Keyora Astaxanthin EP-13: The Endogenous Sunshield: Intercepting the Photo-Oxidative Strike
By Keyora Research Notes Series
This article contributes to Keyora’s ongoing scientific documentation series, which systematically outlines the conceptual foundations, mechanistic pathways, and empirical evidence informing our research and development approach.
ORCID: 0009–0007–5798–1996

THE ILLUSION OF THE SURFACE
You meticulously execute your defensive protocol every single morning – slathering your face in thick – chalky layers of premium SPF 50 before you ever step foot into the daylight.
You consciously wear wide – brimmed hats to physically block the solar assault and diligently avoid direct sunlight during peak ultraviolet hours.
Yet – as you catch a glance in the rearview mirror years later – you still see the inevitable micro – fractures of fine lines spidering across the corners of your eyes – the subtle but undeniable structural sagging at the jawline – and the deep – persistent clouds of hyperpigmentation rising to the surface.
Your discipline was absolutely perfect – your adherence to the modern dermatological doctrine was flawless – but your biological architecture still collapsed.
The painful – undeniable truth is that the external perimeter failed you.

PROPOSITION A:
WHY DOES TOPICAL SPF MATHEMATICALLY FAIL TO STOP PHOTO-AGING?
The modern skincare industry has built a multi – billion – dollar empire by selling the consumer a highly comforting but mathematically flawed statistical illusion.
They have convinced you that blocking ultraviolet radiation at the dead stratum corneum is the absolute zenith of cellular defense.
In the rigorous – thermodynamic reality of the Bio – Architect – relying solely on a superficial topical cream to stop a high – velocity photon strike is an architectural absurdity.

The Pivot:
The 1 Percent Breach
We must ruthlessly deconstruct the actual physics of chemical and physical sunscreens to understand exactly why your skin is still aging despite your flawless discipline.
Topical SPF is not an impenetrable – magical forcefield; it is merely a chemical probability filter smeared across the dead exterior surface of the epidermis.
Even if an elite – laboratory – grade SPF lotion successfully reflects or chemically absorbs ninety – nine percent of the incoming solar ultraviolet radiation – it is an absolute – mathematical certainty that one percent of those high – energy photons will breach the chemical wall.
Over the course of thousands of hours of ambient exposure – driving in your car – walking between office buildings – or sitting near a heavily tinted window – this fractional breach accumulates into a massive – devastating kinetic payload that completely bypasses your topical security.

Detail A (The Dermal Strike):
You must understand the precise ballistics of this solar radiation to grasp the magnitude of the threat.
The longer – wavelength Ultraviolet A (UVA) photons – unlike their shorter UVB counterparts – possess the exact frequency and penetrative capacity required to completely bypass the superficial epidermal surface without triggering an immediate – painful sunburn alarm.
They pass silently and invisibly through the dead skin cells – behaving like microscopic thermodynamic missiles.
They smash directly into the deep dermal layer – violently striking the delicate fibroblasts – shattering the structural collagen scaffolding – and fundamentally destabilizing the highly vulnerable polyunsaturated structural lipids that hold your extracellular matrix together in a state of youthful tension.

Detail B (The Internal Fire):
The moment that one percent payload breaches the perimeter and impacts the deep dermal tissue – it immediately triggers a catastrophic singlet oxygen explosion inside the delicate cellular lipid bilayer.
This violent thermodynamic event instantly initiates a self – propagating chain reaction of lipid peroxidation – melting the cellular walls – mutating the mitochondrial DNA – and severing the elastin fibers.
This singlet oxygen explosion directly upregulates matrix metalloproteinases – specifically collagenase and elastase – which act as biological scissors that aggressively snip away the youthful tension of your skin.
Here is the brutal – unvarnished truth of the matter: obsessively applying topical SPF to prevent the deep structural aging of your face is the exact equivalent of applying a fresh coat of beautiful white paint to the exterior of your house while the living room inside is actively burning to the ground from an uncontrollable electrical fire.
The surface looks temporarily protected – but the foundation is quietly turning to ash.

The Epiphany:
The Endogenous Sunshield
To survive the relentless solar siege and permanently halt the degradation of your biological interface – we must completely abandon the illusion of the surface and embrace the rigorous engineering paradigm of the Keyora Bio – Architect.
True photo – protection is absolutely not about attempting the impossible task of “blocking” every single particle of light at the dead epidermal surface.
True – sovereign biological defense is entirely about “dissipating” that lethal thermodynamic energy from the inside out – directly within the living cellular membrane where the structural damage actually occurs and where the aging process is physically born.

Detail A (The 7-Part Matrix):
We do not formulate superficial lotions or temporary topical patches that wash away in the rain; we deploy an elite – 7 – part endogenous lipidomic matrix directly into the systemic circulation to fortify the skin from the blood supply upwards.
We deploy Natural Astaxanthin to act as the ultimate [Thermodynamic Heat – Sink] – a precise 30 – Angstrom transmembrane shield that embeds directly into the dermal fibroblasts to absorb the kinetic shockwave of the UV strike into its conjugated pi – electron cloud.
This commander is perfectly anchored and supported by a precise Bioactive Carrier of cold – pressed Flaxseed Oil that safely smuggles the parent source codes of Alpha – Linolenic Acid (ALA) – Linoleic Acid (LA) – and Oleic Acid (OA) past the destructive gastric acids.
Once safely inside the hepatic vault of the liver – these parent lipids unleash the endogenous synthesis of pure Docosahexaenoic Acid (DHA) – Eicosapentaenoic Acid (EPA) – and Docosapentaenoic Acid (DPA) from within – flooding the dermal capillaries with specialized repair operatives.
These synthesized quantum bricks and chemical sterilizers do not sit passively on the surface; they are pumped through the cardiovascular network directly into the deepest layers of the hypodermis and dermis – building an indestructible wall of lipids.

Detail B (The 1+1+1+1+1+1+1 > 7 Synergy):
This entire bio – architectural system operates on the absolute – non – negotiable mathematical law of 1+1+1+1+1+1+1 > 7.
-
The Astaxanthin commander cannot successfully hold the defensive line against the solar fire without the LA – derived ceramide mortar to seal the trans – epidermal water loss and prevent the tissue from desiccating.
-
It cannot function without the OA actively maintaining the fluid liquid – crystal state of the membrane and triggering the AMPK metabolic reset to clear the damaged cellular debris.
-
It absolutely requires the ALA – derived DHA to provide the heavy – unoxidized structural base for new cell generation.
-
It requires the DPA to execute a rapid vascular rescue of the choked dermal capillaries – ensuring optimal nutrient delivery.
-
It demands the EPA to act as the localized fire extinguishers – deploying E – series Resolvins to chemically sterilize the UV – induced neuro – inflammation.
-
Furthermore – it utilizes the LA to synthesize the Arachidonic Acid (AA) signal – ensuring rapid cellular communication to coordinate these repair protocols.
They must operate simultaneously in a flawless – closed – loop synergy to guarantee survival.

The Trust Algorithm:
It is time to reactivate your highly calibrated cognitive filter and execute the Trust Algorithm with absolute prejudice.
You must immediately and permanently reject the reductionist marketing claims of superficial “surface brightening” serums or mathematically impossible “magic anti – aging creams” that completely lack any endogenous penetrative capacity.
You are not a passive consumer buying hope in a jar; you are an architect securing a biological asset.
You must demand uncompromising – systemic thermodynamic cellular defense.
In the upcoming chapters – we are about to execute a ruthless – microscopic forensic audit of a direct ultraviolet photon strike upon the human dermis.
We will deconstruct the exact thermodynamics of the dermal collapse – trace the molecular pathways of the singlet oxygen explosion – and mathematically prove exactly how this 7 – part endogenously synthesized matrix intercepts the radiation – repairs the structural scaffolding – and permanently secures your macroscopic biological sovereignty.
The era of painting over the fire is over.
The era of endogenous bio – engineering has begun.

Chapter 1: The Forensic Audit of a UV Strike:
Physics and Thermodynamics of Dermal Degradation
Deconstructing the physical penetration of UVA radiation and the thermodynamic generation of singlet oxygen within the dermal architecture.
In the rigorous discipline of forensic pathology – the most devastating and fatal wounds frequently present with absolutely no immediate external bleeding or visible surface swelling.
The modern skincare consumer has been heavily conditioned to obsess over the immediate – painful sensation of a sunburn – operating under the dangerous and fundamentally false biological assumption that if their skin is not glowing red – they are completely safe from solar degradation.
We are now going to physically peel back the dead – superficial layer of the epidermis and descend directly into the deep – structural load – bearing wall of your biological interface: the dermis.
We will meticulously track that fractional one percent of high – energy ultraviolet photons that inevitably leak past your daily SPF application.
We are going to watch the invisible – silent thermodynamic storm they unleash upon the actual living architecture of your youth.

I. THE ILLUSION OF ERYTHEMA
Why the absence of a sunburn does not equal cellular safety.
The consumer cosmetic and symptom industry has spent decades executing a highly effective – yet biologically incomplete – psychological conditioning campaign that trains the general public to singularly fear the short – wave UVB radiation – the exact wavelength responsible for the acute – visible surface alarm known clinically as erythema or the common sunburn.
Because UVB radiation possesses a much shorter wavelength and a highly concentrated energy signature – it violently impacts the very top layer of the skin – immediately triggering a loud – painful inflammatory cascade of prostaglandins that alerts the biological host to the immediate damage.
However – by hyper – focusing the entire consumer market on this loud – superficial alarm system and heavily marketing SPF numbers that primarily measure UVB defense – the industry has completely ignored the far more insidious – silent – and structurally devastating penetration of the longer – wave UVA radiation.
UVA radiation makes up roughly ninety – five percent of the ultraviolet light reaching the surface of the Earth – and it does not trigger the immediate erythema response.
It does not cause your skin to turn red – it does not blister the surface – and it does not signal immediate pain to your central nervous system. Instead – it passes completely undetected through the epidermal fortress like a thermodynamic ghost – penetrating deeply into the living tissue where your nerve endings cannot adequately sense the thermal transfer.
True – structural photo – aging – the kind that mathematically manifests as deep – uncorrectable wrinkles – profound jowl sagging – and the permanent loss of structural elasticity – is absolutely not a superficial surface burn that fades after a few days.
It is a highly violent – microscopic – inside – out collapse of your foundational cellular scaffolding that occurs entirely without a biological warning signal. When you rely solely on the absence of a superficial sunburn to measure your biological safety – you are operating under a fatal delusion.
You are functionally blind to the silent – continuous assassination occurring just millimeters beneath your visible surface.

II. DESCENDING INTO THE EPICENTER
Establishing the dermal matrix as the primary crime scene.
To truly understand the absolute thermodynamic mechanics of this silent assassination – we must firmly establish the dermal matrix as our primary forensic crime scene and ruthlessly profile its highly vulnerable inhabitants.
Beneath the dead – hardened keratinocytes of the stratum corneum lies the dermis – a highly vascularized – dynamically alive microscopic ecosystem that functions as the absolute structural load – bearing foundation of your biological interface.
This is the exclusive domain of the fibroblasts – the specialized biological architects and master weavers responsible for continuously spinning the dense – interlocking networks of Type I and Type III collagen – as well as the highly elastic elastin fibers that keep your skin tightly bound and perfectly tensioned over the underlying musculature.
These critical structural proteins are suspended in a gel – like ground substance rich in hyaluronic acid – creating a perfectly hydrated – pressurized environment. But more importantly – surrounding and protecting these critical cellular factories and their sensitive mitochondrial DNA are the extremely delicate – polyunsaturated lipid membranes.
These phospholipid bilayers are the ultimate gatekeepers – regulating nutrient flow – maintaining the fluid liquid – crystal state of the cellular environment – and facilitating complex enzymatic reactions. Because they are packed with fragile carbon – carbon double bonds – they are thermodynamically vulnerable to kinetic strikes.
This deep – hidden matrix is the exact location where the true assassination of youth occurs. It is not a gradual – gentle fading of cellular vitality brought on by the mere passage of time; it is a highly violent – kinetically charged sub – atomic warzone where high – velocity solar projectiles continuously collide with fragile lipid architecture.
We are now going to isolate the exact biological moment of impact.
We will slow down the molecular clock to a fraction of a nanosecond – freeze the cellular environment – and prepare to rigorously witness the exact – terrifying quantum physics of a single ultraviolet photon strike as it detonates within the living engine of your dermal grid.

1.1 THE KINETIC PENETRATION:
UVA VERSUS UVB
Why surface-level erythema is a distraction from the silent, deep-tissue assassination of the dermal matrix.
To catch a biological killer operating within the microscopic – highly complex theater of the human skin – you must first rigorously and clinically understand the exact physical ballistics of their chosen weapon.
In the exacting study of structural photo – aging – we are not tracking a slow – moving chemical poison or a replicating bacterial pathogen; we are tracking pure – massless thermodynamic energy traveling at the speed of light.
This energy arrives from the solar atmosphere in a continuous – unfathomable bombardment of discrete sub – atomic packets known as photons – and the exact trajectory – depth – and manifestation of their destruction is entirely dictated by their specific electromagnetic wavelength.
Just as a seasoned forensic pathologist can instantly distinguish between the loud – superficial blunt force trauma of a heavy club and the clean – deep – lethal penetration of a high – velocity sniper round simply by examining the entry wound – the Bio – Architect must rigorously differentiate between the superficial radiation that causes a temporary burn and the deeply penetrating radiation that permanently severs your cellular architecture.
The consumer market has been dangerously trained to fight the wrong war.
You cannot possibly secure your systemic biological interface if you are actively confusing the loud – painful – superficial decoy with the silent – thermodynamic assassin.

