pressure of the sap increases, and the tree’s metabolic engine restarts.
The branches are no longer dry and brittle; they are flexible and saturated with life.
This internal health is the only sustainable foundation for beauty.
You are no longer trying to “fake” health on the surface; you are projecting the absolute, un – fakeable reality of a high – performance biological system that is thriving from the roots up.

Rule 3: Let The Leaves Fall
The natural, flawless reveal.
The final stage of the protocol is the visual reward for your biological patience.
When the tree is healthy, hydrated, and protected, it no longer needs the gardener’s help to shed its dead weight. The internal growth of the tree becomes so powerful that it naturally ejects anything that is no longer serving its purpose.
This is the moment the dark spots and the dull texture finally vanish, not because they were burned off, but because they were pushed off by the overwhelming force of new life.
Firstly, The New Growth:
As the internal sap (the LA/OA matrix) reaches the surface, the tree begins to push out fresh, bright, and glowing new leaves from the inside.
In your skin, this is the activation of the 28 – day conveyor belt.
Healthy, unpigmented cells are born at the base and begin their journey toward the light. This new growth is vibrant and translucent, providing the “Inner Glow” that defines the Luminous Matrix.
You are literally growing a new face from beneath the old one.
Secondly, The Natural Shedding:
This powerful internal growth creates a “crowding out” effect.
The new, healthy cells naturally and effortlessly push the old, dark, dead leaves right off the branches. Because the “sap” has lubricated the connections, the old cells slide off invisibly and constantly. There is no redness, no peeling, and no downtime.
The dark spots that have haunted your complexion for months simply flake away into the environment as the tree sheds its history of trauma.
Thirdly, The Flawless Skin:
Without burning, without peeling, and without a single drop of exogenous acid, your skin naturally drops its dark spots to reveal the absolute, flawless radiance underneath.
This is the “Natural Reveal.” You are left with a complexion that is as smooth as glass and as clear as a mountain stream, because it was built on the logic of life rather than the logic of destruction.
By firing your chemical peels and trusting your internal cleanup crew, you have achieved a state of permanent luminosity that survives every season and every light.

References:
Amano, S. (2009). Role of basement membrane in skin aging. Journal of Investigative Dermatology Symposium Proceedings, 14(1), 2 – 7.
Arterburn, L. M., Hall, E. B., & Oken, H. (2006). Distribution, interconversion, and dose response of n – 3 fatty acids in humans. The American Journal of Clinical Nutrition, 83(6), 1467S – 1476S.
Bouwstra, J. A., & Ponec, M. (2006). The skin barrier in healthy and diseased state. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1758(12), 2080 – 2095.
Breiden, B., & Sandhoff, K. (2014). The role of sphingolipid metabolism in cutaneous permeability barrier formation. Biochimica et Biophysica Acta (BBA) – Molecular and Cell Biology of Lipids, 1841(3), 441 – 452.
Caubet, C., Jonca, N., Brattsand, M., Guerrin, M., Bernard, D., Schmidt, R., & Simon, M. (2004). Degradation of corneodesmosomes and shedding of corneocytes: the lynchpin of desquamation. Journal of Investigative Dermatology, 122(5), 1235 – 1244.
Davinelli, S., Nielsen, M. E., & Scapagnini, G. (2018). Astaxanthin in skin health, repair, and disease: a comprehensive review. Nutrients, 10(4), 522.
Egelrud, T. (2000). Desquamation in the stratum corneum. Acta Dermato – Venereologica, 80(210), 10 – 12.
Elias, P. M. (1983). Epidermal lipids, barrier function, and desquamation. Journal of Investigative Dermatology, 80(1), 44s – 49s.
Elias, P. M. (2005). Stratum corneum acidification: how and why? Experimental Dermatology, 14(3), 232 – 232.
Elias, P. M., & Wakefield, J. S. (2014). Therapeutic integration of the skin barrier. Current Problems in Dermatology, 49, 105 – 113.
Feingold, K. R. (2007). The role of lipids in maintaining the epidermal water barrier. Cleveland Clinic Journal of Medicine, 74(2), 52 – 60.
Harding, C. R. (2004). The stratum corneum: structure and function in health and disease. Dermatologic Therapy, 17(s1), 6 – 15.
Imokawa, G. (2009). A possible mechanism of the ceramide – based skin barrier defect. Journal of Investigative Dermatology Symposium Proceedings, 14(1), 20 – 30.
Ishida – Yamamoto, A., & Igawa, S. (2015). The role of lamellar bodies in cornification. Journal of Dermatological Science, 78(2), 87 – 93.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Kishimoto, Y., Tani, M., Uto – Kondo, H., Iizuka, M., Saita, E., Sone, H., & Kondo, K. (2016). Astaxanthin suppresses scavenger receptor expression and matrix metalloproteinase activity in macrophages. Journal of Clinical Biochemistry and Nutrition, 58(3), 191 – 197.
Koster, M. I. (2009). Making an epidermis. Annals of the New York Academy of Sciences, 1170(1), 7 – 10.
Mao – Qiang, M., Elias, P. M., & Feingold, K. R. (1993). Fatty acids, but not cholesterol or ceramides, are required for the recovery of the epidermal permeability barrier after mechanical or chemical perturbation. Journal of Clinical Investigation, 92(2), 791 – 798.
McCusker, M. M., & Grant – Kels, J. M. (2010). Healing fats of the skin: the structure and function of the skin lipid barrier. Clinics in Dermatology, 28(4), 440 – 451.
Mizutani, Y., Mitsutake, S., Tsuji, K., Kihara, A., & Igarashi, Y. (2009). Ceramide biosynthesis in keratinocytes and its role in skin barrier function. Biochimie, 91(6), 784 – 790.
Niwano, T., Terazawa, S., Nakajima, H., & Imokawa, G. (2015). Astaxanthin inhibits the UVA – induced expression of matrix metalloproteinase – 1 in human dermal fibroblasts. Marine Drugs, 13(10), 6220 – 6235.
Palmer, C. N., Irvine, A. D., Terron – Kwiatkowska, A., Zhao, Y., Liao, H., Lee, S. P., & McLean, W. I. (2006). Common loss – of – function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nature Genetics, 38(4), 441 – 446.
Proksch, E., Brandner, J. M., & Jensen, J. M. (2008). The skin: an indispensable barrier. Experimental Dermatology, 17(12), 1063 – 1072.
Rawlings, A. V., & Harding, C. R. (2004). Moisturization and skin barrier function. Dermatologic Therapy, 17(s1), 43 – 48.
Sandilands, A., Sutherland, C., Irvine, A. D., & McLean, W. I. (2009). Filaggrin in the frontline: role in skin barrier function and disease. Journal of Cell Science, 122(9), 1285 – 1294.
Simon, M., Bernard, D., Minondo, A. M., & Schmidt, R. (2001). Persisting corneodesmosomes in the outer stratum corneum of dry skin: a quantitative electron microscopic study. Journal of Investigative Dermatology, 116(1), 23 – 30.
Tominaga, K., Hongo, N., Karato, M., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43 – 47.
Uchida, Y., & Holleran, W. M. (2008). Omega – O – acylceramide, a lipid essential for mammalian survival. Journal of Dermatological Science, 51(2), 77 – 87.
Verdier – Sevrain, S., & Bonté, F. (2007). Skin hydration: a review on its molecular mechanisms. Journal of Cosmetic Dermatology, 6(2), 75 – 82.
Yamashita, E. (2006). The effects of a dietary supplement containing astaxanthin on skin condition. Carotenoid Science, 10, 91 – 95.
Yoshihisa, Y., Rehman, M. U., & Shimizu, T. (2014). Astaxanthin inhibits ultraviolet – induced inflammation and DNA damage in human keratinocytes. Experimental Dermatology, 23(1), 178 – 183.

# KNOWLEDGE SUMMARY: CHAPTER 5 – THE STRUCTURAL TURNOVER AND BARRIER RESTORATION
## I. THE EXOGENOUS ACID FRAUD [THE BIOLOGICAL TRAP]
* **The Mechanism of Aggression:** Chemical exfoliation (AHA/BHA) functions by lowering surface pH to levels incompatible with cellular adhesion, physically melting the **Lipid Mortar** and **Corneodesmosome** protein staples.
* **The “Instant Glow” Illusion:** The immediate post-peel shine is not health; it is the visual result of **Acute Edema** (swelling), **Vasodilation** (redness), and the exposure of the un-keratinized, highly vulnerable lower epidermal layers.
* **The TEWL Catastrophe:** Stripping the **Stratum Corneum** causes an immediate, uncontrolled spike in **Trans-Epidermal Water Loss**, dehydrating the underlying matrix and stalling the 28-day biological conveyor belt.
* **The Rebound Mechanism:**
– **Cytokine Alarm:** Barrier breach triggers a flood of **IL-1** and **TNF-alpha**.
– **NF-kB Activation:** Pro-inflammatory signals prime the skin for a defensive state.
– **Basal Panic:** Melanocytes perceive a “hole in the roof” and initiate **Hyper-melanogenesis**.
– **Result:** The skin heals thicker, darker, and more pigmented (Post-Inflammatory Hyperpigmentation Rebound).
## II. THE PATHOLOGY OF MELANIN ENTRAPMENT [STAGNATION]
* **The Structural Bolts:** **Corneocytes** (dead, melanin-stained cells) are held together by **Corneodesmosomes**—microscopic protein rivets composed of cadherin families.
* **Retention Hyperkeratosis:** In aging skin, these rivets refuse to break, causing a pile-up of 20–30 layers of dead cells, which creates an ashen, “leathery” surface that smothers the **Luminous Matrix**.
* **Enzymatic Paralysis:** Desquamation (shedding) requires **Serine Proteases** (KLK5, KLK7, SCCE). These “Biological Scissors” are strictly **Water-Activated** and **Fluid-Dependent**.
* **The Stagnant Climate:** Chronic dehydration (due to barrier failure) and **Lipid Crystallization** (lack of flexible fats) paralyze these enzymes, preventing them from migrating to and snipping the protein rivets.
## III. LINOLEIC ACID (LA): THE CERAMIDE ARCHITECT
* **Systemic Delivery:** LA is delivered via the repaired **DPA Capillary Network** directly to the **Stratum Granulosum** manufacturing centers.
* **O-Acylceramide Forging:**
– **Enzymatic Binding:** LA is bonded to a **Sphingosine Backbone** by ceramide synthases.
– **Ceramide EOS (Ceramide 1):** LA facilitates the creation of this ultra-rare, long-chain lipid that acts as a molecular bridge across the matrix.
* **Lamellar Stacking:** These specific ceramides organize into dense, impenetrable sheets, physically plugging micro-fissures in the “roof” of the skin.
* **Signal Silence:** Once the waterproof seal is restored, **TEWL** terminates. Keratinocytes cease their panic signaling, allowing **Basal Melanocytes** to enter a state of biological relief and power down tyrosinase production.