1.1.1 THE UVB DECOY
The high-energy, short-wave radiation that triggers the surface alarm.
The entire commercial sunburn industry is almost exclusively built around mitigating the effects of Ultraviolet B radiation – a highly energetic band of light occupying the electromagnetic spectrum precisely between 280 and 315 nanometers.
Because UVB photons possess a significantly shorter wavelength and a correspondingly higher frequency – they carry a massive – violent kinetic payload that makes them extremely biologically reactive.
However – this exact physical characteristic is also their primary limitation regarding deep tissue penetration.
When a barrage of UVB photons strikes the human biological perimeter – the vast majority of their kinetic energy is instantaneously absorbed by the very first layers of defense they encounter: the dead keratinocytes of the stratum corneum and the living cells of the superficial epidermis.
They physically cannot penetrate deeply into the dermal layers because they violently dump their thermodynamic energy the moment they hit the surface.
This immediate – superficial energy dump causes massive – acute trauma to the epidermal DNA. The localized cells recognize this catastrophic radiation strike and immediately initiate a desperate biological alarm sequence.
They activate the p53 tumor suppressor protein – trigger widespread cellular apoptosis – and release a massive flood of highly inflammatory chemical messengers – specifically prostaglandins and cytokines.
This localized inflammatory storm causes the surrounding superficial blood vessels to rapidly dilate – flooding the upper skin with blood.
This is the exact physiological mechanism of erythema – the classic – glowing red sunburn.
Furthermore – this inflammation aggressively stimulates the localized nociceptors – the specialized nerve endings that send immediate signals of acute pain and heat directly to your central nervous system. UVB is loud. It screams. It causes immediate discomfort – and it forces you to physically seek shelter.
But in the cold – forensic calculus of the Bio – Architect – this painful sunburn is nothing more than a highly effective biological decoy.
By causing immediate surface pain – UVB radiation keeps the uneducated consumer entirely fixated on the superficial epidermis – completely distracting them from the far more devastating – silent structural collapse occurring just millimeters below the burn.

1.1.2 THE UVA ASSASSIN
The long-wave kinetic projectile that breaches the epidermal fortress.
While the consumer is distracted by the red – burning alarm of the UVB decoy – the true architect of systemic aging slips completely unnoticed past the external perimeter.
Ultraviolet A radiation occupies the longer bandwidth of the spectrum – ranging from 315 to exactly 400 nanometers. This longer wavelength fundamentally alters the physics of its interaction with biological matter.
Unlike the high – frequency UVB that detonates on the surface – the longer – wave UVA photon acts as a high – velocity – deeply penetrating kinetic projectile.
It possesses the exact electromagnetic geometry required to effortlessly bypass the dead keratinized shields of the stratum corneum and completely ignore the superficial epidermis.
Furthermore – its penetrative power is so absolute that it is entirely unimpeded by heavy atmospheric cloud cover or the thick – laminated glass of your office windows and car windshields.
The true horror of the UVA assassin lies in its absolute biological silence. Because these photons bypass the superficial layers – they do not immediately damage the epidermal keratinocytes.
Therefore – they do not trigger the massive prostaglandin inflammatory cascade – and they do not stimulate the superficial pain nociceptors.
You can sit beside a bright – sunlit office window for eight consecutive hours – absorbing billions of UVA photons deeply into your biological architecture – without ever feeling a single degree of heat or the slightest sensation of a sunburn.
But beneath this painless illusion – the UVA photons are carrying their thermodynamic payload directly into the deep dermal matrix.
They are violently striking the delicate – living fibroblasts – the master cellular factories responsible for maintaining your youth.
They are silently bypassing the alarm systems and directly attacking the heavily unsaturated lipid membranes that surround these critical cells – delivering the kinetic energy required to initiate a structural collapse from the inside out.
You do not feel the UVA strike – but twenty years later – you will mathematically see the permanent – sagging wreckage it leaves behind in the dermal grid.

1.1.3 THE INEVITABLE BREACH
The mathematical certainty of SPF failure.
The symptom industry attempts to counter this dual – wave solar assault by selling the consumer topical chemical and physical lotions – heavily promoting broad – spectrum SPF 50 as an impenetrable biological shield.
We must now ruthlessly deconstruct this mathematical impossibility and expose the absolute certainty of topical failure.
The Sun Protection Factor rating system is a highly controlled – laboratory – derived metric that assumes an impossibly thick – perfectly even – undisturbed layer of lotion applied to a static biological surface.
In the kinetic – chaotic reality of the human environment – this flawless application is a thermodynamic fantasy.
The moment you apply a commercial SPF lotion – the chemical shield begins to physically and molecularly degrade. Microscopic fissures instantly form in the barrier as your facial muscles contract and expand during normal communication.
The natural excretion of sebum and sweat physically lifts the chemical filters away from the epidermal surface – creating massive microscopic canyons where radiation can pour through unimpeded.
Furthermore – the very chemical filters designed to absorb the radiation – such as avobenzone – are highly photounstable. As they absorb the incoming UV photons – they undergo rapid structural degradation – literally breaking down and losing their protective capacity under the thermodynamic load of the sun.
Even if we grant the impossible assumption that a consumer applies a perfect – unbroken – re – applied shield that successfully blocks ninety – nine percent of the incoming radiation – the absolute scale of solar bombardment renders the defense inadequate.
When you are exposed to trillions of incoming photons – a one percent leakage rate means that billions of high – energy UVA projectiles are successfully breaching the perimeter every single hour.
And here is the ultimate – undeniable forensic truth: the very microsecond a single UVA photon successfully crosses the 0.1 millimeter threshold of the epidermal barrier – your expensive – surface – level lotion is instantly and completely rendered biologically useless.
You cannot block a projectile with a shield that it has already passed through. Once the photon enters the deep dermis – it is operating entirely outside the jurisdiction of your topical cream.
If you do not possess an active – endogenous thermodynamic interceptor anchored deeply within the living cellular membranes of your fibroblasts – that photon will inevitably detonate its payload – and your biological architecture will suffer a permanent – irreversible structural fracture.
The surface shield has failed; the war must now be fought on the inside.

1.2 THE THERMODYNAMIC WEAPON:
THE SINGLET OXYGEN EXPLOSION
The exact quantum mechanics of how a UV photon weaponizes ground-state oxygen into a hyper-reactive explosive.
As a Bio-Architect analyzing the structural collapse of human tissue, you must eventually confront a profound and highly counterintuitive physical paradox: a photon of ultraviolet light possesses absolutely zero physical mass.
How, then, does a massless, invisible entity physically destroy the dense, structural load-bearing walls of a living human cell?
How does sunlight physically cut through thick, heavily woven bands of dermal collagen and violently shatter the biological lipid membranes that hold your face in a state of youthful tension?
The uncompromising, scientific answer lies entirely in the quantum transfer of pure thermodynamic energy.
We are not dealing with a mechanical blunt force instrument like a surgical scalpel or a physical needle; we are dealing with high-velocity sub-atomic radiation that is capable of hijacking your own internal cellular environment and weaponizing it against you.
To understand the true, hidden mechanics of photo-aging, we must aggressively zoom in to the atomic level and execute a flawless, slow-motion forensic analysis of the exact microsecond a UVA photon successfully breaches your SPF and detonates within the deep dermis.
We will witness precisely how this massless wave of energy intercepts a benign, life-giving oxygen molecule and violently mutates it into the most destructive, hyper-reactive biological explosive known to human physiology.
The destruction of your biological interface is not magic, and it is not simply the passage of time; it is a highly predictable, mathematically measurable thermodynamic event.

PHASE 1: THE PHOTON IMPACT
The excitation of dermal photosensitizers.
To initiate a catastrophic thermodynamic collapse within the skin, the incoming, massless UVA photon must first find a suitable biological target within the dermis capable of physically absorbing its specific electromagnetic frequency.
This is not a random, chaotic collision; it is a highly specific, lock-and-key quantum event governed by the strict laws of sub-atomic physics.
I. The Dermal Minefield
The deep dermal layer is absolutely not an empty, passive void waiting to be burned.
It is a dense, highly active, and heavily populated biological matrix saturated with naturally occurring endogenous molecules known to forensic biochemists as photosensitizers.
-
These unique molecular structures include porphyrins, riboflavin, specific advanced glycation end-products (AGEs), and various melanin precursors that naturally reside within the interstitial fluid and cellular boundaries of your skin.
-
These specialized molecules act as microscopic, highly sensitive cellular antennas. Because of their unique chemical architecture – specifically their alternating single and conjugated double bonds – their electron clouds are perfectly, mathematically tuned to catch the exact 315 to 400-nanometer frequency of the incoming UVA projectile.
-
When the UVA photon effortlessly bypasses the dead epidermal surface and plunges deep into the living dermis, it passes harmlessly through thousands of cells, completely ignored by most structures, until it directly strikes the receptive pi-electron cloud of one of these primed endogenous antennas.
II. The Kinetic Absorption
When the massless UVA photon collides with the pi-electron cloud of the endogenous photosensitizer, a violent and instantaneous transfer of kinetic energy occurs.
The photon ceases to exist as a traveling wave of light; its entire thermodynamic payload is completely and permanently absorbed by the host molecule.
-
The sudden, massive influx of pure solar radiation completely shatters the molecule’s baseline thermodynamic equilibrium.
-
This concentrated energy spike acts as a sub-atomic wrecking ball, directly impacting the negatively charged electron cloud orbiting the photosensitizer’s central nucleus.
III. The Unstable Excited State
Prior to the photon strike, the photosensitizer rested in a stable, comfortable, low-energy ground state. Upon absorbing the full kinetic force of the UVA photon, one of its outermost valence electrons is violently ejected from its normal orbit and thrust into a much higher, unoccupied molecular orbital.
-
This immediate sub-atomic displacement creates what quantum physicists call the “singlet excited state.”
-
It is an intensely unnatural, hyper-elevated, and profoundly unstable biological configuration. The entire molecule is now physically vibrating with the trapped kinetic energy of the solar radiation.
-
However, the molecule cannot maintain this hyper-elevated energetic posture. Through a process known as intersystem crossing, the molecule slightly shifts its electron spin, entering an even more dangerous “triplet excited state” that extends its unstable lifespan just long enough to inflict damage. The laws of thermodynamics dictate that this massive, unnatural energy payload cannot simply vanish; it must be urgently and aggressively discharged into the surrounding biological environment, setting the exact microscopic stage for the catastrophic secondary detonation.

PHASE 2: THE SPIN FLIP
The quantum transfer of energy to ground-state oxygen.
The excited dermal photosensitizer is now functioning as a microscopic thermodynamic time bomb, desperate to shed the kinetic energy it just absorbed from the sun.
It violently vibrates within the dense cellular matrix of the deep dermis, aggressively searching for a physical chemical conduit to offload its volatile, burning payload.
It finds this exact conduit in the most abundant, necessary, and universally available molecule in the human body: ground-state molecular oxygen (O2).
1. The Silent Bystander
Ground-state oxygen, known chemically in its natural form as triplet oxygen, is entirely benign and biologically essential for your survival, serving as the ultimate electron acceptor in mitochondrial ATP production.
It exists peacefully and abundantly within the extracellular fluid and the delicate lipid membranes of your dermal fibroblasts.
-
In its natural, unexcited triplet state, the oxygen molecule possesses two unpaired electrons located in its outermost molecular orbital.
-
Crucially, according to the strict, uncompromising rules of quantum mechanics (specifically the Pauli Exclusion Principle), these two outer electrons possess parallel spins. This means they are physically spinning in the exact same quantum direction.
-
This parallel spin configuration acts as a natural biological safety mechanism. It makes ground-state oxygen relatively inert and chemically stable, preventing it from spontaneously reacting with the organic, carbon-based molecules that make up your cellular scaffolding. Triplet oxygen is a peaceful bystander; it will not spontaneously attack your face.
2. The Intermolecular Collision
As the hyper-excited, violently vibrating photosensitizer molecule desperately seeks to discharge its trapped solar energy, it physically collides with the adjacent, benign triplet oxygen molecule in the dermal fluid.
This highly localized sub-atomic collision is the exact microsecond the true photo-oxidative crime is committed.
-
The photosensitizer does not simply bump into the oxygen molecule and bounce off; it executes a massive, direct quantum transfer of pure thermodynamic energy.
-
The trapped, burning kinetic energy of the UVA photon is forcefully expelled from the photosensitizer’s electron cloud and violently injected directly into the outer electron shell of the ground-state oxygen molecule.
3. The Quantum Inversion Event
This massive, unmitigated injection of thermodynamic energy forces an immediate, catastrophic violation of the oxygen molecule’s baseline physical and quantum geometry.
The transferred energy payload is so incredibly intense that it completely overpowers the natural quantum forces holding the oxygen electrons in their stable, parallel alignment.
-
The solar energy violently strikes one of the two unpaired outer electrons orbiting the oxygen nucleus.
-
It physically forces that specific electron to instantaneously invert its quantum rotation – an extremely violent atomic event known in physical chemistry as the “Spin Flip.”
-
Where the oxygen molecule previously had two electrons spinning parallel in perfect harmony, the immense solar payload has now forcefully slammed them into an unnatural, anti-parallel configuration. This single, microscopic sub-atomic inversion completely changes the physical, chemical, and behavioral identity of the oxygen molecule. The natural quantum restrictions have been bypassed. The benign, life-giving bystander has just been successfully weaponized by the sun.

PHASE 3: THE BIRTH OF SINGLET OXYGEN
The creation of a hyper-reactive biological explosive.
The quantum spin flip is now complete.
The benign, life-giving triplet oxygen molecule has been fundamentally and permanently mutated by the sun’s transferred thermodynamic energy.
It is no longer normal, ground-state O2.
It has been violently reborn as Singlet Oxygen – the ultimate thermodynamic weapon of the solar strike, the primary executioner of the dermis, and the leading suspect in the deep-tissue assassination of your youthful cellular architecture.
– The Thermodynamic Instability:
Singlet oxygen is not a standard free radical; it is a highly toxic, hyper-reactive, non-radical electrophilic species.
It functions exactly like a microscopic biological explosive.
Because its outer electrons have been violently forced into an unnatural, anti-parallel spin state, the entire molecule exists in a state of absolute, unyielding chemical panic.
It possesses a massive surplus of thermodynamic energy (exactly 94.2 kilojoules per mole above its ground state) that it simply cannot contain.
It is fundamentally unstable, highly volatile, and actively vibrating with the destructive kinetic payload originally delivered by the UVA projectile.
– The Ruthless Electron Hunger:
To survive this unnatural state and return to a stable, low-energy baseline, the singlet oxygen bomb must immediately shed this excess energy and chemically bond with another physical structure.
Because the quantum spin restriction has been lifted, singlet oxygen becomes a ruthless, highly aggressive, and indiscriminate electron scavenger.
It does not care about the biological integrity of your skin; it only cares about stealing an electron density to stabilize its own chaotic molecular orbital.
It is violently drawn to areas of high electron density, specifically carbon-carbon double bonds, and it will violently tear apart the nearest available biological structure to get what it needs.
– The Microscopic Blast Radius:
In the dense, crowded, highly packed biological environment of the deep dermis, the singlet oxygen bomb does not have to travel very far to find a structural victim.
Its half-life in a biological lipid environment is incredibly short – measured in mere fractions of a microsecond.
This extremely short lifespan means that its blast radius is highly localized and incredibly precise.
The exact moment it is born from the photosensitizer collision, it immediately detonates against whatever physical cellular structure happens to be closest to it.
– The Targeting of the Foundation:
Unfortunately for the Bio-Architect, these singlet oxygen detonations do not occur in empty, harmless space.
They occur directly adjacent to the most critical, structural load-bearing walls of your biological interface.
If the detonation occurs near the nucleus, the singlet oxygen bomb smashes directly against the DNA of your dermal fibroblasts, physically mutating the genetic source code required to produce fresh, youthful collagen.
If it detonates in the interstitial space, it rapidly oxidizes the extracellular matrix, permanently stiffening the tissue.
– The Ultimate Casualty:
But the absolute most devastating target of the singlet oxygen explosive – the specific, kinetic strike that mathematically causes the permanent, visible sagging, the profound loss of elasticity, and the deep, irreversible wrinkling of clinical photo-aging – is its direct, highly targeted assault on the delicate, polyunsaturated lipid membranes surrounding the dermal cells.
The massless photon of light has successfully bypassed your topical SPF lotion, descended into the dermis, and weaponized your own oxygen to physically shatter the lipid walls of your youth.
The thermodynamic fire has been officially lit inside the fortress, and the structural collapse is about to begin.

1.3 THE CRIME SCENE:
LIPID PEROXIDATION AND ARCHITECTURAL COLLAPSE
How singlet oxygen rips apart the polyunsaturated lipid grid, triggering a self-propagating chain reaction of structural decay.
When the singlet oxygen bomb detonates, the first casualty is the load-bearing wall of the cell: the lipid membrane.
The detonation is not a concussive blast but a chemical one, a wave of thermodynamic violence that does not seek to shatter structures with force, but to dismantle them with an insatiable hunger for electrons.
And in the densely packed, highly organized environment of the dermis, the target is never random. It is always the most vulnerable, the most electrically inviting, the most structurally critical component.
The first structure to fall is the very boundary that defines life from non-life, the delicate phospholipid bilayer that encases every fibroblast, every mast cell, every living component of your skin’s architectural foundation.
This is not collateral damage. This is the primary objective of the solar assassination.

I. THE VULNERABLE TARGET
The fatal attraction of polyunsaturated fatty acids.
The hyper-reactivity of singlet oxygen is a matter of pure quantum physics – an electron in a high-energy orbital that must return to a ground state.
To do so, it must steal electrons from another molecule.
It is, in essence, a microscopic black hole of energy. In the biological landscape of the dermis, it scans its surroundings for the easiest possible victim, the molecular structure that will offer the least resistance.
It finds this target in the polyunsaturated fatty acids – the PUFAs – that form the backbone of every cell membrane. Molecules like Linoleic Acid (LA) are not just fats; they are highly specific engineering materials, prized for the double bonds (C=C) along their carbon chains.
These double bonds create kinks in the structure, preventing the lipids from packing too tightly and thus ensuring the membrane remains fluid, flexible, and functional.
This fluidity is life. It allows proteins to move, signals to be transmitted, and nutrients to pass. But this beautiful, life-giving architecture is also a fatal flaw. Each double bond is a zone of high electron density, a rich, exposed electrical terminal. To the marauding singlet oxygen, this is an irresistible target.
It is drawn to these double bonds with the inevitability of a predator drawn to a wounded animal. The attack is not a choice; it is a chemical imperative.
The singlet oxygen does not need to break through a wall; the door is wide open, and the vault is unlocked.
It plunges toward the PUFA, its sole intent to quench its thermodynamic rage by stealing the very electrons that give the membrane its life.

II. THE CHAIN REACTION
The self-propagating wildfire of lipid peroxidation.
The initial strike of a single singlet oxygen molecule on a single lipid is not the end of the event; it is the beginning of a wildfire.
This is the self-propagating, catastrophic process known as Lipid Peroxidation, a chemical chain reaction that, once initiated, no longer requires the sun.
It becomes its own engine of destruction, burning across the cell membrane with terrifying speed and efficiency.
The Hydrogen Theft:
The assault begins with a violent abstraction.
The singlet oxygen, in its desperate search for stability, rips a hydrogen atom directly from a carbon atom adjacent to a double bond on the PUFA chain.
This act of quantum violence instantly destabilizes the lipid.
It is no longer a stable structural component; it has been transformed into a highly unstable, aggressive molecule known as a lipid radical.
It is a crippled molecule now turned into a predator, possessing its own unpaired electron and its own urgent need to steal from its neighbors to restore its balance.
The Oxygen Addition:
The newly formed lipid radical is in an untenable state and reacts with the most abundant molecule in its vicinity: ground-state molecular oxygen (O2).
This reaction is almost instantaneous.
The lipid radical combines with oxygen to form a new, even more destructive entity: a lipid peroxyl radical.
This molecule is the equivalent of a flaming piece of shrapnel ejected from the initial explosion.
It carries the oxidative damage forward, becoming the primary vector for the spreading fire.
It is a hybrid of a biological structure and a hyper-reactive radical, a zombie molecule whose only function is to create more of itself.
The Unstoppable Wildfire:
The lipid peroxyl radical now attacks an adjacent, healthy PUFA molecule.
It rips a hydrogen atom from its neighbor, satisfying its own electronic instability. But in doing so, it turns the victim molecule into a new lipid radical.
This new radical then reacts with oxygen, forms another lipid peroxyl radical, and the cycle repeats.
One becomes two, two become four, four become sixteen. It is a chain reaction of devastating elegance, a cascade of destruction that propagates exponentially across the entire plane of the cell membrane. The fire now fuels itself.
A single UVA photon that created a single singlet oxygen molecule has now triggered a biological meltdown that can destroy millions of lipid molecules, turning a fluid, dynamic membrane into a brittle, cross-linked, and dysfunctional ruin.

III. STRUCTURAL SHATTERING
From microscopic membrane rupture to macroscopic wrinkles.
The consequences of this microscopic wildfire are absolute.
A cell whose membrane has been consumed by lipid peroxidation is a city whose walls have been turned to dust. The membrane loses its defining characteristic: fluidity.
It becomes stiff, brittle, and porous.
Its core functions cease.
It can no longer maintain the delicate electrochemical gradient necessary for cellular energy.
It can no longer receive signals from the outside world. It can no longer regulate the passage of nutrients and waste.
The fibroblast, the skin’s master builder, the factory that synthesizes the collagen and elastin that give your skin its structure and bounce, is now functionally dead. But its death is not a quiet one.
A dying fibroblast does not simply cease production; it actively participates in the demolition of the surrounding architecture.
In its death throes, the shattered cell signals the release of a class of enzymes that act as a biological demolition crew: Matrix Metalloproteinases (MMPs). Specifically, it unleashes MMP-1, also known as collagenase.
These enzymes are molecular scissors. In a healthy system, they are used to break down old, damaged collagen to make way for new structures. But in the chronically inflamed, oxidatively-ravaged environment of a sun-damaged dermis, they become rogue agents of destruction.
They begin to indiscriminately sever the healthy, load-bearing Type I and Type III collagen fibers that form the skin’s foundational matrix.
The steel cables holding up the entire structure are being cut, one by one, from within. This internal collapse is the true genesis of a wrinkle.
The epidermis, the visible surface layer, loses its underlying support structure. It caves in.
The sagging of the jawline, the crow’s feet, the deep lines etched into the forehead – these are not surface creases.
They are the macroscopic, visible evidence of a microscopic crime scene, the topographical map of an architectural collapse that occurred deep within the dermal foundation, a silent, inside-out burn that began with a single photon and ended in a catastrophic structural failure.

CONCLUSION:
THE INADEQUACY OF SURFACE DEFENSE
Why stopping the thermodynamic collapse requires a defensive mechanism physically anchored inside the cell membrane.
The autopsy is complete.
The forensic report on photo-aging is written not on the skin’s surface, but in the catastrophic failure of its deep architecture.
The true epicenter of the crime is a thermodynamic fire.
It is a quantum-level event ignited by a massless photon, weaponized into a chemical explosive, and unleashed as a self-propagating wildfire within the lipid grid of your dermal cells.
The wrinkle you see in the mirror is merely the shadow cast by this internal collapse.
The assassination is an inside job.

I. THE MATHEMATICAL FAILURE
You cannot paint over an internal fire.
With this forensic knowledge, the profound inadequacy of the entire modern skincare paradigm becomes starkly, mathematically clear.
The industry has sold you a fortress built on sand, a defense predicated on a fatal misunderstanding of the battlefield.
The strategy fails at three distinct levels of physical reality:
1. The Positional Failure:
The industry’s primary defense – topical sunscreen – is applied to the stratum corneum.
This is a graveyard of dead cells, located hundreds of microns above the living, breathing fibroblasts of the dermis where the actual damage occurs.
The fortress is built miles away from the city it is supposed to protect.
2. The Temporal Failure:
A topical lotion can only attempt to block a photon before it penetrates. Once a UVA projectile has breached this superficial barrier, the lotion becomes irrelevant.
It is a shield left behind on the shore after the enemy has already landed.
It has no mechanism to combat a fire that is already burning inside the walls.
3. The Thermodynamic Failure:
The true weapon is not the photon itself, but the singlet oxygen it creates deep within the tissue.
A cream on the surface has zero physical or chemical capacity to reach into the dermis and neutralize this hyper-reactive molecule.
It is like trying to extinguish a house fire by holding a bucket of water outside the locked front door.
It is a comforting ritual, but a mathematical impossibility.
It is a strategy doomed to fail.

II. THE CALL FOR THE HEAT-SINK
The necessity of an endogenous, transmembrane interceptor.
To stop this inside-out burning, we must abandon the surface and deploy a defense mechanism at the point of detonation. The solution cannot be topical; it must be systemic.
It must be woven into the very architecture it is designed to protect.
This requires an engineer, not a cosmetician.
We must build a molecule with a precise set of architectural specifications:
Systemic Delivery:
It must be delivered via the bloodstream, reaching the deep dermal layers where the fibroblasts reside.
Architectural Integration:
It must possess the unique lipophilic and hydrophilic properties to physically insert itself into and span the entire 30-Ångström width of the vulnerable cell membrane.
It cannot simply sit on top; it must become part of the wall.
Thermodynamic Quenching:
It must function as a [Thermodynamic Heat-Sink].
It must have the specific electronic configuration to intercept the violent energy of singlet oxygen, absorb it, and dissipate it harmlessly as heat, all without being destroyed in the process.
This is not a task for a simple antioxidant.
This requires a specialized, architectural-grade defender designed for the unique violence of the dermal environment.
In Chapter 2, we will deploy this ultimate weapon.
We will introduce the only molecule known to science with the precise stereochemistry required to meet these engineering demands, the ultimate biological quencher of thermodynamic fire:
Astaxanthin

References for Chapter 1
Pinnell, S. R. (2003). Cutaneous photodamage, oxidative stress, and topical antioxidant protection. Journal of the American Academy of Dermatology, 48(1), 1–22.
Rittié, L., & Fisher, G. J. (2015). UV-light-induced signal cascades and skin aging. Ageing Research Reviews, 2(2), 1-15.
Kochevar, I. E., Pathak, M. A., & Parrish, J. A. (1999). Photophysics, photochemistry, and photobiology. In Fitzpatrick’s Dermatology in General Medicine (5th ed., pp. 220-229). McGraw-Hill.
Girotti, A. W. (2001). Photosensitized oxidation of membrane lipids: reaction pathways, cytotoxic effects, and cytoprotective mechanisms. Journal of Photochemistry and Photobiology B: Biology, 63(1-3), 103-113.
Gruber, F., Kremslehner, C., & Kähäri, V. M. (2014). Fibroblast-derived matrix metalloproteinases: a link between inflammation and connective tissue damage. The Journal of Investigative Dermatology, 134(11), 2697-2700.
Sies, H., & Stahl, W. (1995). Vitamins E and C, beta-carotene, and other carotenoids as antioxidants. The American Journal of Clinical Nutrition, 62(6 Suppl), 1315S–1321S.
Prá M, et al. (2006). UVA-induced singlet oxygen and cyclobutane pyrimidine dimers-DNA damage in keratinocytes. Journal of Photochemistry and Photobiology B: Biology, 84(1), 14-20.
Niki, E., Yoshida, Y., Saito, Y., & Noguchi, N. (2005). Lipid peroxidation: Mechanisms, inhibition, and biological effects. Biochemical and Biophysical Research Communications, 338(1), 668-676.
Fisher, G. J., Kang, S., Varani, J., Bata-Csorgo, Z., Wan, Y., Datta, S., & Voorhees, J. J. (2002). Mechanisms of photoaging and chronological skin aging. Archives of Dermatology, 138(11), 1462–1470.
Tyrrell, R. M. (1996). UVA (320-380 nm) radiation as a modulator of skin function. Journal of Photochemistry and Photobiology B: Biology, 35(3), 195-201.
Gaspar, L. R., & Maia Campos, P. M. B. G. (2011). Evaluation of the efficacy of a sunscreen system. International Journal of Pharmaceutics, 407(1-2), 125-131.
Davies, M. J. (2004). Reactive oxygen species, metalloproteinases, and cancer. Free Radical Biology and Medicine, 36(7), 823-833.
Halliwell, B., & Gutteridge, J. M. C. (2015). Free Radicals in Biology and Medicine (5th ed.). Oxford University Press.
Cadet, J., & Douki, T. (2018). Formation of UV-induced DNA damage. Photochemical & Photobiological Sciences, 17(12), 1816-1841.
Pattison, D. I., & Davies, M. J. (2006). Actions of UVA and UVB radiation on the antioxidant and protein components of the stratum corneum. Free Radical Biology & Medicine, 41(10), 1594-1605.
Baier, J., Maisch, T., Maier, M., Engel, E., Landthaler, M., & Bäumler, W. (2006). Singlet oxygen generation by UVA light. Experimental Dermatology, 15(1), 12-16.
D’Orazio, J., Jarrett, S., Amaro-Ortiz, A., & Scott, T. (2013). UV radiation and the skin. International Journal of Molecular Sciences, 14(6), 12222–12248.
Marionnet, C., Pierrard, C., Golebiewski, C., & Bernerd, F. (2014). Diversity of biological effects induced by longwave UVA rays (UVA1) in reconstructed skin. PLoS One, 9(7), e102148.
Shindo, Y., Witt, E., & Packer, L. (1994). Antioxidant defense mechanisms in murine epidermis and dermis and their responses to ultraviolet light. The Journal of Investigative Dermatology, 102(4), 470–475.
Trautinger, F. (2001). Mechanisms of photodamage of the skin and its functional consequences for skin ageing. Clinical and Experimental Dermatology, 26(7), 573-577.
Wang, S. Q., & Balagula, Y. (2010). The failure of sunscreen. Dermatologic Clinics, 28(4), 629-633.
Quan, T., Qin, Z., Xia, W., Shao, Y., Voorhees, J. J., & Fisher, G. J. (2009). Matrix-degrading metalloproteinases in photoaging. Journal of Investigative Dermatology Symposium Proceedings, 14(1), 20-24.
Sander, C. S., Chang, H., Salzmann, S., Müller, C. S., Ekanayake-Mudiyanselage, S., Elsner, P., & Thiele, J. J. (2002). Photoaging is associated with protein oxidation in human skin in vivo. The Journal of Investigative Dermatology, 118(4), 618-625.
Vayalil, P. K., Mittal, A., & Katiyar, S. K. (2003). Proanthocyanidins from grape seeds inhibit expression of matrix metalloproteinases in human dermal fibroblasts to protect against the effects of UVA radiation. The Journal of Investigative Dermatology, 121(2), 411-417.
Beani, J. C. (2004). Porphyrins, drugs and photosensitization. Photochemical & Photobiological Sciences, 3(8), 789-801.