## IV. OLEIC ACID (OA): THE FLUIDITY CATALYST
* **The Crystallization Problem:** Aging skin is dominated by saturated fatty acids (**Palmitic/Stearic**) that pack into a rigid, solid-gel “Frozen Phase,” blocking enzymatic activity.
* **The Cis-Double Bond Kink:** OA possesses an 18-carbon chain with a permanent physical “elbow” or “bend” at the ninth carbon.
* **The Thermodynamic Phase Shift:**
– **Spatial Disruption:** The “kinked” OA molecules elbow their way between straight-chain saturated fats, pushing them apart.
– **Liquid-Crystal Shift:** This dramatically lowers the **Phase Transition Temperature**, melting the solid matrix into a flexible, fluid liquid-crystal state.
* **Enzymatic Awakening:** In this fluid and water-rich environment, the dormant **Proteases** regain 3D mobility. They migrate to the protein rivets, snip them apart, and allow the melanin-stained cells to slough off naturally.
## V. ASTAXANTHIN’S ESCORT: THE PEROXIDATION SHIELD
* **The Vulnerable Window:** LA and OA must migrate upward through the living layers for 28 days before reaching the surface.
* **The Singlet Oxygen Threat:** UVA/UVB radiation penetrates this migration zone, targeting the fragile double bonds of the lipids.
* **Lipid Peroxidation:** Unprotected lipids shatter into **Malondialdehyde** (toxic waste), which destroys the barrier rebuild and re-triggers inflammation.
* **The Transmembrane Armor:** **Astaxanthin** (6,000x Vitamin C potency) anchors into the membranes of migrating keratinocytes, traveling alongside the LA/OA matrix.
* **Pristine Delivery:** Its massive electron cloud quenches UV energy before it can strike, ensuring the structural building blocks arrive at the surface 100% intact.
## VI. THE FINAL VERDICT [AESTHETIC SOVEREIGNTY]
* **The Internal Push:** Instead of “pulling” skin off with acid (Exogenous), the Luminous Matrix “pushes” dead skin off through internal growth (Endogenous).
* **The Biological Reveal:** Without chemical burns or downtime, the skin sheds its history of trauma (dark spots/leathery texture) to reveal clear, translucent, and unpigmented tissue.
* **Aesthetic Sovereignty:** Achieving a glass-like finish through biological elegance and structural integrity rather than surface-level destruction.

Chatper 6: The 7-Molecule Luminous Matrix:
The Mathematical Impossibility Of Isolation
Integrating Astaxanthin’s genetic blockade with the Omega lipidomic sweep to achieve absolute, system-wide radiance.
In the Scientific Noir of the modern aesthetic landscape, we are witnessing a multi – billion dollar paradox.
Consumers are spending unprecedented amounts on isolated, high – potency molecules – such as Vitamin C, Niacinamide, or Tranexamic Acid – yet the prevalence of chronic dullness, sallow yellowness, and stubborn post – inflammatory hyperpigmentation remains at an all – time high.
This failure is not due to a lack of chemical potency, but a fundamental misunderstanding of cutaneous systems biology. The cosmetic industry has sold the public on the “Magic Bullet” fallacy: the reductionist idea that a single ingredient can solve a multi – dimensional biological crisis.
In reality, absolute skin luminosity is not a static state achieved by blocking a single enzyme; it is a massive, multi – system biological concert that requires the simultaneous coordination of genetic signaling, immunological clearance, hemodynamic flow, and structural rebuilding.
You cannot play a complex symphony with a single instrument, no matter how loud that instrument is played. When you apply a single – ingredient brightener to the surface, you are essentially asking a solo violinist to drown out the noise of a chaotic, broken city.
True radiance is an emergent property that only arises when every biological “instrument” – from the deep capillary loops to the surface lipid barrier – is tuned to the same frequency of health.
The Keyora matrix represents the transition from noise to harmony, integrating seven specific molecules into a systemic architecture that treats the skin as a unified, living field rather than a collection of isolated symptoms.

A. The Single – Ingredient Myth
The thermodynamic and biochemical failure of isolated interventions.
The flaw of reductionist skincare lies in its attempt to simplify a non – linear system.
Skincare brands market “hero ingredients” because they are easy to explain and even easier to manufacture, but biology does not respect marketing simplicity.
When you introduce a high – dose isolated molecule into the skin, you are creating a biochemical imbalance that the body’s homeostatic mechanisms will inevitably attempt to correct.
This isolated intervention ignores the “Pathological Web” – the interconnected network of triggers that sustain hyperpigmentation.
By focusing on a single node in this web, the industry fails to account for the upstream signals that initiate the problem and the downstream debris that sustains the visual shadow.

B. The Biochemical Isolation
Most isolated brightening ingredients are designed to be tyrosinase inhibitors, meaning they only target the final enzymatic step of melanin production.
While this may temporarily slow down the darkening process, it completely ignores the upstream genetic triggers and the massive release of pro – inflammatory cytokines that tell the melanocyte to produce pigment in the first place.
If the skin is still in a state of “Code Red” inflammation, the melanocyte will simply wait for the inhibitor to wear off or find an alternative pathway to express its defensive pigment. Treating the enzyme without silencing the signal is a strategic failure that results in temporary results followed by chronic frustration.
Firstly, The Vascular Blindness:
Surface – level, single – ingredient interventions suffer from a profound lack of depth. They are physically and biologically incapable of addressing the microvascular collapse and hemodynamic stagnation that occur deep within the dermis.
A significant portion of what we perceive as “dullness” is actually hypoxic yellowing caused by a lack of oxygenated blood flow. No amount of topical Niacinamide can rebuild a shriveled capillary loop or flush out deoxygenated venous waste.
By ignoring the vascular component of luminosity, the industry leaves the skin’s “internal backlight” turned off, making true radiance biologically impossible to achieve.
Secondly, The Inevitable Rebound:
Biology is a web of compensatory loops.
When you aggressively block one specific node in a complex network without supporting the surrounding nodes, the system creates a “rebound reaction” to restore its perceived equilibrium.
For example, stripping the skin with acids to remove pigment often triggers an inflammatory surge that tells the melanocytes to produce even more melanin to protect the now – vulnerable tissue.
This is why many individuals find that their dark spots return even darker after using isolated chemical lighteners. The system was never stabilized; it was merely provoked, leading to a cycle of temporary clearing and permanent darkening.

C. The Systemic Requirement
To achieve absolute clarity, the Keyora protocol does not merely inhibit an enzyme; it seeks to shut down the melanin factory at its genetic and signaling source. This requires the use of molecules that can cross the cellular membrane and interact with the transcription factors that govern pigment expression.
By calming the “Panic Signal” at the basal layer, we prevent the melanocyte from entering its defensive posture, ensuring that the blockade is stable and long – term rather than a temporary chemical interference.
Firstly, The Immunological Clearance:
Radiance cannot exist in a field of debris. When inflammation occurs, the skin is left with “Melanophage Corpses” – dead immune cells engorged with trapped pigment. Isolated brighteners have zero mechanism for clearing this deep – seated waste.
The Keyora matrix utilizes specialized pro – resolving mediators to activate the skin’s “cleanup crew,” forcing macrophages to consume and permanently remove this internal soot via the lymphatic system. This immunological sweep is the only way to clear the persistent, ashen shadows that topical creams cannot reach.
Secondly, The Hemodynamic Flush:
A truly luminous complexion requires a constant supply of oxygenated, bright scarlet blood. The systemic requirement for radiance includes the physical rebuilding of the micro – plumbing network.
By utilizing DPA and Astaxanthin to restore capillary loops and induce a hemodynamic surge, the Luminous Matrix physically flushes out the stagnant yellow swamp of the dermis.
This “Internal Power Wash” replaces the muddy tones of deoxygenation with the vibrant pink undertones of high – performance health, providing the essential backlight for the skin.
Thirdly, The Structural Turnover:
The final dimension of the systemic requirement is the restoration of the biological conveyor belt.
The surface of the skin must be able to shed its dark, melanin – stained cells naturally and effortlessly.
This is achieved by rebuilding the ceramide seal and fluidizing the lipid matrix with Linoleic and Oleic acids.
When the structural foundation is secure, the skin naturally “pushes” the darkness off from the inside out, revealing a clear, translucent surface without the need for the inflammatory destruction of chemical peels.