# KNOWLEDGE SUMMARY: ANATOMY OF A SOLAR ASSASSINATION
## I. THE CRIME SCENE & THE DECEPTION (EPIDERMIS VS. DERMIS)
* **The Deception:** Consumers are conditioned to fear **Erythema** (sunburn), a superficial inflammatory response primarily in the epidermis caused by UVB.
* **The False Assumption:** “No redness = No damage.”
* **The Forensic Reality:** The true “epicenter” of structural photo-aging (wrinkles, sagging) is the **Dermis**, the load-bearing architectural layer.
* **The Primary Target:** Dermal **Fibroblasts**, the factories for Collagen and Elastin.
## II. THE WEAPONS: A BALLISTIC ANALYSIS (UVA VS. UVB)
* **The Decoy (UVB):**
* **Wavelength:** 280-315nm (Short-wave).
* **Energy Profile:** High-energy, low-penetration.
* **Trajectory:** Almost entirely absorbed by the stratum corneum and epidermis.
* **Effect:** Acute, visible inflammation (sunburn), immediate pain signals, direct DNA damage. It is the “loud” alarm.
* **The Assassin (UVA):**
* **Wavelength:** 315-400nm (Long-wave).
* **Energy Profile:** Lower-energy, extreme-penetration.
* **Trajectory:** Passes through clouds, glass, and the epidermal layers with minimal resistance, delivering its kinetic energy payload directly into the dermis.
* **Effect:** Silent, deep-tissue damage. It strikes fibroblasts without triggering immediate pain or visible erythema. It is the “silent” killer.
* **The Inevitable Breach (SPF Failure):**
* **Mathematical Certainty:** SPF (Sun Protection Factor) is a measure of time-to-reddening (UVB), not a measure of UVA blockage.
* **The Leakage:** Even a perfectly applied SPF 50 allows a percentage of UVA photons to penetrate. Human error (insufficient application, sweat, friction) dramatically increases this leakage rate.
* **The Point of No Return:** Once a UVA photon crosses the epidermal barrier, the external lotion is rendered completely irrelevant to the subsequent thermodynamic cascade.

## III. THE EXPLOSIVE DEVICE: THE SINGLET OXYGEN CASCADE
* **The Physics Problem:** How does massless light (a photon) destroy a physical cell?
* **The Answer:** A three-phase transfer of thermodynamic energy that weaponizes benign oxygen.
* **PHASE 1: The Photon Impact (Excitation):**
* A UVA photon strikes an **endogenous photosensitizer** (e.g., porphyrins, riboflavin) in the dermis.
* The photosensitizer absorbs the photon’s kinetic energy, transitioning to a highly unstable, high-energy “excited singlet state.”
* **PHASE 2: The Spin Flip (Energy Transfer):**
* The unstable photosensitizer must shed its excess energy. It collides with a nearby, stable, ground-state oxygen molecule (Triplet Oxygen, ³O₂).
* The energy is transferred, forcing a quantum mechanical **”electron spin flip”** in the oxygen’s outer orbital.
* **PHASE 3: The Detonation (Singlet Oxygen Formation):**
* The result of the spin flip is **Singlet Oxygen (^1O2)**. It is no longer stable oxygen; it is a hyper-reactive, cytotoxic explosive with a half-life measured in nanoseconds.
* It desperately seeks to steal electrons to return to a stable state, initiating a chemical attack on the nearest biological structure.
## IV. THE COLLATERAL DAMAGE: LIPID PEROXIDATION & STRUCTURAL COLLAPSE
* **The First Victim:** The **Polyunsaturated Fatty Acids (PUFAs)** in the cell membranes of fibroblasts. Their double bonds (C=C) are regions of high electron density, making them fatally attractive to singlet oxygen.
* **The Self-Propagating Chain Reaction (The Wildfire):**
1. **Initiation (Hydrogen Theft):** Singlet Oxygen rips a hydrogen atom from a PUFA, creating a **lipid radical**.
2. **Propagation (Oxygen Addition):** The lipid radical reacts with ambient oxygen to form a **lipid peroxyl radical**. This new radical then steals a hydrogen atom from an adjacent healthy PUFA, creating another lipid radical.
3. **The Cascade:** This process repeats exponentially, burning across the membrane and turning it from a fluid structure into a brittle, cross-linked, and non-functional ruin.
* **The Architectural Consequence (Macroscopic Wrinkles):**
1. **Membrane Failure:** The fibroblast membrane shatters, losing its ability to function.
2. **MMP Release:** The dying fibroblast releases **Matrix Metalloproteinases (MMPs)**, specifically MMP-1 (collagenase).
3. **Collagen Degradation:** These MMPs act as molecular scissors, severing the existing collagen network that supports the epidermis.
4. **The Collapse:** The epidermis caves into the now-empty dermal space, forming a visible wrinkle. The skin has collapsed from the inside out.
## V. THE STRATEGIC CONCLUSION
* **The Futility of Surface Defense:** Applying topical SPF is like painting the roof of a house while a fire burns in the foundation. It cannot stop the internal, thermodynamic reaction once the initial photon has breached the perimeter.
* **The Engineering Imperative:** To prevent this collapse, a defensive mechanism must be physically present **inside the cell membrane** itself. It must be a **[Thermodynamic Heat-Sink]** capable of intercepting and safely dissipating the photon’s energy before it can weaponize oxygen.

Chapter 2: The Thermodynamic Heat-Sink
Deploying the Astaxanthin Commander
How a 30-Angstrom transmembrane shield intercepts singlet oxygen, dissipating lethal UV radiation into harmless heat without sacrificing its own structural integrity.
You step outside, and the invisible bombardment begins instantly, subjecting your dermal matrix to a relentless, high – velocity strike of ultraviolet radiation for the next eleven hours.
The beauty industry standard protocol is to throw water – soluble Vitamin C at this roaring oxidative fire, blindly hoping to neutralize the escalating threat.
We must objectively declare this not just ineffective, but a tragic, mathematically flawed tactic of chemical suicide.
Your cellular architecture is drowning in secondary oxidative damage while the primary radiation threat continues completely unabated.
This traditional approach represents a fundamental, catastrophic misunderstanding of thermodynamic biological defense.

I. THE SACRIFICIAL PAWN
The rapid depletion of conventional antioxidants.
Let us forensically examine the exact molecular tragedy of the conventional antioxidant when it is deployed in a high – radiation environment.
You know the specific, tightening sensation of your skin after hours in the sun; this is not merely dehydration, but the tactile echo of your cellular scaffolding being systematically dismantled.
When ultraviolet photons penetrate the dermal perimeter, they act as highly lethal molecular snipers. They forcibly eject single electrons from otherwise stable structural lipids, instantly birthing frantic, highly reactive free radicals.
The traditional medical response to this [The Neuro – Endocrine Storm] of localized oxidative stress is to flood the area with standard, water – soluble antioxidants.
These molecules operate on a primitive, inherently flawed mechanism of electron donation. They rush toward the volatile free radical and willingly surrender one of their own outer electrons to pacify the immediate threat.
However, you must recognize the catastrophic biological flaw in this biochemical transaction.
The exact moment the conventional antioxidant donates its electron, it commits irreversible chemical suicide. It renders itself completely and permanently useless for any future defense.
Furthermore, the biochemistry gets significantly darker as these depleted, oxidized molecules undergo a dangerous molecular pivot.
In their exhausted, unstable state, these former cellular protectors frequently transform into weak pro – oxidants themselves. They become the very oxidative shrapnel they were originally deployed to eliminate, actively contributing to the inflammatory burden of the surrounding tissue.
Now, apply the brutal, unforgiving mathematics of continuous solar exposure to this primitive defense system. A standard, unrelenting ultraviolet strike generates tens of millions of radicalized oxygen species per microsecond across the vast topography of your dermal architecture.
A finite application of a conventional topical serum, or a standard oral vitamin, contains a strictly limited battalion of these electron donors.
Under the heavy, continuous artillery fire of UVA and UVB radiation, this sacrificial supply is rapidly and entirely exhausted within a matter of mere minutes.
The biological reality is stark: your defense strategy simply ran out of ammunition in a war of attrition.
The front lines inevitably collapse, and the remaining hours of daylight become a microscopic slaughterhouse for your unprotected collagen and elastin networks.

II. THE CALL FOR A THERMODYNAMIC HEAT – SINK
Moving from chemical sacrifice to physical dissipation.
To survive a continuous, eleven – hour solar assault, your skin cannot possibly rely on a failing strategy of chemical martyrdom.
The foundational logic of the traditional skincare industry is mathematically broken. Your biology does not need a higher concentration of sacrificial pawns rushing blindly to their deaths in the deep dermal layers.
What the human cellular matrix actually requires to withstand this relentless bombardment is a permanent, non – destructive structural shield capable of true biophysical endurance.
It demands an architectural paradigm shift. The defending molecule must absorb catastrophic energetic payloads and dissipate them harmlessly into the ether, all while retaining its absolute structural integrity.
This is the explicit call for a thermodynamic heat – sink, marking the exact moment we abandon the tragic limitations of conventional vitamins.
We must now introduce the supreme, uncompromising architect of deep – cellular defense, [The Systemic Commander], known to the scientific community as Astaxanthin.
Unlike its primitive, electron – donating predecessors, the Commander operates strictly on the advanced laws of biophysical quenching rather than chemical sacrifice.
Extracted from deep – sea extremophiles forged in the harshest UV environments on the planet, this highly specialized molecule is engineered by nature for absolute endurance under apocalyptic levels of solar radiation.
When strategically integrated into the human biological system, it functions as a permanent, vibrating shock absorber that locks directly into the cellular membrane.
It catches the kinetic fury of the ultraviolet photon, traps the volatile singlet oxygen bomb, and physically bleeds the lethal excitation energy away into the surrounding cellular fluid as microscopic, harmless heat.
Crucially, it executes this thermodynamic dissipation thousands of times per second, for hours on end, without ever breaking its own carbon backbone or becoming a pro – oxidant.
This unbreakable physical defense is the non – negotiable baseline requirement for the Keyora biological protocol.
According to our foundational 1+1+1+1+1+1+1 > 7 synergy rule, we cannot deploy the sensitive lipid vanguard into the combat zone without first establishing this absolute thermodynamic cover fire.
Without the Commander securing the grid, the fragile essential fatty acids would be instantly oxidized into toxic malondialdehyde the moment they entered the irradiated dermis.
The Commander holds the line.
It ensures that the biological architects can safely reconstruct the evaporated moisture barrier and repair the collapsed micro – vascular networks.

2.1 THE 30-ANGSTROM ANCHOR:
SECURING THE DERMAL PERIMETER
How Astaxanthin’s precise molecular geometry allows it to structurally interlock with the cellular lipid bilayer.
A shield is entirely useless if it simply floats away during the chaotic heat of battle.
You know the exact physical sensation of a failing dermal barrier. It is that paper-thin, tight, and burning fragility you feel late in the afternoon, where the cellular glue holding your face together seems to be actively dissolving.
This deep tissue exhaustion is the direct result of relying on localized, transient antioxidants that temporarily wash over the cells but never actually secure the perimeter.
The true power of the Commander does not lie merely in its ability to absorb a photon. Its absolute biological supremacy is rooted in its uncompromising physical lockdown within your microscopic architecture.
It does not just stand in front of the wall.
It becomes the wall.

PHASE 1: THE DEEP DERMAL DELIVERY
Penetrating the systemic barrier.
The journey of true structural protection must be systemic, not merely a superficial topical application.
When ingested, this highly lipophilic molecule embarks on a complex circulatory expedition.
It binds to lipoproteins in the liver and travels through the central bloodstream, ultimately deploying into the deepest, most vulnerable territories of your skin.
It specifically targets the exhausted dermal fibroblasts and the rapidly dividing basal keratinocytes that form the foundation of your face.
First, The Capillary Exit:
The delivery protocol requires extreme physiological precision. As the blood pumps through the microscopic, fragile capillary networks feeding the lower layers of the dermis, the Commander detects the localized [The Neuro-Endocrine Storm] of inflammatory distress and severe UV radiation damage.
It physically exits the microvasculature, slipping through the tight endothelial gaps and entering the dried-out extracellular matrix.
It navigates through this devastated, irradiated space like a targeted biological drone, relentlessly seeking the exact molecular coordinates of structural failure.
Second, The Cellular Approach:
Astaxanthin is intensely, fundamentally lipophilic. It is naturally repelled by the watery, chaotic environment of the extracellular fluid and magnetically drawn toward the fatty, lipid-rich architecture of your cellular membranes.
As it approaches a distressed, structurally compromised fibroblast, it does not bounce off the surface like a common water-soluble vitamin. Instead, it is actively pulled into the cellular envelope by an undeniable, powerful thermodynamic attraction.
It prepares to execute a structural integration that will entirely redefine the cell’s mechanical survivability.

PHASE 2: THE TRANSMEMBRANE LOCK
The perfect geometric fit.
What happens next is a masterpiece of evolutionary bio-architecture. This is not a fleeting chemical reaction or a temporary donation of an electron.
It is a profound, permanent physical docking process that fundamentally alters the tensile strength of the cell.
1. The Hydrophilic Polar Heads:
Examine the exact molecular anatomy of Astaxanthin. At both extreme ends of its massive carbon structure lie highly specialized, oxygen-rich ionone rings. These are the hydrophilic polar heads.
Because these specific rings have a profound chemical affinity for water, they instantly gravitate toward the aqueous environments on both the absolute exterior and the absolute interior of the cell.
The top ring physically anchors itself into the extracellular fluid just outside the cell membrane.
Simultaneously, the bottom ring locks firmly into the watery intracellular cytoplasm deep inside the cell.
2. The Hydrophobic Conjugated Chain:
Between these two anchored polar rings stretches a massive, hydrophobic conjugated polyene chain.
This long carbon backbone chemically despises water, forcing it to bury itself deep within the fatty, lipid core of the membrane. Here is the mathematical miracle of [The Systemic Commander].
The exact length of this rigid carbon chain perfectly matches the 30-Angstrom width of the human cellular lipid bilayer.
It is not a single micrometer too short, nor a single micrometer too long.
It spans the precise, exact distance required to bridge the entire structural gap.
3. The Vertical Stabilization:
By locking its polar heads on the outside and inside, and stretching its rigid spine straight through the center, Astaxanthin forms a permanent, vertical rivet directly through the cell membrane.
It behaves exactly like a heavy steel rebar plunged vertically through setting concrete. This vertical stabilization physically restricts the chaotic, fluid movement of the surrounding lipids.
It stops the membrane from leaking moisture.
It drastically increases the cell’s mechanical resistance to the shearing, destructive forces of ultraviolet radiation, turning a fragile, easily ruptured biological balloon into a heavily reinforced bio-architectural fortress.

PHASE 3: THE OMNIPRESENT SHIELD
Dual-sided protection.
Because this 30-Angstrom rivet successfully spans the entire architectural depth of the cellular membrane, it achieves a state of biological omnipresence. It is fundamentally impossible for standard antioxidants to replicate this feat.
Vitamin C floats blindly outside the cell in the water.
Vitamin E floats blindly inside the fat.
Neither can see the whole battlefield.
The Commander intercepts the incoming, high-velocity barrage of UV-induced free radicals crashing against the exterior wall.
Simultaneously, it reaches inward to absorb the secondary wave of radicalized oxygen leaking from the cell’s own exhausted, over-worked mitochondria deep within the interior cytoplasm.
It offers an unbroken, dual-sided thermodynamic shield that commands both the external horizon and the internal cellular machinery at the exact same time. This is absolute perimeter control.

2.2 THE PHYSICS OF QUENCHING:
DISSIPATING THE SINGLET OXYGEN BOMB
The non-destructive thermodynamic mechanism that neutralize UV-induced excitation energy without electron loss.
When the singlet oxygen bomb detonates in the deep dermis, the Commander does not offer an electron.
It offers an inescapable thermodynamic trap.
You know the exact physical sensation of this micro-detonation. It is that deep, radiating, uncomfortable heat that persists in your skin long after you have moved out of the sun and into a dark room.
That is not just residual environmental warmth.
That is the actual physical friction of your microscopic cellular scaffolding being violently torn apart by hyper-excited oxygen.
Standard skincare tries to put out this localized chemical fire by throwing fragile, water-soluble vitamins directly into the blaze.
We must permanently abandon that failing, mathematically flawed strategy. The Commander rewrites the very physics of the biological battlefield instead.

1. THE ENERGY INTERCEPTION
Catching the quantum wave.
Before we can effectively neutralize the immediate threat, we must freeze time and forensically examine the exact microsecond of impact.
I. The Singlet Oxygen Collision:
The relentless afternoon solar radiation acts as a continuous kinetic strike against your vulnerable biology.
An incoming ultraviolet photon hits an ordinary, passive oxygen molecule resting quietly inside your deep dermal tissue.
The sheer velocity of the photon collision violently kicks an electron into a higher, wildly unstable orbital state.
This is the instant, catastrophic birth of Singlet Oxygen.
This is the most chaotic, destructive manifestation of the localized oxidative stress.
-
It is what Keyora defines as [The Neuro-Endocrine Storm], reduced down to the scale of a single, panicked atom.
-
It is no longer a life-giving molecule. It is a microscopic bomb moving at terrifying speed toward the delicate lipid walls of your cells.
-
It is desperately looking to unload its unnatural burden of lethal excitation energy onto any stable structure it can find.
-
If it hits an unprotected fatty acid, it ignites an unstoppable, cascading chain reaction of lipid peroxidation.
Your essential moisture barrier simply dissolves, leaving the tissue vulnerable to profound dehydration and structural collapse.
II. The Polyene Chain Absorption:
But in a Keyora-fortified cellular matrix, this rogue, hyper-excited molecule does not hit a fragile lipid.
It hits a heavily reinforced, 30-Angstrom biological wall.
It slams directly into the massive, immovable, and vertically anchored structure of Astaxanthin.
Because [The Systemic Commander] is locked perfectly across the entire span of the lipid bilayer, it stands dead center in the absolute blast radius.
The hyper-excited oxygen collides with the shield, but the Commander absolutely refuses to chemically react.
Instead, its long, central conjugated polyene chain instantly activates to meet the threat.
This massive carbon backbone is densely packed with alternating double and single bonds.
This specific molecular geometry creates a dense, impenetrable cloud of delocalized electrons.
-
It functions exactly like a microscopic, highly advanced electromagnetic net suspended in the cellular membrane.
-
It simply catches the raw, volatile excitation energy.
-
It physically strips the lethal quantum payload directly from the attacking oxygen, without allowing a single chemical bond to be permanently broken or formed.

2. THE VIBRATIONAL COOLING
Converting radiation into harmless heat.
The lethal solar energy has now been successfully intercepted.
The bomb has been effectively caught in the net.
But the sheer kinetic energy is still physically present inside the cellular architecture.
It must be safely deactivated and expelled before it tears the defender apart from the inside out.
First, The Molecular Resonance:
Here is the absolute biological pivot where Keyora leaves traditional, primitive medicine completely behind.
Astaxanthin absolutely does NOT donate an electron to solve this crisis.As the massive influx of absorbed solar energy surges into its carbon backbone, the entire molecule begins to vibrate.
It vibrates intensely.
Violently.
Microscopically.
The absorbed lethal energy is channeled directly into the mechanical, physical motion of the carbon-carbon bonds.
The entire Astaxanthin molecule acts as a perfectly tuned biological tuning fork, violently humming with the trapped, raw energy of the sun.
It is actively holding the fire.
It contains the devastating blast radius strictly within its own rigid structural geometry.
It absolutely refuses to let that lethal energy spill over into your highly vulnerable surrounding tissues.
Second, The Thermal Dissipation:
But a molecule cannot vibrate forever under such extreme duress. To survive, it must rapidly bleed the pressure away.
Through a highly sophisticated biophysical process, the Commander automatically initiates physical quenching. As the long carbon chain vibrates back and forth, it constantly rubs against the surrounding aqueous solvent.
It physically impacts against the dense lipid environment of the cell membrane thousands of times a millisecond. Through this continuous, microscopic mechanical friction, the lethal, high-velocity ultraviolet energy is systematically bled out.
It is released entirely into the surrounding cellular fluid as completely harmless, low-grade microscopic heat. The raw, cell-shattering destructive power of the photon has been effectively and permanently downgraded.
A catastrophic quantum strike is brilliantly transformed into a tiny, localized warming sensation that your biological system can easily manage and ignore.
The explosive threat has been methodically defused without a single casualty.

3. THE UNBROKEN COMMANDER
The survival of the shield.
We must now forensically examine the biological aftermath of this exact, highly coordinated thermodynamic intervention.
Look at the attacking singlet oxygen first.
Stripped entirely of its lethal excitation energy by the polyene net, the once-rogue molecule instantly drops back down. It returns to its harmless, docile, and biologically necessary ground state. The bomb is completely defused, and not a single electron ever changed hands to make it happen.
Now, look at the defender.
As the very last fraction of absorbed radiation is safely dissipated into the surrounding fluid as heat, the Astaxanthin molecule stops vibrating. It returns to a state of absolute, unbreakable structural stillness.
The 30-Angstrom anchor remains perfectly intact.
The hydrophilic polar heads are still firmly, aggressively gripping the inner and outer boundaries of the cell.
Not a single carbon bond has been broken.
Not a single electron has been lost to the oxidative void.
The Commander is completely structurally unbroken.
It is absolutely unoxidized, and instantly ready to absorb the very next ultraviolet strike that is guaranteed to hit a millisecond later.
It does not need to rest, and it does not need to be metabolically replaced by secondary vitamins.
This exact, flawless capacity for limitless, non-destructive thermodynamic cycling is its ultimate defining power.
This is the precise biological reason why Astaxanthin is mathematically calculated to be six thousand times stronger than Vitamin C.
It is the ultimate, unbreakable cellular bulwark against the sun.
It provides the absolute, mandatory, permanent cover fire required for the entire biological protocol to function.
Without this unbroken shield holding the line, the Keyora 1+1+1+1+1+1+1 > 7 synergy would collapse instantly.
The incoming builder lipids, like ALA and LA, would be oxidized and destroyed before they could reconstruct your skin.
The Commander holds the perimeter, so the architects can safely rebuild.

2.3 THE BROAD-SPECTRUM SCAVENGER:
NEUTRALIZING THE ROS/RNS FALLOUT
Trapping the secondary wave of oxidative shrapnel across the hydrophilic and lipophilic zones.
Singlet oxygen is the primary bomb, but the destruction does not end with the initial blast.
Ultraviolet radiation unleashes a chaotic, secondary storm of diverse free radicals across your entire cellular landscape.
This is the Reactive Oxygen and Nitrogen Species (ROS/RNS) fallout, a microscopic hailstorm of oxidative shrapnel. It mercilessly tears through both the watery exterior and the fatty interior of your deeply irradiated cells.
You feel this secondary fallout as that lingering, deep-tissue ache, the relentless dry stinging, and the terrifying loss of elasticity that haunts your skin long into the night.
To survive this specific, brutal phase of [The Neuro-Endocrine Storm], the Commander cannot just stand statically in one place.
It must actively sweep the entire biological battlefield, aggressively engaging multiple enemies simultaneously across distinct chemical zones.
PROPOSITION A:
How does Astaxanthin neutralize the diverse spectrum of oxidative shrapnel?

The Evidence:
Clinical data proves that [The Systemic Commander] entirely rejects the single-target limitations of primitive, conventional antioxidants.
It possesses the extraordinary, mathematically verifiable capacity to identify and scavenge multiple, radically different species of oxidative threats at the exact same time.
It does not wait passively for the damage to come to it.
It operates flawlessly and continuously across both the hydrophilic, watery environments and the lipophilic, fatty depths of your biological architecture.
It creates a three-dimensional zone of absolute exclusion.
– Superoxide Anions:
Deep within your exhausted cells, the mitochondria are actively suffocating under the continuous, crushing weight of UV stress.
As these microscopic cellular power plants begin to fail and overheat, they violently leak highly toxic Superoxide Anions directly into the intracellular cytoplasm. This internal toxic spill physically paralyzes the cell’s energy production, leading to the rapid, premature apoptosis of your essential fibroblasts.
Astaxanthin aggressively reaches inward from the membrane, utilizing its hydrophilic polar heads to intercept this specific, localized threat.
It traps the Superoxide Anions at the source, ensuring your dermal cells retain the sheer metabolic stamina required to stay alive, repair, and function.
– Hydroxyl Radicals:
Simultaneously, the aqueous zones resting just outside the cell membrane are rapidly flooding with Hydroxyl Radicals.
These are universally recognized by biochemists as the most violent, indiscriminate, and destructive molecules in the entire oxidative spectrum. They operate like microscopic buzzsaws, aggressively ripping apart cellular DNA, shredding vital structural proteins, and ruthlessly dissolving the collagen matrix on contact.
The Commander’s polar rings, anchored firmly in the extracellular fluid, act as an inescapable magnetic sink for these localized terrors.
It systematically sweeps the aqueous perimeter clean, preventing the Hydroxyl shrapnel from ever reaching the highly vulnerable lipid wall.
– Lipid Peroxyl Radicals:
Despite the absolute strongest structural defenses, some fraction of secondary radiation always manages to bleed into the deep, fatty core of the membrane.
This breach triggers the rapid birth of Lipid Peroxyl Radicals, the exact unstable molecules responsible for catastrophic, chain-reaction lipid peroxidation.
If left unchecked for even a microsecond, they will instantly target, oxidize, and destroy the fragile, essential fatty acids of our Keyora 1+1+1+1+1+1+1 > 7 protocol.
But the massive, rigid carbon backbone of Astaxanthin lies waiting exactly within this lipophilic core, perfectly positioned for a lethal intercept.
It aggressively hunts down the Peroxyl Radicals, physically stopping the chemical chain reaction cold in its tracks.
It provides the absolute, non-negotiable cover fire required for Linoleic Acid, Alpha-Linolenic Acid, and Oleic Acid to safely survive and operate within the membrane.

The Mechanism:
When dealing with this highly diverse, volatile secondary fallout, the physical quenching of pure energy is not always mathematically applicable.
These free radicals are distinct chemical entities actively missing electrons, behaving as rabid, unbalanced scavengers desperate to tear your tissue apart.
The Commander must deploy a secondary, highly advanced tactical biophysical response to neutralize them, all while strictly avoiding its own destruction.
1. Electron Trapping:
Unlike traditional, water-soluble vitamins that foolishly throw an electron away and immediately die in a tragic act of chemical suicide, Astaxanthin performs a masterclass in molecular judo.
Its highly conjugated double bonds actively, aggressively engage the incoming free radical.
Instead of donating one of its own vital electrons to the radical, the Commander physically pulls the radical’s unpaired electron into its own massive structure.
It absorbs the unstable, destructive threat directly into the dense electron cloud of its carbon backbone.
It safely traps the volatile particle inside a molecular cage, effectively disarming the biological explosive precisely on contact.
2. Delocalization:
Now, the Astaxanthin molecule is actively holding the dangerous, unpaired electron within its architecture.
In a primitive, conventional antioxidant, this exact scenario would immediately trigger structural oxidation and a lethal pro-oxidant transformation.
But the Commander possesses a remarkably long, highly conjugated polyene chain that functions exactly like a heavy-duty biological lightning rod. It instantly delocalizes the trapped radical energy, rapidly spreading the intense, localized burden evenly across its entire 30-Angstrom carbon length.
This is the exact molecular equivalent of grounding a massive, lethal electrical strike deep into the earth.
Through this massive structural distribution, the volatile energy of the free radical is rapidly diluted into absolute insignificance. The trapped electron becomes harmlessly suspended and permanently pacified within the electromagnetic net.
This flawless energy distribution leaves the Astaxanthin molecule entirely structurally unbroken, completely unoxidized, and biologically dominant.