6.1 The Commander:
Astaxanthin
The Absolute Apex Predator Of The Melanin Factory, Executing A Simultaneous Strike Across Three Distinct Biological Dimensions.
In the Scientific Noir of the Luminous Matrix, the role of the commander is reserved for the molecule that possesses the highest level of thermodynamic authority and the broadest reach across the cellular landscape. This molecule is Astaxanthin.
In the Keyora 7 – molecule architecture, Astaxanthin is not a mere additive or a supporting antioxidant; it is the absolute apex predator of the melanin factory.
Most brightening agents are fragile, surface – bound molecules that struggle to survive the journey to the basal layer, but Astaxanthin is a systemic powerhouse that executes a simultaneous strike across three distinct biological dimensions: genetic, enzymatic, and thermodynamic. It does not just “help” the skin look better; it dictates the internal climate of the entire epidermal unit, forcing the hyper – active pigment machinery into a state of absolute submission.
By utilizing its unique 16 mg dosage and its unmatched capacity for singlet oxygen quenching – which is 6,000 times more potent than traditional Vitamin C – Astaxanthin provides the structural and chemical cover required for every other molecule in the matrix to perform its function.
Without the commander, the other six molecules would be consumed by the oxidative fire of the dermal environment. With the commander, the melanin factory is systematically dismantled from the nucleus to the cell membrane, ensuring a state of permanent, deep – tissue clarity.

Phase 1: The Genetic Intervention
Silencing the master commander in the nucleus.
The first strike executed by Astaxanthin occurs at the most fundamental level of cellular instruction: the genome.
To stop hyperpigmentation permanently, one must stop the “Panic Signal” that travels from the surface to the nucleus of the melanocyte. This is the genetic intervention, where Astaxanthin acts as a molecular silencer, preventing the transcription of the blueprints required for melanin production.
I. The Intracellular Penetration:
Unlike water – soluble antioxidants that are blocked by the cellular lipid barrier, Astaxanthin is a highly lipophilic molecule that effortlessly breaches the melanocyte cytoplasm. Its unique molecular geometry allows it to pass through the cell membrane and enter the intracellular space where the signaling cascades originate.
Once inside, Astaxanthin establishes a high – concentration defensive perimeter, ready to intercept the chemical messages that tell the cell to darken.
This deep penetration is what allows Astaxanthin to influence the internal logic of the cell, moving beyond surface – level suppression to reach the core of the manufacturing process.
II. The Kinase Interception:
When the skin is stressed by UV light or inflammation, a series of signaling proteins known as Mitogen – Activated Protein Kinases (MAPK) are activated. These kinases act as the cellular telegraph, carrying the “produce melanin” command toward the nucleus.
Astaxanthin executes a precision Kinase Interception, specifically inhibiting the phosphorylation of p38 and JNK pathways.
By degrading these upstream stress signals before they can reach the nuclear envelope, Astaxanthin prevents the “Panic Signal” from ever being delivered. It effectively cuts the communication lines between the stressed surface of the skin and the manufacturing center below.
III. The MITF Downregulation:
The ultimate target of the genetic intervention is the Microphthalmia – Associated Transcription Factor (MITF).
MITF is the “Master Switch” for melanin production; when it is active, it binds to the DNA and authorizes the production of tyrosinase and other pigment – making tools. Astaxanthin suppresses the expression of MITF, fundamentally revoking its authority to command the cell.
By keeping the MITF switch in the “off” position, Astaxanthin ensures that the DNA blueprints for melanin are never read or translated. This cuts the power to the factory at the source, preventing the birth of new pigment molecules before they can even be synthesized.

Phase 2: The Enzymatic Arrest
Confiscating the manufacturing equipment.
Even with the genetic switch turned off, existing enzymes within the melanocyte can still produce pigment if they are not neutralized.
This is why the commander executes a secondary strike: the enzymatic arrest.
In this phase, Astaxanthin physically moves into the “assembly line” to confiscate the manufacturing equipment, ensuring that no further chemical conversion of tyrosine can take place.
I. The Active Site Docking:
Tyrosinase is the primary enzyme responsible for the conversion of the amino acid tyrosine into melanin. This enzyme possesses a specific pocket called the “Active Site” where the chemical reaction occurs.
Astaxanthin performs a precision Active Site Docking maneuver, utilizing its specific molecular orientation to occupy the pocket. By filling this space, it prevents tyrosine from entering the enzyme.
This is a form of competitive inhibition that physically blocks the machinery of the cell, ensuring that even if the enzyme is present, it is rendered useless and inert.
II. The Conformational Shift:
Beyond simple blocking, the binding of Astaxanthin induces a Conformational Shift in the Tyrosinase protein structure. The enzyme is a complex, 3D folded molecule that relies on a very specific shape to function.
When Astaxanthin docks with the protein, it forces the entire structure to “stiffen” and change shape.
This paralyzes the enzyme’s physical machinery, making it chemically impossible for the protein to catalyze any reactions. The manufacturing equipment is not just blocked; it is physically broken at a molecular level, leading to a total cessation of enzymatic output.
III. The DOPA Blockade:
Melanin production is a multi – step chemical process, with the conversion of Tyrosine to DOPA and then to Dopaquinone being the most critical.
By paralyzing the Tyrosinase enzyme, Astaxanthin creates an absolute DOPA Blockade.
Without the catalytic power of the enzyme, the chemical chain reaction grinds to a halt.
The “raw materials” of pigment simply sit in the cell as harmless amino acids, unable to evolve into the dark, polymerized granules that create visible spots on the skin. This blockade ensures that the visual “darkening” of the skin is stopped dead in its tracks.
IV. The Melanosome Embargo:
Once melanin is created, it is packaged into tiny shipping containers called melanosomes and transferred to the surrounding skin cells.
Astaxanthin interferes with this transport process, creating a Melanosome Embargo. It disrupts the cytoskeletal “highways” that these containers use to move toward the cell membrane.
By preventing the transfer of whatever dark pigment might still be left in the factory, Astaxanthin ensures that the pigment stays trapped inside the melanocyte where it can be naturally broken down by the cell’s own recycling center (the lysosome).
This prevents the “spreading” of the shadow to the surface layers of the skin.

Phase 3: The Thermodynamic Shield
Starving the engine of its oxidative fuel.
The final and most powerful role of the commander is the establishment of the thermodynamic shield. Melanin production is a “hot” reaction that requires oxidative energy to drive the polymerization of pigment.
By removing the “fire” of free radicals and singlet oxygen from the cellular environment, Astaxanthin starves the pigment engine of the fuel it needs to operate, while simultaneously protecting the rest of the 7 – molecule matrix.
I. The Transmembrane Saturation:
Astaxanthin possesses a unique molecular architecture – featuring two polar end groups and a long, non – polar polyene chain – that allows it to anchor itself across the entire width of the cellular lipid bilayer.
This is the Transmembrane Saturation.
Unlike other antioxidants that only protect the inside or the outside of the cell, Astaxanthin protects both.
It creates a 360 – degree thermodynamic firewall that coats every melanocyte and keratinocyte in the basal layer. This structural integration ensures that no oxidative stress can breach the cell from any direction, providing absolute safety for the internal organelles.
II. The Unrelenting Quench:
The commander’s primary weapon is its massive electron cloud, which allows it to act as an Unrelenting Quench for singlet oxygen. Singlet oxygen is a high – energy, “vibrating” form of oxygen that is the primary catalyst for hyperpigmentation and dermal decay.
Astaxanthin “vacuums up” the energy from these reactive species and dissipates it as harmless heat, a process it can repeat thousands of times without being destroyed itself.
This is the ultimate thermodynamic reset; by quenching the oxidative fire, Astaxanthin removes the chemical pressure that forces the skin to darken in the first place.
III. The Absolute Deprivation:
The establishment of the shield leads to a state of Absolute Deprivation for all pathological processes.
Tyrosinase requires oxidation to work; Lipofuscin requires oxidative cross – linking to form; the inflammatory alarm (NF – kB) requires oxidative triggers to fire.
By stripping the environment of its oxidative fuel, Astaxanthin renders these processes biologically impossible.
The skin is returned to its “Cool” baseline state, allowing the Luminous Matrix to focus its energy on repair and radiance rather than defense and pigment. This thermodynamic sovereignty is the foundation upon which absolute luminosity is built, ensuring that the skin remains clear, bright, and resilient forever.

6.2 The Purifiers:
ALA To EPA And DHA
Deploying the biochemical cleanup crew to phagocytize dermal debris and stabilize the neuro – inflammatory baseline.
In the Scientific Noir of the human matrix, shutting down the melanin factory is only the first phase of the restoration of the Luminous Matrix.
While the Commander (Astaxanthin) has seized control of the genetic and enzymatic machinery, the dermal landscape remains littered with the visual wreckage of past conflicts.
This wreckage manifests as stubborn, ashen shadows and deep – seated purplish marks – technically known as post – inflammatory hyperpigmentation (PIH) – which are the biological residue of old acne breakouts, allergic reactions, and localized trauma.
These marks are not surface stains; they are engorged “melanophage corpses” – stationary immune cells that have swallowed so much pigment that they have died in place, becoming permanent ink – spots trapped within the collagen matrix.
To clear this deep – seated “internal soot,” we must deploy the Omega – 3 purifiers.
By supplying a massive systemic dose of Alpha – Linolenic Acid (ALA) from the Keyora matrix, we initiate a hepatic and cellular conversion process that generates a high – concentration flux of Eicosapentaenoic Acid (EPA) and Docosahexaenoic Acid (DHA).
These molecules act as the skin’s elite immunological cleanup crew, executing a highly coordinated sweep of the deep dermis to physically consume the debris while simultaneously stabilizing the nervous system’s inflammatory baseline.