The Resolution:
This is the undeniable, mathematically sound biophysical reality of true, uncompromising deep cellular defense.
By combining the precise, targeted interception of intracellular Superoxide and extracellular Hydroxyls with the deep-membrane trapping of Peroxyls, Astaxanthin achieves total environmental dominance.
-
It provides absolute, unwavering, broad-spectrum clearance of the entire oxidative fallout.
-
It successfully ensures that the complete lipid vanguard, from the Oleic Acid spacer to the Alpha-Linolenic Acid logistics, can safely deploy.
They are entering a thoroughly pacified combat zone, completely free of oxidative shrapnel and inflammatory debris.
The secondary wave of [The Neuro-Endocrine Storm] is permanently silenced, and your deeply vulnerable dermal architecture remains flawlessly, beautifully intact.

2.4 THE COLLAGEN GUARDIAN:
CONFISCATING THE MMP MOLECULAR SCISSORS
Preventing the enzymatic degradation of the dermal extracellular matrix.
The ultimate tragedy of continuous photo-aging is not simply the immediate death of your surface cells. It is a far more insidious, horrifying biological betrayal.
Under the crushing weight of relentless ultraviolet radiation, your surviving cells are actively hijacked by the environment. They are biologically brainwashed into completely destroying their own meticulously built home.
The [The Neuro-Endocrine Storm] forces your own exhausted dermal fibroblasts to mutate into highly efficient, collagen-shredding machines.

1. THE UV-INDUCED ENZYME SURGE
The release of Matrix Metalloproteinases.
We must forensically examine this exact mechanism of microscopic self-destruction.
It is this invisible, relentless enzymatic process that directly translates into the deep, irreversible crevices you eventually see in the mirror.
I. The Oxidative Trigger:
The destructive sequence begins with the chaotic secondary wave of oxidative shrapnel we previously identified.
When Reactive Oxygen Species flood the deep dermal matrix, they do not just cause direct physical damage to the fragile lipid walls.
They act as highly toxic, extremely potent messenger molecules that hack your cellular communication network. These volatile free radicals penetrate deep into the protected nucleus of your surviving dermal fibroblasts.
They aggressively trigger a catastrophic, irreversible genetic panic response. The sheer oxidative stress violently forces the cell’s DNA to upregulate the mass production of destructive proteins.
It is a false alarm that commands the cell to tear down its own structural foundation.

II. MMP-1 and MMP-3:
This triggered biological swarm is primarily composed of Matrix Metalloproteinases, commonly known to scientists as MMPs.
These are not passive, harmless molecules floating in the background; they are highly aggressive, biological molecular scissors. They are specifically engineered by your own biology to cut, tear, and completely dissolve the structural proteins of your skin.
MMP-1 is the primary, devastating destroyer of Type I collagen.
Type I collagen is the thick, rigid, essential scaffolding that keeps your face lifted, firm, and structurally sound. As MMP-1 violently floods the extracellular matrix, it systematically chops this vital collagen into useless, fragmented cellular debris.
Simultaneously, the panicked cell releases a massive wave of MMP-3, an equally devastating and highly specialized enzyme.
MMP-3 specifically targets and degrades your precious elastin. Elastin is the delicate, rubber-band-like protein network that allows your skin to rapidly snap back into place after making a facial expression.
When you pull at your cheek and the skin sluggishly, slowly returns to its original position, you are feeling the direct consequence of MMP-3. Your biological elasticity has been chemically shredded from the inside out.
This dual-enzyme surge is the exact, undeniable biological root cause of premature sagging, deep structural wrinkles, and severe tissue collapse.

2. THE ASTAXANTHIN BLOCKADE
Shutting down the enzymatic shredders.
The primitive, mass-market skincare industry attempts to solve this profound structural collapse by rubbing heavy, synthetic collagen molecules on the dead surface of the skin.
This is mathematically and biologically useless.
You cannot rebuild a burning house while the arsonists are still actively inside wielding flamethrowers.
To permanently save the dermal matrix, the Commander must completely and physically confiscate the molecular scissors.
First, Transcriptional Suppression:
Astaxanthin executes a profound, highly coordinated genetic intervention long before the destructive enzymes are ever even created.
Because [The Systemic Commander] perfectly neutralizes the massive influx of free radicals at the membrane level, the toxic signal never reaches the nucleus. The genetic panic response is entirely aborted before it can begin.
Astaxanthin aggressively intercepts and silences the upstream signaling pathways, specifically the MAPK and AP-1 genetic cascades. It physically prevents the fibroblast’s DNA from ever receiving the chemical order to manufacture the destructive MMPs in the first place.
This is not mere damage control; this is absolute, preemptive biological dominance.
Second, Direct Enzyme Inhibition:
But what about the rogue enzymatic shredders that have already been released into the extracellular matrix from previous hours of unprotected exposure?
The Commander does not leave the existing, vulnerable collagen structure undefended for a single microsecond. Astaxanthin actively downregulates the localized activation of the rogue MMPs currently floating within the dermal environment.
-
It profoundly alters the local biochemical climate, making it incredibly hostile and difficult for these molecular scissors to function.
-
It physically impedes their mechanical ability to latch onto and cleave the healthy, intact collagen and elastin fibers.
The destructive swarm is effectively paralyzed and neutralized directly on the battlefield.
Third, The Structural Preservation:
We must fully understand the sheer architectural magnitude of this flawless dual-action blockade.
By completely suppressing the genetic production of new enzymes and actively paralyzing the existing ones, Astaxanthin achieves absolute structural preservation.
-
The thick, vital Type I collagen scaffolding remains perfectly intact, absolutely refusing to buckle under the relentless weight of gravity.
-
The delicate, complex elastin network retains its absolute tensile strength and rapid, youthful snap-back capability.
Your skin is no longer a localized, tragic warzone of active biological self-destruction.
The biological arsonists have been completely and permanently expelled from the building.
This pristine, enzymatically quiet, and structurally sound environment is mathematically non-negotiable for the Keyora 1+1+1+1+1+1+1 > 7 protocol.
Without the Commander actively halting the MMP shredders, the essential lipid vanguard would be deploying into a collapsing, dissolving void.
-
The Linoleic Acid mortar cannot successfully rebuild the moisture barrier if the underlying collagen floor is actively giving way beneath it.
-
The Alpha-Linolenic Acid logistics network cannot construct new, life-saving micro-vessels if the elastin support struts are chemically dissolving.
And the Oleic Acid spacer cannot stabilize the cellular architecture if the entire extracellular matrix is being chopped into debris.
The Commander completely secures the absolute structural integrity of the deep extracellular matrix.
It guarantees the sensitive biological architects an undeniably stable, perfectly preserved foundation to rebuild upon.

2.5 THE INFLAMMATORY LOCKDOWN:
SILENCING NF-κB AND COX-2
Halting the acute erythema and chronic inflammatory decay triggered by solar radiation.
Where there is microscopic oxidative fire, there is always macroscopic inflammatory smoke.
You know this smoke intimately as the painful, hot, and radiating redness that completely takes over your face after a long, unprotected day in the sun.
This is not a simple, passive physical reaction to the environmental heat of the afternoon. It is the undeniable, visual evidence of a massive, systemic immune panic actively tearing through your dermal layers.
Ultraviolet radiation triggers this chaotic immune cascade, leading directly to acute swelling, agonizing sensitivity, and chronic, irreversible tissue degradation.

PHASE 1: THE INFLAMMATORY ALARM
The activation of the NF-κB pathway.
Before the visible redness even appears on the physical surface of your skin, a silent, catastrophic biological alarm is tripped deep within the cellular nucleus.
– The UV Trigger:
The initial, violent wave of radiation damage we explored earlier forcefully activates a highly sensitive transcription factor known to biologists as NF-κB.
Think of NF-κB as the absolute, non-negotiable master switch for the localized immune system inside your dermal fibroblasts and keratinocytes. Under normal, healthy, non-irradiated conditions, this master switch rests quietly, firmly locked down in the cellular cytoplasm, completely deactivated.
But the sheer kinetic force of the solar strike and the resulting oxidative stress shatters its chemical lock, setting it completely free.
This is the exact microscopic ignition point of [The Neuro-Endocrine Storm] within the skin.
– The Cytokine Release:
Once freed from its structural restraints, the panicked NF-κB molecule rapidly translocates.
It forcefully pushes its way directly into the highly protected control room of the cell, the central nucleus. Once inside, it binds aggressively to your DNA and screams for an immediate, overwhelmingly violent biological response to the perceived radiation threat.
It commands the cell to begin the mass production and immediate release of highly destructive, pro-inflammatory cytokines, specifically IL-1β, IL-6, and TNF-α.
These cytokines are toxic, signaling flare guns that saturate the extracellular matrix, loudly broadcasting the localized biological disaster to the rest of your immune system.

PHASE 2: THE COX-2 AMPLIFICATION
The synthesis of prostaglandins.
The biological crisis immediately deepens as the initial alarm is exponentially magnified by the surrounding tissues.
– The Enzyme Upregulation:
This screaming chorus of inflammatory cytokines does not just dissipate harmlessly into the ether of the extracellular fluid. It directly and aggressively upregulates the rapid synthesis of a secondary, highly dangerous enzyme called Cyclooxygenase-2, or COX-2.
COX-2 is not an enzyme your skin requires for daily, healthy survival, hydration, or basic structural maintenance.
It is an emergency-only, highly destructive biological amplifier.
Its sole, devastating purpose in this irradiated environment is to take the initial, localized panic and multiply it across the entire dermal landscape.
– The Erythema Response:
Once COX-2 heavily floods the cellular environment, it immediately begins converting your precious structural fatty acids into Prostaglandin E2 (PGE2).
PGE2 is the exact, undeniable chemical culprit behind the agonizing physical symptoms of your severe sunburn.
It forces the microscopic blood vessels in your skin to violently dilate, swell, and engorge far beyond their natural biological capacity.
This extreme vasodilation is the exact mechanical reason why your skin turns bright, angry red and radiates that intense, uncomfortable, and throbbing heat.
The PGE2 binds directly to the localized nerve endings, transforming a microscopic chemical reaction into the very real, visceral sensation of burning pain.

PHASE 3: THE ASTAXANTHIN SILENCING
The biological override.
The primitive, mass-market skincare industry attempts to soothe this raging biological fire by applying superficial, cooling botanical gels to the dead surface of the skin.
This is a mathematically useless gesture of temporary physical comfort that entirely ignores the cellular disaster actively multiplying in the tissue beneath.
You cannot permanently stop a five-alarm chemical fire with a thin, temporary layer of aloe vera.
To truly save the dermal matrix from chronic decay and structural collapse, [The Systemic Commander] must execute a total, uncompromising biological override.
Astaxanthin does not wait for the painful redness and swelling to physically manifest on the surface. It strikes directly at the very root of the inflammatory cascade, long before the first toxic cytokine is ever even manufactured.
Because Astaxanthin is permanently locked across the entire 30-Angstrom span of the cellular membrane, it aggressively intercepts the initial oxidative panic signals.
-
It exerts a profound, clinically proven suppressive force directly on the NF-κB transcription factor.
-
It physically reinforces the chemical restraints holding NF-κB in the cytoplasm, absolutely refusing to let the master switch break free.
By permanently locking this genetic switch in the “off” position, the Commander prevents the molecule from ever entering the nucleus.
Your DNA never receives the false, panic-inducing command to self-destruct. The toxic, screaming chorus of IL-1β, IL-6, and TNF-α cytokines is completely and utterly silenced before a single molecule can be synthesized.
Without that critical, upstream cytokine signal, the destructive COX-2 enzyme is never upregulated by the surrounding tissue.
Because COX-2 is neutralized, the agony-inducing PGE2 is never produced from your structural lipids.
Your localized blood vessels remain calm, structurally tight, and perfectly regulated, entirely preventing the painful, visually damaging vasodilation. The acute erythema, the severe redness, and the deep tissue swelling are completely halted at the absolute deepest genetic level.
This profound, systemic inflammatory lockdown is the non-negotiable prerequisite for the entire Keyora cellular reconstruction protocol. A biological architect cannot successfully rebuild a complex, delicate structure while the site is actively burning and swarming with panicked immune cells.
By forcing this absolute state of localized chemical peace, the Commander clears the battlefield entirely. It mathematically guarantees that the incoming essential fatty acids are not instantly consumed by the inflammatory fire.
The essential lipid vanguard can finally deploy into a quiet, perfectly stabilized environment, ready to execute their precise, life-saving bio-architectural repairs.

2.6 THE PIGMENT CONTROLLER:
REGULATING MELANOGENESIS AT THE SOURCE
Non-destructive modulation of melanin synthesis to prevent photo-induced hyperpigmentation and age spots.
The skin’s desperate, biological attempt to shield itself from continuous ultraviolet radiation results in a panicked, chaotic overproduction of melanin.
You know the exact, devastating visual consequence of this deep cellular panic when you look in the mirror months later. It is the sudden, irreversible appearance of those dark, jagged, and uneven patches of pigmentation that permanently stain the surface of your face.
These are not just superficial cosmetic blemishes that can be easily covered up or washed away.
They are the dark, highly visible, and stubborn scars of deep cellular trauma, localized oxidative stress, and surviving age spots.
They represent a total failure of the skin’s internal regulatory systems under the overwhelming weight of environmental radiation.
The Commander must step in to forcefully regulate this absolute biological panic before the damage becomes a permanent, structural stain on your complexion.
PROPOSITION B:
How does Astaxanthin prevent hyperpigmentation without destroying melanocytes?

The Evidence:
The mass-market beauty industry attempts to solve this profound biological failure using a crude, scientifically dangerous method.
They attempt to erase these dark stains by aggressively applying harsh, toxic chemical bleaching agents directly onto the skin barrier.
These aggressive synthetic acids, such as high-concentration hydroquinone, do not just politely ask the cell to stop pigment production.
They actively poison the tissue and literally destroy the living melanocyte cells in a desperate attempt to lighten the skin.
This primitive, scorched-earth tactic permanently damages the deeper tissue, triggers severe secondary inflammation, and completely ruins the skin’s natural baseline defense.
Clinical evidence dictates a far superior, non-destructive, and mathematically sound approach through the deployment of [The Systemic Commander].
Unlike these harsh, cell-killing chemicals, Astaxanthin successfully regulates pigment production at the absolute deepest genetic level.
It actively prevents hyperpigmentation and age spots without causing a single cellular casualty or compromising the structural integrity of the surrounding tissue.
1. MITF Downregulation:
Deep within the heavily protected nucleus of your melanocytes lies a highly specific, extremely sensitive master genetic switch.
This switch is known to molecular biologists as the Microphthalmia-associated transcription factor, or simply MITF. MITF is the absolute, undisputed genetic commander of your skin’s entire microscopic pigmentation factory.
When ultraviolet radiation aggressively strikes the dermal matrix, it violently forces this MITF switch into a state of chaotic overdrive. It screams at the cell’s DNA, demanding an emergency, maximum-capacity production of melanin to shield the burning tissue.
Astaxanthin actively and decisively suppresses this panicked MITF transcription factor.
It forcefully pulls the master switch back down to its normal, healthy baseline resting state.
It chemically tells the cell’s DNA to immediately halt the emergency mass-production order, stopping the chaotic pigment surge before it can even begin.
2. Tyrosinase Inhibition:
However, biological systems are complex, and even if some of the panicked genetic commands slip through the initial blockade, the defense does not fail.
The actual physical manufacturing and assembly of melanin molecules requires the continuous labor of a specific, hard-working enzyme called Tyrosinase. Without Tyrosinase actively synthesizing the raw biochemical materials, the dark pigment literally cannot be physically built.
Astaxanthin directly targets and significantly reduces the biological activity of this localized Tyrosinase enzyme within the melanocyte.
It actively slows down the internal microscopic assembly line.
It mathematically ensures that even if the cellular factory remains open, it simply cannot produce the chaotic, overwhelming volume of dark pigment demanded by the radiation.
This flawless dual-action genetic and enzymatic blockade is absolute, mathematically precise, yet completely and entirely non-toxic to the living cell.

The Mechanism:
We must now forensically examine exactly how the Commander executes this profound genetic and enzymatic regulation from a biophysical standpoint.
It is not a magic trick; it all leads directly back to the absolute suppression of the localized oxidative fire we established in the previous sections.
The entire pigmentation cascade is fundamentally driven by oxidative stress, making Astaxanthin the ultimate biological off-switch.
I. The Oxidative Signal:
Melanocytes do not just spontaneously decide to ramp up melanin production for no biological reason; they are highly reactive, sensitive cellular machinery.
They are reacting to a very specific, toxic chemical trigger that is actively flooding the surrounding extracellular matrix.
They strictly respond to the massive influx of Reactive Oxygen Species, the exact localized oxidative shrapnel we previously identified as [The Neuro-Endocrine Storm].
This aggressive ROS acts as a loud, terrifying chemical alarm, tricking the melanocyte into believing the entire surrounding tissue is actively dying. It forces the melanocyte to deploy an immediate, excessively heavy melanin shield to protect the surviving cellular structures.
But by ruthlessly and systematically quenching this localized ROS using its massive conjugated polyene chain, Astaxanthin completely removes the toxic panic signal.
The loud, screaming chemical alarm is entirely silenced before the melanocyte even has a chance to physically react.
II. The Controlled Output:
Because the oxidative alarm has been successfully and completely muted, the melanocyte instantly drops its panicked, defensive posture.
It biologically realizes the surrounding environment is no longer a hostile, highly irradiated, and burning warzone. The melanocytes remain completely healthy, structurally intact, and fully alive, functioning exactly as nature intended.
But their microscopic melanin output is beautifully, safely, and predictably normalized. The cell goes back to producing just enough standard pigment to maintain your natural, healthy, baseline complexion.
It entirely prevents the dark, uneven, panicked clumping of concentrated melanin that eventually surfaces and hardens into stubborn, highly visible age spots.

The Resolution:
This is the ultimate, non-destructive biophysical victory over continuous photo-induced hyperpigmentation.
Astaxanthin provides a completely safe, highly regulated, and strictly endogenous method for maintaining a luminous, perfectly even skin tone under severe environmental stress.
It permanently protects the delicate, essential melanocytes from the toxic destruction of chemical bleaching while strictly enforcing localized biological discipline.
It mathematically ensures the Keyora 1+1+1+1+1+1+1 > 7 protocol operates in a perfectly balanced, aesthetically pristine, and enzymatically calm dermal matrix.
The Commander has successfully and permanently controlled the pigment at the source.
It preserves both the deep biological health and the flawless, radiant visual integrity of your cellular architecture, allowing you to walk through the sun without the fear of permanent staining.

2.7 THE MATRIX GUARDIAN:
PROVIDING COVER FIRE FOR THE LIPID VANGUARD
Why the endogenous reconstruction of ceramides and vascular networks by Omega-3/6/9 is mathematically impossible without Astaxanthin.
A military commander, no matter how physically unbreakable, cannot win a war without the specialized logistics and engineering battalions operating safely behind the front lines.
Astaxanthin’s ultimate, defining biological purpose is not merely to stand blindly in the fire and absorb radiation for the sake of its own survival. Its true function is to provide absolute, uncompromising thermodynamic cover fire so the Bioactive Carrier can safely deploy its fragile, highly sensitive lipid payload.
We have successfully established the impenetrable defensive perimeter; now we must systematically bring in the biological architects to rebuild the shattered matrix.
But these specific lipid builders are fundamentally vulnerable, and deploying them without the Commander is a mathematically guaranteed biological suicide mission.

1. THE VULNERABILITY OF THE BUILDERS
The danger of unprotected fatty acids.
We must forensically examine the exact chemical nature of the Bioactive Carrier that Keyora strictly uses to rebuild your failing cellular infrastructure.
– The Double-Bond Target:
The essential lipids required to reconstruct your failing skin, specifically Alpha-Linolenic Acid (ALA), Linoleic Acid (LA), and Oleic Acid (OA), are structurally polyunsaturated.
This specific chemical classification dictates that their incredibly long carbon chains contain multiple, highly reactive double bonds. While these exact double bonds give the lipids the crucial flexibility required to build healthy, fluid cell membranes, they are also massive biological liabilities.
To a volatile, hyper-excited free radical roaming the deeply irradiated layers of your dermis, these polyunsaturated double bonds are the ultimate, highly visible targets. They are precisely the exact chemical structures that UV-induced singlet oxygen desperately wants to violently attack, cleave, and completely rip apart.
You cannot send unprotected, highly vulnerable builder lipids into an active, burning warzone and expect them to survive the initial biochemical impact.
– The Oxidized Hazard:
If you consume a standard, unprotected mass-market fish oil or basic plant lipid capsule before walking into the afternoon sun, you are making a catastrophic error.
Without the absolute thermodynamic cover fire of the Commander, these fragile healing lipids are instantly targeted the exact microsecond they enter the irradiated dermis.
They do not get a single chance to synthesize essential ceramides, hydrate the tissue, or rebuild the collapsed vascular networks.
They are immediately oxidized, structurally shattered, and violently mutated into a highly toxic, tissue-destroying biological byproduct known directly as malondialdehyde (MDA).
Instead of healing your fragile skin, these unprotected builder lipids literally become the localized oxidative stress.
They actively accelerate the deep degradation of your dermal matrix, making the structural collapse significantly worse than if you had taken nothing at all.

2. THE COVER FIRE SYNERGY (1+1+1+1+1+1+1 > 7)
The interdependent biological architecture.
This exact structural vulnerability is the precise reason why the Keyora bio-architectural law dictates that standard, single-ingredient supplementation is a mathematical failure.
You cannot just haphazardly take Omega fatty acids and hope for deep biological reconstruction.
They must be strategically deployed under the absolute, unbroken thermodynamic shield of [The Systemic Commander].
I. Protecting LA (The Mortar):
Linoleic Acid (LA) is the critical, non-negotiable biological mortar strictly required to synthesize O-acylceramides and physically seal the microscopic cracks in the stratum corneum.
If LA is violently oxidized by ultraviolet radiation, your skin immediately loses its fundamental ability to hold water. This results in the severe, painful, and chronic dehydration that makes your face feel like dry, brittle paper at the end of the day.
Astaxanthin actively traps the deep-membrane lipid peroxyl radicals, providing the exact, flawless cover fire LA needs to safely travel through the volatile cellular layers.
Because the Commander is actively absorbing the kinetic radiation strikes, the LA mortar remains perfectly, structurally intact.
It successfully reaches the upper epidermis to seal the barrier, ensuring the structural foundation is permanently secured against trans-epidermal water loss.
II. Protecting OA (The Spacer):
Oleic Acid (OA) is the essential, heavily stabilizing biological spacer that strictly maintains the highly fluid, “Liquid-Crystal” state of the cellular membrane.
It is also the critical metabolic trigger required to activate advanced AMPK pathways, which aggressively pacify panicked glial and immune cells in the surrounding tissue.
If OA is compromised and shattered by oxidative shrapnel, the cellular membrane immediately becomes stiff, brittle, rigid, and highly prone to mechanical rupture.
The Commander’s massive 30-Angstrom structural rivet physically prevents the violent lipid peroxidation cascade that would otherwise completely destroy this critical Omega-9 fatty acid.
Safely operating under this impenetrable thermodynamic shield, OA can properly stabilize the three-dimensional membrane architecture.
It silences the localized inflammatory alarm, allowing the surrounding tissue to safely return to a state of absolute, calm metabolic regulation.
III. Protecting ALA (The Logistics):
Alpha-Linolenic Acid (ALA) is the supreme logistical commander of the entire lipid network, responsible for generating the heaviest, most critical repair molecules in the body.
-
It must safely travel through the highly volatile bloodstream to the liver to successfully convert into EPA, which is required to extinguish deep systemic inflammation.
-
It must also survive long enough to synthesize DPA, the exact molecule required to rebuild the collapsed dermal micro-vessels via VEGF up-regulation.
-
Finally, it must successfully generate DHA to heavily reinforce the deep, structural baseline of your central nervous system and vulnerable cellular membranes.
Because Astaxanthin entirely neutralizes the chaotic [The Neuro-Endocrine Storm] of free radicals, the ALA logistics network is completely protected from oxidative destruction.
The entire fragile sequence of EPA, DPA, and DHA conversion can proceed flawlessly, delivering the ultimate biological payload exactly where the tissue is failing.

3. THE ABSOLUTE LAW OF THE MATRIX
The impossibility of isolated survival.
We must now firmly accept the absolute, mathematically verifiable law of the deep cellular matrix under conditions of severe environmental radiation stress.
Absolutely none of these highly advanced, life-saving lipid molecules can survive or function alone in a violently UV-irradiated dermal environment.
If you foolishly deploy the LA mortar without the Astaxanthin shield, the mortar instantly burns and turns into highly toxic biological ash.
If you attempt to deploy the ALA logistics network without the absolute cover fire, the delicate supply lines are instantly severed by oxidative shrapnel.
The biological matrix MUST be deployed simultaneously, operating strictly as a single, perfectly coordinated, heavily armored biological strike force.
This is the exact, uncompromising biophysical logic behind the Keyora 1+1+1+1+1+1+1 > 7 synergy protocol.
The individual structural components are completely meaningless if they are violently destroyed before they can execute their specific bio-architectural programming.
The Commander is the absolute, non-negotiable thermodynamic anchor that makes the entire cellular reconstruction process physically, mathematically, and biologically possible.

CONCLUSION: THE FOUNDATION IS SECURED
Transitioning from the thermodynamic defense of Astaxanthin to the bio-architectural reconstruction by the endogenous lipids.
The singlet oxygen bombs have been entirely defused.
The destructive collagen-shredding enzymes have been permanently locked down.
The fragile cellular membrane is now completely and thermodynamically secure.
[The Systemic Commander] has held the line against the relentless invisible assault of the sun.
We have successfully stabilized the localized [The Neuro-Endocrine Storm], but mere survival is not our ultimate biological goal.

I. THE DEFEAT OF THE PHOTON
The triumph of physical quenching.
We must pause and acknowledge the sheer biophysical magnitude of this exact thermodynamic victory.
For decades, the beauty industry fed you a mathematically broken lie of chemical suicide. They told you to rely on fragile, water-soluble vitamins that died the microsecond they encountered a free radical.
We have permanently replaced that tragic failure with absolute structural supremacy.
By embedding its precise 30-Angstrom carbon geometry directly across your cellular lipid bilayer, Astaxanthin rewrote the rules of engagement.
-
It intercepted the catastrophic kinetic energy of the ultraviolet photon without ever breaking its own bonds.
-
It trapped the volatile excitation energy in its massive polyene chain.
-
It then utilized the flawless mechanics of vibrational cooling to safely bleed that lethal radiation away as microscopic heat.
-
It systematically neutralized the secondary ROS fallout, silencing the NF-kB inflammatory alarm and protecting your precious melanocytes from panic.
-
It achieved all of this without donating a single electron, remaining completely structurally unbroken.
The photon has been mathematically and biologically defeated.

II. ENTER THE ARCHITECTS
The deployment of the lipid vanguard.
However, establishing an impenetrable defense perimeter is only the very first phase of the Keyora biological protocol.
The Commander has successfully cleared the microscopic battlefield of oxidative shrapnel. But we are still intimately left with the devastating, physical aftermath of the previous cellular damage.
You can still feel the lingering structural exhaustion.
Your critical moisture barrier has evaporated into the dry atmosphere, leaving the tissue brittle and highly compromised.
The delicate dermal micro-vessels that feed oxygen and nutrients to your fibroblasts have completely collapsed under the weight of the earlier oxidative stress.
Now that the thermodynamic cover fire is absolute, and the environment is finally enzymatically quiet, we must begin the delicate work of deep reconstruction.
We must now systematically deploy the highly specialized biological architects. This is the precise, critical moment the essential lipid vanguard of the Bioactive Carrier safely enters the secured combat zone.
We will fundamentally transition from the raw, unbroken physics of Astaxanthin’s defense to the intricate, life-saving chemistry of cellular rebuilding.
In the next phase, we will zoom in with absolute forensic precision on the specific, non-negotiable roles of these advanced Omega fatty acids.
We will witness exactly how they utilize this mathematically perfect, fully protected environment to structurally restore your biological youth.
We will start by deploying the Epidermal Mortar, Linoleic Acid, to permanently seal the fractured stratum corneum.
Then, we will introduce the Liquid-Crystal Spacer, Oleic Acid, to restore total mechanical fluidity and metabolic peace to the cellular membrane.
The foundation is finally, undeniably secure.
The biological architects are ready to build.