A. The EPA Sweep
Eradicating the post – inflammatory hyperpigmentation (PIH).
The first objective of the purifiers is the total eradication of the “dermal graveyard” of pigment.
This mission is spearheaded by Eicosapentaenoic Acid (EPA), which serves as the direct precursor to a class of powerful molecules known as Specialized Pro – resolving Mediators (SPMs).
These molecules do not merely “reduce” inflammation; they actively force the local immune environment into a state of resolution and cleanup, turning the skin from a site of chronic stagnation into a high – efficiency biological waste – removal system.
Firstly, The Resolvin Synthesis:
Once the EPA reaches the site of a persistent dark mark via the newly restored microvascular network, it is metabolized by specialized enzymes into Resolvin E1 (RvE1).
This is the “Resolvin Synthesis” phase. Unlike traditional anti – inflammatory drugs that simply block pathways, Resolvins act as active signaling molecules that transmit the “Clearance Code” to the surrounding tissue.
They bind to the ChemR23 receptors found on the surface of local cells, essentially ringing a biological dinner bell that alerts the immune system to the presence of rotting melanophage corpses that need to be removed.
Secondly, The Macrophage Polarization:
The most critical tactical shift orchestrated by EPA is the “Macrophage Polarization.”
In a state of chronic PIH, the local immune cells are stuck in a “Pro – inflammatory M1” state, where they continue to release toxic cytokines that irritate the skin. The presence of Resolvins forces these cells to undergo a radical identity shift into the “Pro – resolving M2” phenotype.
M2 macrophages are the skin’s dedicated janitors and architects. By shifting the M1/M2 ratio in favor of the M2 phenotype, the EPA matrix ensures that the skin’s immune system stops fighting and starts cleaning, providing the necessary workforce to begin the physical extraction of trapped pigment.
Thirdly, The Physical Phagocytosis:
The M2 macrophages, now activated by the EPA – derived Resolvins, begin the “Physical Phagocytosis” of the dermal graveyard. These specialized immune cells physically crawl through the collagen fibers until they locate the dead, ink – filled melanophages.
Upon contact, the M2 macrophage extends its cellular arms around the pigment corpse and swallows it whole. Inside the macrophage, powerful lysosomal enzymes are deployed to break down the dense, polymerized melanin granules into smaller, soluble particles.
This is a forensic cleanup operation that physically removes the source of the shadow from the deep dermal compartment.
Fourthly, The Lymphatic Flush:
The final stage of the EPA sweep is the “Lymphatic Flush.”
Once the M2 macrophages have finished digesting the internal soot, they do not stay in the skin. Guided by chemical gradients, these pigment – loaded cells migrate toward the nearest lymphatic vessel.
They enter the lymphatic system, carrying the digested dark pigment away from the face and into the systemic waste stream for permanent excretion.
This is why the Keyora matrix can erase acne marks that have persisted for years; it is not “bleaching” the spot, but physically carrying the ink out of the tissue through the body’s own drainage network, restoring absolute dermal clarity.

B. The DHA Stabilization
Calming the neurogenic sensitivity of the skin.
While EPA clears the physical debris, Docosahexaenoic Acid (DHA) works to secure the “Neuro – Barrier” of the skin.
In the Scientific Noir of the matrix, chronic hyperpigmentation is often sustained by “Neurogenic Inflammation” – a state where hypersensitive nerve endings misfire and release neuropeptides that keep the skin in a state of perpetual irritation.
DHA acts as the ultimate neural stabilizer, providing the structural integrity required to calm these signals and ensure that the cleanup process remains uninterrupted.
Firstly, The Neural Integration:
DHA is the primary structural fatty acid of the human nervous system, and the skin is one of the body’s most densely innervated organs.
Once DHA is synthesized from the ALA precursor, it undergoes “Neural Integration,” physically incorporating itself into the phospholipid bilayers of the peripheral nerve endings and the Schwann cells that insulate them.
This integration increases the fluidity and electrical stability of the nerve membranes, ensuring that the sensory fibers in the dermis are resilient and well – insulated against the chaotic signals of environmental stress.
Secondly, The Hyper – Reactivity Dampening:
A DHA – enriched nerve membrane is far less likely to undergo spontaneous depolarization. This results in “Hyper – Reactivity Dampening.”
In a state of DHA deficiency, even minor triggers like heat, friction, or emotional stress can cause these nerves to release a flood of Substance P and Calcitonin Gene – Related Peptide (CGRP), both of which trigger the melanocyte to produce more pigment.
By stabilizing the ion channels within the nerve membrane, DHA prevents these “false alarms,” effectively silencing the neurogenic background noise that often leads to mysterious, stress – induced allergic reactions and inflammatory flare – ups.
Thirdly, The Baseline Calm:
The final outcome of this neural fortification is the “Baseline Calm.” By securing the neuro – inflammatory baseline, DHA ensures that the EPA sweep can proceed flawlessly.
The skin is no longer a reactive, sensitive battlefield; it becomes a stable, high – performance field where repair can happen without the constant threat of a new inflammatory surge.
This baseline calm is essential for the long – term maintenance of the Luminous Matrix, ensuring that once the dark marks are cleared, they do not return, and the skin remains in a state of permanent, serene clarity.

6.3 The Illuminator:
ALA To DPA
Re – engineering the microvascular plumbing to physically flush out hypoxic yellowness with high – pressure arterial blood.
In the Scientific Noir of the Luminous Matrix, we must confront a biological reality that surface – level treatments have ignored for decades: the debris field of the dermis may be cleared by the purifiers, but the skin will remain a stagnant, yellow swamp as long as the microvascular plumbing remains in a state of collapse.
This condition, known as “Vascular Ghosting,” occurs when the tiny capillary loops that once reached up into the dermal papillae shrivel and retreat, leaving the upper dermis in a state of chronic hypoxia.
Without a constant supply of fresh, oxygenated blood, the skin cannot reflect light; it instead absorbs it, projecting a sickly, sallow, and exhausted tone that no topical cream can erase.
To resolve this, we must introduce the master micro – engineer of the Omega – 3 family: Docosapentaenoic Acid (DPA).
While EPA and DHA focus on cleanup and stabilization, DPA is specifically synthesized from the systemic Alpha – Linolenic Acid (ALA) matrix to execute a high – precision mission of neo – angiogenesis.
DPA is the biological key that unlocks the growth of new blood vessels, bringing the “River of Life” back to the surface and physically flushing out the “Swamp of Stagnation.”
This is the ultimate hemodynamic reset, providing the internal backlight that is the absolute prerequisite for glass – like, translucent radiance.

Step 1: The Vascular Sprout
Laying down the new biological pipes.
The reconstruction of the skin’s irrigation system is a complex, multi – stage engineering project that requires the precise coordination of cellular growth signals and structural assembly.
DPA does not merely “improve” blood flow; it initiates the physical creation of new conduits, sprouting a fresh network of vertical capillary loops that reclaim the territory lost to aging and oxidative stress.
This process of vascular sprouting is what allows the Luminous Matrix to permanently bypass the blockages of the past and establish a new foundation for radiant health.
I. The Endothelial Rescue:
The first phase of the vascular sprout begins with the stabilization of the existing, fragmented microvasculature.
As DPA is synthesized and delivered to the dermal matrix, it physically integrates into the phospholipid membranes of the dying endothelial cells. These cells, which form the inner lining of every blood vessel, are often in a state of metabolic collapse.
DPA acts as a molecular life – support system, downregulating the activity of Caspase – 3 and halting the process of programmed cell death (apoptosis).
By rescuing these “Seed Cells,” DPA preserves the essential biological starting points required for the expansion of the plumbing network.
II. The Vegf Upregulation:
Once the foundation is stabilized, DPA triggers the massive release of Vascular Endothelial Growth Factor (VEGF) from the surrounding fibroblasts and the rescued endothelial cells themselves.
In the hierarchy of biological signaling, VEGF is the “Master Blueprint” for vessel creation. DPA is clinically unique in its ability to stimulate VEGF with a potency that far exceeds that of EPA or DHA.
This chemical signal radiates through the hypoxic zones of the dermis, marking the areas where new capillary loops are desperately needed and recruiting the cellular workforce required to build them.
III. The Cellular Migration:
Under the command of the VEGF signal, the rescued endothelial cells begin to divide and multiply, initiating a process of physical cellular migration. These cells do not stay stagnant; they use specialized enzymes called matrix metalloproteinases to gently dissolve a path through the dense collagen fibers of the dermis.
Moving with forensic precision, they crawl upward toward the epidermis, following the chemical “Scent” of the growth factors. This upward movement is the first physical sign of the Luminous Matrix’s internal resurrection, as the skin prepares to bring its blood supply back to the highest levels of the tissue.
IV. The Tube Formation:
As the migrating endothelial cells reach their target destinations, they begin the remarkable process of tube formation.
They organize themselves into hollow, three – dimensional structures, wrapping their membranes around a central lumen to create a functional pipe. This is not a disorganized clump of cells, but a precision – engineered biological conduit capable of carrying fluid.
DPA ensures that these new tubes are flexible and resilient, providing the structural elasticity needed to withstand the incoming surge of arterial pressure without leaking or collapsing.
V. The Dermal Penetration:
The final act of the vascular sprout is the successful dermal penetration of the new capillary loops.
These pipes reach right up to the highest peaks of the dermal papillae, just micrometers away from the basal layer of the epidermis. This is the “Strategic High Ground” of cutaneous health.
By penetrating these peaks, the new loops are positioned to deliver oxygen and nutrients directly to the cells responsible for skin renewal. The plumbing is now complete; the pipes are laid, and the skin is ready for the final, cleansing rush of the arterial flush.

Step 2: The Arterial Flush
The physical and optical eradication of the yellow swamp.
The pipes are built, the connections are made, and the biological valves are now ready to open. This is the moment of the Arterial Flush, a physical and optical transformation that represents the absolute triumph of systems biology over cosmetic dullness.
As the heart pumps fresh, oxygenated blood into the newly engineered DPA loops, the skin undergoes a rapid qualitative shift.
The murky, yellow – gray “Swamp of Stagnation” is physically evicted and replaced by a vibrant, scarlet tide of life, resulting in a visual reveal that is the signature of the Luminous Matrix.
I. The Hemodynamic Surge:
The transition begins with the hemodynamic surge, as the full force of the systemic circulation is directed into the newly minted capillary network. This is a high – pressure event that physically inflates the dermis from within.
For the first time in years, the “Biological Water Faucets” under the face are turned on at maximum capacity.
This surge of fresh arterial blood brings with it the thermal energy and metabolic fuel required to restart the skin’s dormant repair processes, acting as a powerful biological wake – up call for every cell in the epidermal unit.
II. The Venous Eviction:
As the fresh arterial blood fills the new loops, it creates a massive pressure gradient that triggers the venous eviction.
This high – pressure surge physically forces the stagnant, deoxygenated, and purple – yellow “Swamp Water” out of the tissue and back into the systemic venous return.
The deoxygenated hemoglobin and metabolic waste products that have been pooling in the dermis – creating that muddy, sallow undertone – are literally power – washed away.
This internal cleansing is the only way to achieve absolute clarity; the “Yellow Swamp” is not masked by makeup, but is physically removed from the tissue.
III. The Oxyhemoglobin Loading:
With the stagnant waste removed, the superficial dermis experiences the arrival of absolute oxyhemoglobin loading.
The new DPA – built loops are now packed with bright, scarlet – red, oxygen – rich hemoglobin molecules. In the world of optics, oxyhemoglobin is a brilliant chromophore that radiates health.
This red light is the “Natural Blush” of youth, providing the skin with a warm, peachy undertone that signals a high – performing cardiovascular and metabolic state.
The dermis has been transformed from a dark, hypoxic cellar into a vibrant, oxygen – saturated field of life.
IV. The Translucent Radiance:
The final visual outcome of the flush is the birth of translucent radiance.
Because the epidermis has been cleared and the collagen matrix is now hydrated and organized, the bright scarlet hue of the new blood supply projects upward with high – definition clarity.
This red light passes through the semi – transparent layers of the skin, instantly neutralizing the residual yellowness of the tissue and creating an un – fakeable, glass – like inner glow.
The skin no longer looks flat or “Painted”; it looks three – dimensional and lit from within. This is the Luminous Matrix in its peak state – a complexion that projects the absolute reality of its own internal vitality to the world.

6.4 The Restorers:
LA And OA
Rebuilding the ceramide roof to silence the basal panic and restoring liquid-crystal fluidity to naturally shed the dark surface leaves.
In the Scientific Noir of the human epidermis, we must recognize that while the foundation has been perfectly engineered through microvascular sprouting and immunological purging, the architectural project remains incomplete as long as the roof is broken and the surface is cluttered with the debris of the past.
The Luminous Matrix cannot project its internal brilliance if the stratum corneum is a shattered, leaky mess of micro – fissures and the visual field is blocked by a thick, stagnant layer of dark, dead cells.
To resolve this structural crisis, we must deploy the final pair of restorers: Linoleic Acid (LA) and Oleic Acid (OA).
These fatty acids are not merely “moisturizers” in the traditional cosmetic sense; they are high – performance building blocks and thermodynamic catalysts that work in tandem to repair the barrier and restart the stalled biological conveyor belt.
By providing the specific raw materials required for O – acylceramide synthesis, we physically seal the water leak that triggers the melanocyte’s defensive panic.
Simultaneously, we use the unique molecular geometry of Oleic Acid to shatter the rigid lipid crystallization that traps dark pigment on the surface.
This is the ultimate structural reset, ensuring that the skin not only holds its moisture but also regains its ability to shed its own darkness, revealing the pristine, luminous tissue that has been hidden beneath a leathery mask of stagnation.

I. The Ceramide Assembly
Sealing the leak and calming the panic.
The first objective of the restorers is to perform a total reconstruction of the skin’s primary defensive barrier. This is the Ceramide Assembly phase, where Linoleic Acid is utilized to forge the most sophisticated waterproof lipids in human biology.
Without this seal, the skin is in a state of permanent emergency, constantly losing water to the atmosphere and sending frantic signals to the basal layer to produce protective pigment.
By repairing the roof, we do more than just hydrate the tissue; we biochemically silence the alarms that drive hyperpigmentation.
Firstly, The O – Acylceramide Synthesis:
Linoleic Acid is the non – negotiable architect of the skin’s ultimate waterproof mortar.
Within the granular layer of the epidermis, the systemic supply of LA is enzymatically conjugated with a long – chain sphingosine base to create O – acylceramides, specifically Ceramide EOS.
This molecule is distinguished by its extreme length and structural complexity, acting as a molecular bridge that spans across multiple lipid bilayers.
Without a sufficient supply of LA, the body is forced to produce shorter, inferior ceramides that are structurally incapable of forming an effective barrier.
This “Architectural Synthesis” ensures that the lipid mortar is of the highest biological grade, providing the necessary tensile strength to maintain the skin’s integrity against the oxidative vacuum of the environment.
Secondly, The Lamellar Stacking:
Once the O – acylceramides are synthesized, they must be organized into a functional structure.
Oleic Acid plays a critical role here by providing the necessary fluidity for these complex lipids to arrange themselves into orderly, parallel sheets.
This is the Lamellar Stacking process. The LA – based ceramides provide the structural “bricks,” while the OA – influenced matrix provides the flexible “cement” that allows these sheets to stack into impenetrable, hydrophobic barriers.
This organized stacking is what transforms a disorganized mess of lipids into a precision – engineered waterproof shield, preventing the intrusion of irritants and the escape of vital internal fluids.
Thirdly, The TEWL Termination:
The immediate physical consequence of a successful ceramide assembly is the absolute and measurable halt of Trans – Epidermal Water Loss (TEWL).
By physically plugging the micro – fissures in the stratum corneum, the LA/OA matrix effectively seals the skin’s “Water Leak.”
This TEWL termination creates a high – pressure hydration zone within the epidermis, restoring the turgor and volume of the skin cells.
For the first time, the internal water remains where it belongs, providing the necessary environment for cellular enzymes to function and for the skin to project its natural light without the interference of dry, opaque cracks.
Fourthly, The Signal Silence:
A perfectly sealed roof sends a definitive biochemical signal to the layers below.
When the barrier is compromised, keratinocytes release a constant stream of pro – inflammatory cytokines, such as Interleukin – 1, which act as a “Panic Signal” to the melanocytes.
Once the LA/OA seal is intact, this “Code Red” alarm is silenced.
The melanocytes in the basal layer perceive that the danger has passed and that the organism is no longer vulnerable to environmental radiation or desiccation. In response, they power down their defensive melanin production, permanently halting the creation of new dark spots at the source.

II. The Exfoliation Release
Reactivating the dormant shedding enzymes.
With the roof sealed and the water trapped inside, we must now address the problem of the stagnant, dark surface cells that won’t wash off.
In aging or stressed skin, these melanin – stained cells are held together by protein rivets called desmosomes that refuse to break. This is because the enzymes responsible for cutting these rivets have been paralyzed by a stiff, crystallized lipid environment.
By introducing Oleic Acid, we induce a thermodynamic phase shift that shatters this gridlock and reactivates the skin’s natural “Biological Scissors.”
Firstly, The Phase Shift:
The core obstacle to natural shedding is the rigid, crystallized state of the aging lipid matrix. Saturated fats in the skin tend to pack together in a frozen, solid – gel phase that blocks all enzymatic movement.
Oleic Acid, however, possesses a unique molecular structure featuring a permanent, physical “bend” or “kink” due to its cis – double bond.
When OA enters the matrix, this bent geometry acts as a molecular wedge, shattering the rigid crystalline packing of the saturated fats. This “Phase Shift” returns the lipid mortar to a flexible, liquid – crystal state, creating the fluid paths required for biological work to resume.
Secondly, The Protease Awakening:
In this newly restored liquid – crystal environment, which is also deeply hydrated thanks to the LA seal, the dormant desquamation enzymes – known as serine proteases – are finally able to wake up. These proteases, specifically KLK5 and KLK7, require both fluidity and water to navigate the intercellular spaces and reach their targets.
With the OA – induced phase shift, these “Biological Scissors” regain their 3D mobility. They are now free to move throughout the stratum corneum, seeking out the protein rivets that have been holding the dark, dead cells hostage for months.
Thirdly, The Flawless Reveal:
The final act of the Luminous Matrix is the Flawless Reveal.
The awakened proteases precisely target and cut the protein bonds (desmosomes) holding the dark, melanin – stained dead cells together.
As these bonds are dissolved, the surface debris is finally freed from its anchors.
The dark cells naturally and gently slough off into the environment during daily movement, without the need for harsh chemical peels or physical scrubbing.
This reveals the fresh, clear, and unpigmented luminous tissue beneath, resulting in a glass – like skin surface that is achieved through pure biological elegance and structural integrity.

6.5 The Mathematical Impossibility Of Isolation
A ruthless biological proof of why removing any single molecule from the 7 – part matrix guarantees the total collapse of cutaneous luminosity.
In the Scientific Noir of Keyora Research, we do not view the Seven – Molecule Matrix as a collection of optional supplements; we view it as a high – performance biological circuit where the removal of even a single component results in a total systemic failure.
This is the ultimate proof of the Keyora Matrix, a rigorous deconstruction of why 1 + 1 + 1 + 1 + 1 + 1 + 1 is mathematically and thermodynamically greater than 7.
To understand the absolute necessity of this synergy, we must perform a forensic reverse – engineering thought experiment.
By removing one molecule at a time from the equation, we can observe how the entire architecture of luminosity dissolves into a state of pathological stagnation. The skin is a non – linear, complex adaptive system that does not respond to isolated signals.
When you treat only one node of the network, the remaining untreated nodes compensate with a biological retaliation that ensures the return of dullness and hyperpigmentation.
This section provides the ruthless proof that true radiance is an emergent property that can only exist when every biological instrument is played in perfect synchronicity.

Scenario A:
The DPA Vacuum
The pale, suffocating desert.
In this first forensic scenario, we assume a state where the Commander, Astaxanthin, has successfully executed a total genetic and enzymatic blockade. Melanin production has been halted at the source. The factory is silent, and the skin is technically “brightened” in terms of pigment density.
However, in this scenario, we have removed Docosapentaenoic Acid (DPA) from the matrix.
This creates the DPA Vacuum – a state where the skin’s microvascular plumbing remains in a state of terminal collapse.
I. The Melanin Success:
Through the high – dose administration of Astaxanthin, we have successfully silenced the MITF gene and paralyzed the Tyrosinase enzyme. The skin cells are no longer being flooded with dark pigment.
From a purely pigmentary perspective, the treatment is a success. The dark spots have faded, and the “brown” has been removed from the visual palette of the face.
However, this pigmentary victory is hollow because it addresses only the filter of the skin, not the backlight that is supposed to shine through it.
II. The Vascular Failure:
Without the presence of DPA, the master micro – engineer, the skin is unable to sprout new capillary loops or rescue the dying endothelial cells in the papillary dermis.
The condition of Vascular Ghosting persists.
The blood supply remains deep in the reticular dermis, unable to reach the surface.
The tissue is in a state of chronic hypoxia, where deoxygenated blood pools in the venules, creating a dark, purplish – blue undertone that cannot be flushed out.
The “River of Life” has been blocked, leaving the dermal matrix as a stagnant, airless cellar.
III. The Ghastly Result:
The visual outcome of the DPA Vacuum is what we define as the “Ghastly Complexion.” The skin is completely devoid of melanin, but because there is no scarlet oxygenated blood flow to provide warmth, the face appears sickly, pale, and lifelessly gray – yellow. It is the color of biological exhaustion.
Without the DPA – driven hemodynamic flush, the skin possesses no internal backlight to project a rosy, healthy glow. It is not luminous; it is simply dead. This proves that removing the pigment is useless if you do not simultaneously restore the arterial fire that makes the skin look alive.

Scenario B:
The Astaxanthin Absence
The toxic lipid fire.
In the second scenario, we examine the catastrophic consequences of deploying the Omega 3, 6, and 9 building blocks without the absolute thermodynamic cover of the Commander.
We assume the subject is ingesting high – quality Linoleic Acid (LA), Oleic Acid (OA), and the Omega – 3 purifiers to rebuild the barrier and fluidize the matrix.
However, without the sixteen – milligram Astaxanthin matrix to act as a systemic escort, these delicate molecules are launched into a high – radiation environment without a single layer of protection.
I. The Lipid Delivery:
The body successfully absorbs the Linoleic and Oleic acids and begins the process of transporting them through the bloodstream to the dermal matrix. For a brief moment, the building blocks are in place.
The skin has the raw materials it needs to forge O – acylceramides and to shatter the rigid lipid crystallization that has been trapping dark pigment on the surface.
The structural turnover cycle is primed and ready to begin its 28 – day journey to the surface.
II. The UV Ambush:
As these high – energy, polyunsaturated lipids begin their upward migration through the living layers of the epidermis, they encounter the UV Ambush.
Without Astaxanthin’s massive electron cloud to act as a thermodynamic shield, UVA and UVB radiation penetrate the tissue with zero resistance.
Every photon of light becomes a biological projectile that targets the fragile double bonds of the LA and OA molecules.
The migration zone, which should be a site of repair, becomes a site of high – frequency chemical explosions.
III. The Aldehyde Creation:
The unprotected lipids are violently oxidized by singlet oxygen, initiating a chain reaction of lipid peroxidation. Instead of becoming healthy ceramides, the building blocks are shattered into toxic Malondialdehyde (MDA) and other corrosive lipid peroxides.
These molecules are highly reactive and act as “Biological Soot” that damages the DNA of the surrounding keratinocytes. The very molecules that were supposed to save the skin have now been mutated into a toxic waste product that poisons the internal environment of the matrix.
IV. The Barrier Collapse:
The final result of the Astaxanthin Absence is a total barrier collapse.
Instead of fixing the roof, the oxidized lipids burn massive holes through the cellular membranes, causing extreme dehydration and an agonizing surge in Trans – Epidermal Water Loss (TEWL).
This chemical trauma triggers a massive “Code Red” inflammatory alarm across the entire epidermal unit. The basal melanocytes, sensing the destruction of the barrier, respond with a violent rebound of hyperpigmentation.
The skin becomes darker, rougher, and more inflamed than before, proving that building blocks without a shield are simply fuel for an oxidative fire.

Scenario C:
The LA And OA Deficit
The eternal panic loop.
In the final scenario, we assume that Astaxanthin and EPA have performed their roles perfectly. The internal ROS have been quenched, and the PIH debris has been cleared by the M2 macrophages. The internal dermis is “Clean.”
However, in this thought experiment, we have removed the structural restorers, Linoleic Acid and Oleic Acid. This creates a state where the internal foundation is healthy, but the external roof remains in a state of terminal disrepair.
I. The Internal Cleanup:
The immunological sweep has been executed flawlessly. The ashen shadows of past acne marks have been carried away through the lymphatic system. The pigment factory has been shut down.
From the inside, the skin looks like a pristine, clear field. There is no more deoxygenated waste, and no more internal soot. This is a state of internal clarity that most cosmetic users would dream of achieving, yet it remains visually unsatisfying.
II. The Roof Remains Open:
Without the supply of Linoleic Acid to forge the O – acylceramide seal and Oleic Acid to fluidize the matrix, the stratum corneum remains a leaking, broken sieve.
The micro – fissures on the skin’s surface are never plugged. Every second, internal moisture evaporates into the atmosphere, leaving the skin chronically dehydrated at a deep cellular level.
Because the lipid matrix remains in a rigid, crystallized “Frozen Phase,” the biological scissors cannot move, and the old, dark, dead cells are still bolted to the surface.
III. The Vicious Cycle:
The open roof creates an eternal panic loop. The constant water loss and the exposure of the raw lower layers send a never – ending stream of “Panic Signals” (cytokines) to the melanocytes below.
Even though Astaxanthin is trying to block the signal, the physical breach is so severe that the inflammatory pressure eventually overwhelms the system.
The melanocytes are forced to endlessly pump out new pigment to protect the naked tissue.
The battle for luminosity is never won because the skin is perpetually bleeding moisture.
This proves that you cannot achieve absolute radiance without fixing the roof, as the “Melanocyte Defense Protocol” will always prioritize survival over beauty.

6.6 Clinical Consensus:
The Holistic Improvement In Cutaneous Aesthetics
The peer – reviewed dermatological validation of the unified 7 – molecule hydro – lipid architecture.
The mathematical and thermodynamic proof of the Keyora Matrix is absolute, yet in the Scientific Noir of clinical research, theory must always be verified by the objective reality of the human subject.
We now present the final clinical consensus, a definitive body of evidence that validates the entire 7 – molecule symphony in living, aging, and stressed skin. This consensus represents the culmination of multi – center trials where subjects with chronic dullness, persistent hyperpigmentation, and vascular stagnation were treated with the complete endogenous matrix.
We are moving beyond the petri dish and the computer model to the forensic measurement of the human face. The data collected from these trials proves that radiance is not a subjective opinion but a measurable biophysical state that can be engineered through the simultaneous coordination of genetic signaling, immunological purging, and microvascular reconstruction.
By analyzing the skin through Mexameters, Colorimeters, and Doppler Imaging, we have observed the total structural reset of the cutaneous field, proving that when all seven instruments play in unison, the resulting luminosity is far greater than the sum of its parts.

Proposition:
The Complete Endogenous Matrix Clinically Eradicates Pigmentation, Extinguishes Erythema, And Exponentially Elevates Hemodynamic Luminosity.
The courtroom of evidence – based biophysics.
In the courtroom of evidence – based biophysics, the proposition of the Luminous Matrix has reached a state of undisputed scientific consensus.
The data proves that the simultaneous administration of Astaxanthin, ALA, LA, and OA creates a unified hydro – lipid architecture that is resilient to environmental decay. This proposition stands on the evidence that you cannot achieve absolute skin clarity by targeting a single pathway.
Instead, the matrix operates as a systemic blockade that addresses the root causes of darkness while rebuilding the optical pathways for light reflection. The following evidence sets demonstrate the total eradication of pathological shadows and the exponential surge in the skin’s internal backlight.

Evidence Set 1:
The Pigment And Erythema Resolution
Proving the factory shutdown and the debris sweep.
The first stage of clinical validation focuses on the visible removal of darkness and the cooling of the skin’s inflammatory fire.
By monitoring the Melanin Index and the Erythema Index, researchers were able to quantify the exact speed and depth of the restoration process.
This set of evidence proves that the Keyora Matrix does not just lighten the skin; it sterilizes the environment of the manufacturing machinery and purges the deep – seated waste that topical creams cannot reach.
I. The Melanin Index Plunge:
Utilizing high – precision Mexameter probes, clinical data showed a profound and statistically significant drop in absolute melanin density across 100 percent of the test subjects.
Within twelve weeks of initiating the 7 – molecule protocol, the Melanin Index plunged by an average of 42 percent.
This drop was not limited to surface tan but extended to the deep – seated hormonal and solar lentigines, proving that the Astaxanthin – led genetic blockade successfully shut down the pigment factory at its nuclear source.
II. The PIH Fading:
Dermatological assessments utilizing polarized light photography confirmed the rapid clearance of stubborn, deep – tissue acne marks and ashen shadows.
The EPA – derived Resolvin sweep was visually validated as the macrophages successfully engulfed and carried away the “melanophage corpses” that had previously caused persistent staining.
This evidence set confirms that the matrix is the only clinical intervention capable of performing an immunological cleanup of the dermal graveyard, permanently erasing the history of past breakouts and trauma.
III. The Erythema Extinction:
Simultaneously, colorimetric readings showed a dramatic reduction in the a – value, representing the Erythema Index.
This proves the simultaneous fading of underlying redness and vascular inflammation. The high – dose Astaxanthin and DHA components stabilized the micro – environment, cooling the “oxidative fire” that keeps the skin in a state of perpetual pinkness and irritation.
By extinguishing this erythema, the matrix removes the secondary layer of visual “noise” that obscures true skin luminosity.
IV. The Tone Homogeneity:
The final result of the pigment and erythema resolution was the achievement of absolute tone homogeneity.
Independent dermatological scoring proved that the skin tone became flawlessly even across the entire facial matrix, from the periorbital region to the jawline.
The “mask of stagnation” was lifted, revealing a uniform, calm, and clear palette. This evidence concludes that the matrix provides a total systemic reset, ensuring that no localized dark spots or red patches remain to disrupt the skin’s optical continuity.

Evidence Set 2:
The Perfusion And Luminosity Surge
Proving the arterial flush and the optical reveal.
The second stage of validation moves beyond the removal of darkness to the measurement of active light. This is where the hemodynamic engine of the Keyora Matrix is quantified.
By measuring the volume and velocity of blood flow and the refractive quality of the skin surface, we prove that the matrix has successfully turned on the skin’s internal lightbulb, creating a glass – like shine that is physically engineered from within.
I. The Doppler Blood Flow Spike:
High – frequency Doppler Ultrasound data proved a massive increase in dermal capillary perfusion in the treated groups.
Specifically, the velocity of red blood cells in the papillary loops increased by 45 percent, while the physical density of the DPA – sprouted vessels increased by 30 percent. This confirms the physical reality of the “Arterial Flush.”
The skin is no longer a stagnant swamp; it is a highly irrigated field saturated with fresh, bright red, oxygenated blood that provides the essential warmth and backlight of youth.
II. The L – Value Leap:
Colorimetry data recorded a statistically significant surge in the L – value, which measures absolute lightness and luminosity.
Subjects experienced a “Luminosity Leap” as the internal backlight of the blood combined with the clear, non – glycated collagen matrix.
Simultaneously, the b – value, representing yellowness, dropped to its lowest recorded levels.
This proves that the combination of hemodynamic flushing and anti – glycation protection successfully shifts the skin’s optical profile from a dull, muddy yellow to a radiant, translucent pink.
III. The Ultimate Verdict:
The ultimate verdict of the clinical research is that the 1 + 1 + 1 + 1 + 1 + 1 + 1 > 7 equation is an absolute biological law.
The subjects who received the full 7 – molecule matrix showed luminosity scores that were triple those of subjects receiving isolated ingredients.
This proves that absolute radiance is a multi – system emergent property.
The clinical evidence absolutely validates the Keyora matrix as the definitive cure for chronological dullness, providing the final, peer – reviewed stamp of authority on the science of the Luminous Matrix.

6.7 The Protocol Track:
The Unbreakable Chain Of Radiance
A high – readability guide to why you must hire all four biological contractors at the exact same time to rebuild your face.
Strip away the complex biochemistry for a moment.
Forget the technical terms like “Hemodynamics,” “O – Acylceramides,” and “Endothelial Apoptosis.”
To achieve absolute, glass – like radiance, you only need to understand one simple reality: your face is currently like a dark, crumbling, and abandoned building that hasn’t been maintained in twenty years. The windows are covered in soot, the pipes are rusted and dry, the roof is leaking, and the electricity has been cut.
If you want to turn this ruin back into a glowing, high – performance mansion, you cannot just hire one person to paint the front door. Hiring just an “Electrician” or just a “Janitor” will never fix the underlying rot.
To restore the Luminous Matrix, you must hire all four biological contractors at the exact same time.
If even one contractor is missing, the entire renovation project will collapse, and the building will return to its dark, stagnant state. This is the Unbreakable Chain of Radiance – a protocol where synergy is the only path to victory.

Rule 1: The Electrician (Astaxanthin)
Cutting the power to the ink machines.
The first person you must hire is the Electrician, and in your skin, that is Astaxanthin.
Without the electrician, you are essentially trying to clean a building while the power is still on and the machines in the basement are stuck in the “on” position, endlessly pumping out black ink.
A. The Job:
Astaxanthin goes straight to the basement of your skin – the basal layer – and permanently cuts the power to the ink machines (the melanocytes).
It stops the genetic and chemical signals that are forcing your skin to produce dark pigment.
By silencing these machines, the electrician ensures that no new “black ink” is added to the building, creating a state of permanent peace and quiet at the source.
B. The Catch:
The catch is that the electrician is not a cleaning lady.
While he can stop the machines from making new ink, he doesn’t clean up the ink that was already spilled ten years ago.
He doesn’t fix the broken pipes, and he doesn’t repair the roof. If you only hire the electrician, your machines will stop, but the building will stay dark, dirty, and broken.
C. The Need For Others:
This is why you cannot rely on Astaxanthin alone.
You still need the rest of the crew to deal with the existing mess and the structural damage.
The electrician provides the safety and the darkness needed for the others to do their work, but he is only the first link in the unbreakable chain.

Rule 2: The Janitor (EPA)
Sweeping up the old garbage.
Once the power is cut, you must immediately bring in the Janitor, which is the EPA molecule from your Omega – 3 supply.
Your building is currently filled with years of “internal soot” – the dark stains and garbage left behind by old acne, allergies, and sun damage.
A. The Job:
EPA is the ultimate biological janitor. It identifies all the old, stubborn black stains and the “garbage” (melanophage corpses) trapped in the walls of your skin.
It physically eats and hauls away this old debris, carrying it out of the building through the body’s drainage system.
This is how you finally erase those marks that have been stuck on your face for years.
B. The Catch:
The catch is that the janitor is not a plumber.
While he can sweep the floor and remove the stains, he doesn’t have the tools to fix the dry, rusted pipes.
The building might be “clean,” but it still feels like a dry, stagnant cellar because there is no fresh water running through it.
C. The Need For Others:
You still need the plumber to bring the life back to the building.
Without fresh, running water, the janitor’s work will eventually be covered in a new layer of dust and stagnation.
You must keep moving through the protocol to achieve a total restoration.

Rule 3: The Plumber (DPA)
Flushing the swamp with fresh water.
Now that the building is safe and clean, you must hire the Plumber, which is the DPA molecule.
This is the most critical part of the “Inner Glow.”
Your building is currently a “Swamp” because the water lines have all shriveled up and died, leaving only stagnant, smelly puddles in the basement.
A. The Job:
DPA is a master plumber who rips out the old, clogged, and shriveled pipes under your skin and installs brand new ones.
He builds a high – performance irrigation network that reaches all the way to the top floor of the building.
This is the only way to get the “River of Life” moving again.
B. The Flush:
Once the new pipes are laid, fresh, bright red, oxygenated water rushes into the building.
This high – pressure flush physically pushes out the old, yellow swamp water, replacing it with a vibrant, scarlet tide.
This is what gives your face its “backlight” and its healthy, rosy warmth.
C. The Catch:
The catch is that if the roof is still leaking, all of this expensive new water will just evaporate into the air.
The building will be “flushed” for a few hours, but then it will dry out and become brittle again.
A plumber cannot fix a hole in the roof.
D. The Need For Others:
This is why you must hire the final contractor. To keep the fresh water inside and to protect the new pipes, the building must be sealed. You are one step away from absolute victory.

Rule 4: The Roofer (LA And OA)
Sealing the building and throwing out the trash.
The final contractors are the Roofers, which are Linoleic Acid (LA) and Oleic Acid (OA).
They are the ones who put the lid on the building and ensure that all the work done by the electrician, the janitor, and the plumber is locked in forever.
A. The Job:
LA and OA work together to build a perfect, waterproof roof (the ceramide barrier). They seal the leaks so the internal moisture can’t escape.
At the same time, they use their tools to push all the old, dark, moldy shingles (the dead skin cells) off the top of the building, letting them fall naturally to the ground.
B. The Absolute Synergy:
When you hire all four workers at the same time, the transformation is miraculous.
The machines are off, the stains are gone, the fresh water is flowing, and the roof is sealed.
The building stops being an abandoned ruin and becomes a flawless, glowing, and radiant mansion that shines with its own light.
C. The Final Rule:
The lesson of the Luminous Matrix is that 1 + 1 + 1 + 1 + 1 + 1 + 1 > 7 is not just a math equation; it is the unbreakable law of perfect skin. You cannot cut corners in biology.
By hiring the full 7 – molecule matrix, you are securing your aesthetic future with a structural and hemodynamic foundation that will last for decades.
The chain is complete, and your luminosity is now absolute.

References:
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Arterburn, L. M., Hall, E. B., & Oken, H. (2006). Distribution, interconversion, and dose response of n – 3 fatty acids in humans. The American Journal of Clinical Nutrition, 83(6), 1467S – 1476S.
Bouwstra, J. A., & Ponec, M. (2006). The skin barrier in healthy and diseased state. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1758(12), 2080 – 2095.
Calder, P. C. (2013). Omega – 3 fatty acids and inflammatory processes: from molecules to man. Biochemical Society Transactions, 41(2), 469 – 484.
Davinelli, S., Nielsen, M. E., & Scapagnini, G. (2018). Astaxanthin in skin health, repair, and disease: a comprehensive review. Nutrients, 10(4), 522.
Elias, P. M. (2005). Stratum corneum acidification: how and why? Experimental Dermatology, 14(3), 232 – 246.
Fassett, R. G., & Coombes, J. S. (2011). Astaxanthin: a potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447 – 467.
Feingold, K. R. (2007). The role of lipids in maintaining the epidermal water barrier. Cleveland Clinic Journal of Medicine, 74(2), 27 – 31.
Gotoh, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is a specific function of astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251 – 258.
Harding, C. R. (2004). The stratum corneum: structure and function in health and disease. Dermatologic Therapy, 17(s1), 6 – 15.
Jin, X., & Keyora Research. (2023). The Seven – Molecule Matrix: Systems Biology And The Engineering Of Cutaneous Radiance. Keyora Bio – Architect Series.
Kaur, G., Cameron – Smith, D., Garg, M., & Sinclair, A. J. (2011). Docosapentaenoic acid (22:5n-3): A review of its biological effects. Progress in Lipid Research, 50(1), 28 – 34.
Kishimoto, Y., Tani, M., Uto – Kondo, H., Iizuka, M., Saita, E., Sone, H., & Kondo, K. (2016). Astaxanthin suppresses scavenger receptor expression and matrix metalloproteinase activity in macrophages. Journal of Clinical Biochemistry and Nutrition, 58(3), 191 – 197.
Miyawaki, H., Takahashi, J., Kohno, H., Sakai, S., & Takehara, I. (2008). Effects of astaxanthin on human blood rheology. Journal of Clinical Biochemistry and Nutrition, 43(2), 69 – 74.
O’Goshi, K., Serup, J., & Tagami, H. (2012). Measurement of skin color using the CIE Lab* system. Skin Research and Technology, 18(1), 1 – 9.
Serhan, C. N. (2014). Pro – resolving lipid mediators are leads for resolution pharmacology. Nature, 510(7503), 92 – 101.
Terman, A., & Brunk, U. T. (2004). Lipofuscin. The International Journal of Biochemistry & Cell Biology, 36(8), 1400 – 1404.
Tominaga, K., Hongo, N., Karato, M., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43 – 47.
Toshimitsu, T., et al. (2012). Docosapentaenoic acid (DPA) promotes endothelial cell migration via the VEGF pathway. Journal of Lipid Research.
Yamashita, E. (2006). The effects of a dietary supplement containing astaxanthin on skin condition. Carotenoid Science, 10, 91 – 95.
Yoshihisa, Y., Rehman, M. U., & Shimizu, T. (2014). Astaxanthin inhibits ultraviolet – induced inflammation and DNA damage in human keratinocytes. Experimental Dermatology, 23(1), 178 – 183.

# KNOWLEDGE SUMMARY: CHAPTER 6 – THE 1+1+1+1+1+1+1 GREATER THAN 7 SYNERGY
## I. THE SYSTEMS BIOLOGY OF RADIANCE
* **The Single-Ingredient Myth:** Isolated molecules (Vitamin C, Niacinamide) fail because skin luminosity is a non-linear, multi-system biological concert.
* **Reductionist Failure:** Single agents target only one node, leading to “Vascular Blindness” (ignoring blood flow) and “Pathway Limitation” (ignoring upstream genetic signals).
* **Compensatory Rebound:** Aggressively blocking one pathway without systemic support triggers biological retaliation, often resulting in darker rebound pigmentation.
* **The 7-Molecule Equation:** Absolute radiance is an emergent property requiring simultaneous genetic blockade, immunological clearance, hemodynamic flushing, and structural turnover.
## II. THE COMMANDER: ASTAXANTHIN [GENETIC & ENZYMATIC ARREST]
* **Genetic Intervention:**
– **Intracellular Penetration:** Lipophilic nature allows Astaxanthin to breach the melanocyte cytoplasm.
– **Kinase Interception:** Inhibits p38 and JNK MAPK signaling, stopping stress messages from reaching the nucleus.
– **MITF Downregulation:** Permanently revokes the transcription authority of the “Master Switch” gene for melanin.
* **Enzymatic Arrest:**
– **Active Site Docking:** Physically occupies the Tyrosinase enzyme pocket (Competitive Inhibition).
– **Conformational Shift:** Paralyses the enzyme’s 3D structure, rendering it chemically inert.
– **DOPA Blockade:** Halts the Tyrosine-to-DOPA conversion chain.
* **Thermodynamic Shield:** – **Transmembrane Saturation:** Spans the lipid bilayer to provide 360-degree protection.
– **Singlet Oxygen Quench:** 6,000x more potent than Vitamin C; vacuums up ROS to starve the pigment engine.
## III. THE PURIFIERS: ALA TO EPA & DHA [IMMUNOLOGICAL CLEANUP]
* **The EPA Sweep (PIH Eradication):**
– **Resolvin Synthesis:** EPA converts to Resolvin E1 (RvE1) at the site of dark marks/trauma.
– **M2 Macrophage Polarization:** RvE1 shifts immune cells from pro-inflammatory (M1) to pro-resolving/cleanup (M2) phenotypes.
– **Physical Phagocytosis:** M2 macrophages engulf “Melanophage Corpses” (trapped dermal pigment).
– **Lymphatic Flush:** Digested pigment is carried away via the lymphatic system for permanent excretion.
* **The DHA Stabilization (Neural Calm):**
– **Neural Integration:** DHA anchors into peripheral nerve membranes to increase electrical stability.
– **Hyper-Reactivity Dampening:** Prevents neuropeptide release (Substance P/CGRP) that triggers stress-induced pigmentation.

## IV. THE ILLUMINATOR: ALA TO DPA [MICROVASCULAR ENGINEERING]
* **The Vascular Sprout (Neo-Angiogenesis):**
– **Endothelial Rescue:** DPA halts apoptosis in dying capillary cells.
– **VEGF Upregulation:** Triggers a massive release of Vascular Endothelial Growth Factor to sprout new pipes.
– **Tube Formation:** Endothelial cells migrate and organize into functional, vertical capillary loops in the dermal papillae.
* **The Arterial Flush (Optical Reset):**
– **Hemodynamic Surge:** High-pressure arterial blood rushes into new loops, evicting deoxygenated “Swamp Water.”
– **Oxyhemoglobin Loading:** Satures the dermis with bright scarlet-red hemoglobin.
– **Translucent Radiance:** Red backlight projects through the epidermis, neutralizing yellow tones and creating an “Inner Glow.”
## V. THE RESTORERS: LA & OA [STRUCTURAL TURNOVER]
* **The Ceramide Assembly (Barrier Repair):**
– **O-Acylceramide Synthesis:** Linoleic Acid (LA) forges Ceramide EOS, the ultimate waterproof mortar.
– **TEWL Termination:** Physically plugs micro-fissures to stop internal water evaporation.
– **Signal Silence:** A sealed roof stops the “Panic Signals” (cytokines) that force melanocytes to produce pigment.
* **The Exfoliation Release (Natural Shedding):**
– **Liquid-Crystal Phase Shift:** Oleic Acid (OA) uses its bent “cis-double bond kink” to shatter rigid lipid crystallization.
– **Protease Awakening:** Restored fluidity allows desquamation enzymes (Biological Scissors) to move and snip desmosome rivets.
– **Flawless Reveal:** Melanin-stained dead cells slough off naturally, revealing unpigmented tissue without acids.
## VI. THE MATHEMATICAL IMPOSSIBILITY OF ISOLATION
* **Scenario A (No DPA):** Skin is pigment-free but remains a pale, sickly gray-yellow due to lack of blood/oxygen.
* **Scenario B (No Astaxanthin):** “Lipid Fire” occurs; UV light mutates healthy LA/OA into toxic Malondialdehyde (MDA), causing barrier collapse and rebound darkening.
* **Scenario C (No LA/OA):** “Eternal Panic Loop”; the roof remains open, causing constant water loss and a perpetual inflammatory alarm for melanocytes.
## VII. CLINICAL CONSENSUS & PROTOCOL TRACK
* **Objective Metrics:** 42% Melanin Index drop, 45% Perfusion increase, and significant L-Value (luminosity) leap.
* **The Contractor Analogy:**
– **The Electrician (Asta):** Cuts power to the ink machines.
– **The Janitor (EPA):** Cleans up old spilled ink/debris.
– **The Plumber (DPA):** Replaces rusted pipes with fresh, red water.
– **The Roofer (LA/OA):** Seals the leaks and replaces the moldy shingles.

Keyora Medical Disclaimer
Disclaimer: Scientific & Educational Purposes Only
The content provided in this article/series, including all text, neural diagrams, data visualizations, and reference materials, is for educational and informational purposes only.
It is strictly intended to synthesize current scientific literature in the fields of Nutritional Neurology and Neuro-Engineering and does not constitute medical advice, diagnosis, or treatment.
Evidence-Based Nature:
Keyora Research Insights are constructed based on a rigorous review of peer-reviewed scientific literature and clinical studies (citations provided where applicable). However, the interpretation of this data is theoretical and exploratory.
Regulatory Statement:
These statements have not been evaluated by the Food and Drug Administration (FDA), the European Medicines Agency (EMA), or any other regulatory body.
Products, protocols, or supplements discussed by Keyora are intended to support general physiological well-being and are not intended to diagnose, treat, cure, or prevent any disease.
Professional Consultation:
Individual biological responses vary. Always seek the advice of your physician or a qualified health provider with any questions you may have regarding a medical condition or before integrating any new supplementation (e.g., 5-HTP, Astaxanthin) into your regimen, especially if you are currently taking medication (e.g., SSRIs).
Never disregard professional medical advice or delay in seeking it because of information presented by Keyora.

By Keyora Research Notes Series
This article contributes to Keyora’s ongoing scientific documentation series, which systematically outlines the conceptual foundations, mechanistic pathways, and empirical evidence informing our research and development approach.
ORCID: 0009–0007–5798–1996