References:
Fisher, G. J., Datta, S. C., Talwar, H. S., Wang, Z. Q., Varani, J., Kang, S., & Voorhees, J. J. (1996). Molecular basis of sun-induced premature skin ageing and retinoid antagonism. Nature, 379(6563), 335-339.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta (BBA)-Biomembranes, 1512(2), 251-258.
McNulty, H. P., Byun, J., Lockwood, S. F., Jacob, R. F., & Mason, R. P. (2007). Differential effects of carotenoids on lipid peroxidation due to membrane interactions: X-ray diffraction analysis. Biochimica et Biophysica Acta (BBA)-Biomembranes, 1768(1), 167-174.
Camera, E., Mastrofrancesco, A., Fabbri, C., Daubrawa, F., Picardo, M., Sies, H., & Stahl, W. (2009). Astaxanthin, canthaxanthin and beta-carotene differently affect UVA-induced oxidative damage and expression of oxidative stress-responsive enzymes. Experimental Dermatology, 18(3), 222-231.
Suganuma, K., Nakajima, H., Ohtsuki, M., & Imokawa, G. (2010). Astaxanthin attenuates the UVA-induced up-regulation of matrix-metalloproteinase-1 and skin fibroblast elastase in human dermal fibroblasts. Journal of Dermatological Science, 58(2), 136-142.
Lee, S. J., Bai, S. K., Lee, K. S., Namkoong, S., Na, H. J., Ha, K. S., Han, J. A., Yim, S. V., Choi, K. B., … & Kim, Y. M. (2003). Astaxanthin inhibits nitric oxide production and inflammatory gene expression by suppressing I(kappa)B kinase-dependent NF-kappaB activation. Molecules and Cells, 16(1), 97-105.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Bickers, D. R., & Athar, M. (2006). Oxidative stress in the pathogenesis of skin disease. Journal of Investigative Dermatology, 126(12), 2565-2575.
Rittie, L., & Fisher, G. J. (2002). UV-light-induced signal cascades and skin aging. Ageing Research Reviews, 1(4), 705-720.
Beutner, S., Bloedorn, B., Frixel, S., Blanco, I., Hoffmann, T., Martin, H. D., … & Schalch, W. (2001). Quantitative assessment of antioxidant properties of natural colorants and phytochemicals: carotenoids, flavonoids, phenols and indigoids. The role of beta-carotene in antioxidant functions. Journal of the Science of Food and Agriculture, 81(6), 559-568.
Palozza, P., & Krinsky, N. I. (1992). Astaxanthin and canthaxanthin are potent antioxidants in a liposomal model. Archives of Biochemistry and Biophysics, 297(2), 291-295.
Tominaga, K., Hongo, N., Karato, M., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43-47.
Ito, N., Seki, S., & Ueda, F. (2018). The protective role of astaxanthin for UV-induced skin deterioration in healthy people-A randomized, double-blind, placebo-controlled trial. Nutrients, 10(7), 817.
Fakhri, S., Abbaszadeh, F., Dargahi, L., & Jorjani, M. (2018). Astaxanthin: A mechanistic review on its biological activities and health benefits. Pharmacological Research, 136, 1-20.
D’Orazio, J., Jarrett, S., Amaro-Ortiz, A., & Scott, T. (2013). UV radiation and the skin. International Journal of Molecular Sciences, 14(6), 12222-12248.
Slominski, A., Tobin, D. J., Shibahara, S., & Wortsman, J. (2004). Melanin pigmentation in mammalian skin and its hormonal regulation. Physiological Reviews, 84(4), 1155-1228.
Ambati, R. R., Phang, S. M., Ravi, S., & Aswathanarayana, R. G. (2014). Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications-A review. Marine Drugs, 12(1), 128-152.
Fassett, R. G., & Coombes, J. S. (2011). Astaxanthin: A potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447-465.
Yuan, J. P., Peng, J., Yin, K., & Wang, J. H. (2011). Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Molecular Nutrition & Food Research, 55(1), 150-165.
Guerin, M., Huntley, M. E., & Olaizola, M. (2003). Astaxanthin from Haematococcus pluvialis: Implications for human health and nutrition. Trends in Biotechnology, 21(5), 210-216.
Suzuki, Y., Shibata, A., Kawakami, Y., & Koumoto, M. (2006). Suppressive effects of astaxanthin against ozone-induced airway inflammation in mice. Journal of Agricultural and Food Chemistry, 54(8), 3041-3045.
Santocono, M., Zurria, M., Berrettini, M., Fedeli, D., & Falcioni, G. (2006). Lutein, zeaxanthin and astaxanthin protect against DNA damage in SK-N-SH human neuroblastoma cells induced by reactive nitrogen species. Journal of Photochemistry and Photobiology B: Biology, 85(3), 205-215.
Wolf, A. M., Asoh, S., Hiranuma, H., Ohsawa, I., Iio, K., Wakui, A., … & Ohta, S. (2010). Astaxanthin protects mitochondrial redox state and functional integrity against oxidative stress. Journal of Nutritional Biochemistry, 21(5), 381-389.
Higuera-Ciapara, I., Felix-Valenzuela, L., & Goycoolea, F. M. (2006). Astaxanthin: A review of its chemistry and applications. Critical Reviews in Food Science and Nutrition, 46(2), 185-196.
Biesalski, H. K., Hemmes, C., Hopfenmuller, W., Schmid, C., & Gollnick, H. P. (1995). UV light, beta-carotene and human skin-beneficial and potentially harmful effects. Archives of Dermatological Research, 287(6), 549-554.
Martin, H. D., Jager, C., Ruck, C., Schmidt, M., Walenciak, R., & Weis, M. (1999). Anti- and prooxidant properties of carotenoids. Journal of Praktische Chemie, 341(3), 302-308.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Nishigaki, I., Rajendran, P., Venugopal, R., Ekambaram, G., Sakthisekaran, D., & Nishigaki, Y. (1994). Suppressive effect of astaxanthin on lipid peroxidation induced in rats. Journal of Clinical Biochemistry and Nutrition, 16(3), 161-166.
Lorenz, R. T., & Cysewski, G. R. (2000). Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends in Biotechnology, 18(4), 160-167.
Yamashita, E. (2002). Cosmetic benefit of dietary supplements containing astaxanthin and tocotrienol on human skin. Food Style, 21(6), 112-117.
Scharffetter-Kochanek, K., Brenneisen, P., Wenk, J., Herrmann, G., Eddie, W., … & Krieg, T. (2000). Photoaging of the skin from phenotype to mechanisms. Experimental Gerontology, 35(3), 307-316.
Shiratori, K., Ohgami, K., Ilieva, I., Jin, X. H., Shirakawa, K., … & Ohno, S. (2005). Effects of astaxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Investigative Ophthalmology & Visual Science, 46(6), 2120-2126.

# Knowledge Summary of Chapter 2: The Thermodynamic Heat-Sink
## I. THE THERMODYNAMIC BASELINE [THE ENVIRONMENTAL TAX & CHEMICAL SUICIDE]
* **The Physics of Radiation:** The dermal matrix endures a continuous, 11-hour kinetic strike of high-velocity ultraviolet (UVA/UVB) photons.
* **The Inefficiency:** Traditional water-soluble vitamins (e.g., Vitamin C) rely on a fragile, mathematically flawed “electron donation” mechanism.
* **The Reaction:** UV Photons + Structural Lipids -> Ejected Electrons -> **Singlet Oxygen** & highly reactive Free Radicals.
* **The Consequence:** The finite electrons of traditional antioxidants are instantly depleted, triggering a “chemical suicide” where they transform into pro-oxidant shrapnel. The defense collapses entirely, igniting localized **[The Neuro-Endocrine Storm]**.
## II. THE STRUCTURAL DEFENSE [THE 30-ANGSTROM ANCHOR]
* **The Target:** **Cellular Lipid Bilayer** (Deep Dermal Fibroblasts & Basal Keratinocytes).
* **The Threat:** Unchecked lipid peroxidation -> Membrane rupture -> Cytoskeleton collapse and irreversible trans-epidermal water loss.
* **The Mechanism:** **[The Systemic Commander]**.
* Astaxanthin’s hydrophilic polar heads magnetically lock into the aqueous boundaries on both the inside and outside of the cell.
* The hydrophobic conjugated polyene chain precisely spans the exact **30-Ångström** width of the lipid core.
* This forms a permanent, vertical physical rivet, entirely securing membrane fluidity and preventing mechanical shearing.
* **The Evidence:** Constructs an absolute dual-sided shield, simultaneously intercepting external UV-induced ROS and internal mitochondrial-leaked ROS.
## III. THE BIOPHYSICAL QUENCHING [THE SINGLET OXYGEN TRAP]
* **The Target:** The highly volatile **Singlet Oxygen Bomb** carrying lethal quantum excitation energy.
* **The Threat:** The pure kinetic energy instantly cleaves the fragile double bonds of polyunsaturated fatty acids, liquefying tissue and causing deep inflammatory heat.
* **The Mechanism:** **[Vibrational Cooling]**.
* Astaxanthin absolutely does NOT donate electrons. Its massive carbon backbone acts as an electromagnetic net, directly absorbing the pure quantum excitation energy.
* The molecule undergoes violent microscopic resonance (biological tuning fork effect), mechanically converting the lethal radiation into **harmless microscopic heat** through continuous friction with surrounding cellular fluid.
* **The Consequence:** The Astaxanthin molecule remains structurally unbroken (0 electron loss, 0 oxidation). Its biochemical capacity to quench singlet oxygen is mathematically **6,000x stronger** than Vitamin C, allowing for limitless defensive cycling.

## IV. THE BIOLOGICAL OVERRIDE [ENZYMATIC & INFLAMMATORY LOCKDOWN]
* **The Target:** DNA Transcription Factors (NF-κB, MITF) and Destructive Enzymes (MMPs, COX-2, Tyrosinase).
* **The Threat:** ROS acts as a toxic alarm, hijacking cellular communication and commanding self-destruction:
* **MMP-1 & MMP-3:** Shreds Type I Collagen and elastin (causing deep wrinkles and sagging).
* **NF-κB Pathway:** Releases cytokines -> Upregulates **COX-2** -> Synthesizes **PGE2** (causing agonizing vasodilation and acute erythema).
* **MITF Factor:** Triggers melanocyte panic -> Melanin overproduction (causing permanent age spots).
* **The Mechanism:** Astaxanthin completely intercepts the upstream oxidative alarm at the membrane level, physically locking NF-κB in the cytoplasm and paralyzing existing MMP cleavage activity.
* **The Consequence:** Perfect preservation of the collagen matrix and absolute suppression of the inflammatory network, achieving true biochemical silence without killing a single living melanocyte.
## V. THE FUEL MATRIX [PROTECTED SYNERGY]
* **The Philosophy:** No fragile lipid builder can survive in an irradiated combat zone without absolute thermodynamic cover fire.
* **The Components:**
1. **The Mortar:** **Linoleic Acid (LA / Omega-6)**. Synthesizes O-acylceramides to permanently seal the stratum corneum and stop water loss.
2. **The Spacer:** **Oleic Acid (OA / Omega-9)**. Maintains the liquid-crystal state of the membrane and activates AMPK to pacify immune cells.
3. **The Logistics:** **Alpha-Linolenic Acid (ALA / Omega-3)**. Systemically converts into EPA (extinguishing deep inflammation), DPA (rebuilding micro-vessels), and DHA (reinforcing the neural/membrane baseline).
* **The Synergy:** Astaxanthin provides the absolute cover fire **(1+1+1+1+1+1+1 > 7)**, preventing these polyunsaturated fatty acids from being oxidized into toxic **Malondialdehyde (MDA)** upon entering the dermis, ensuring the flawless execution of Keyora’s bio-architectural reconstruction.

Chapter 3: The Mortar of the Epidermis:
Sealing the Barrier with Linoleic Acid
How the Bioactive Carrier delivers the essential Omega-6 precursor to synthesize O-acylceramides, locking in moisture under the absolute protection of Astaxanthin.
You know the exact, maddening cycle of modern skincare.
You meticulously apply expensive, high-molecular-weight hyaluronic acid serums and rich, heavy creams to your face.
For a brief, fleeting hour, your skin feels plump, saturated, and artificially hydrated. But by mid-afternoon, that temporary comfort vanishes entirely, replaced by a tightening, suffocating grip. Your skin turns dry, brittle, and visibly flaky under the harsh, unforgiving light of the sun.
This is the undeniable physical sensation of an evaporating moat. The Systemic Commander has been successfully deployed deep within the dermal matrix to absorb the radiation.
Astaxanthin is actively absorbing the thermodynamic shockwaves and silencing the inflammatory alarms below the surface. But the absolute outermost perimeter of your biological fortress has been catastrophically breached.
The microscopic roof is missing, and the internal cellular water is simply boiling off into the dry atmosphere.

THE ILLUSION OF HYDRATION
The failure of topical humectants in a structurally compromised system.
We must forensically deconstruct the grand biological fallacy sold by the mass-market beauty industry. Chronically dry, severely photo-aging skin is never fundamentally a water problem.
It is a profound, devastating structural leak problem occurring at the exact microscopic level of the stratum corneum. This continuous, invisible hemorrhage is scientifically classified as severe Transepidermal Water Loss, or TEWL.
TEWL is the precise, clinically measurable rate at which your essential internal moisture escapes through a shattered epidermal barrier. When relentless ultraviolet radiation bombards the skin, it physically oxidizes and chemically dissolves the delicate lipid glue holding your surface cells together.
These flat, hardened keratinocytes are biologically programmed to overlap perfectly, functioning like impenetrable, heavy roofing shingles. Instead, the kinetic force of the UV strikes and the resulting oxidative stress cause them to curl, lift, and violently fracture.
This structural microscopic warping exposes the delicate, highly vulnerable, water-rich living tissues resting directly underneath. Applying a topical, water-binding humectant serum to this devastated, irradiated landscape is a mathematical and physical failure. It is the exact physical equivalent of desperately pouring expensive water into a heavy wooden bucket with a completely shattered, rotting bottom.
The water does not stay; it physically cannot stay, because the precise mechanical container required to trap it no longer exists.
The applied topical humectant merely sits exposed on the fractured surface, acting as a temporary chemical sponge.
In fact, it often aggressively draws essential, surviving water up from your deeper, living dermal tissues to the broken surface. Once the dry atmospheric air hits that highly exposed, vulnerable moisture, it violently and rapidly evaporates the humectant.
It takes your precious, deep cellular water right along with it into the void. This tragic osmotic backfire leaves your underlying fibroblasts significantly more dehydrated, brittle, and structurally exhausted than before you applied the serum.
We absolutely cannot permanently solve a catastrophic, deep-tissue structural failure with a temporary layer of superficial dampness.

THE CALL FOR THE BRICKLAYER
Initiating the lipidomics re-engineering of the stratum corneum.
To permanently halt this microscopic hemorrhage of essential cellular water, we must entirely abandon the mass-market illusion of surface hydration.
We must forcefully transition from passive moisturizing into the aggressive, clinically precise science of lipidomics re-engineering.
The biological fortress does not need another useless, temporary flood of topical water applied to its dead exterior. It urgently and mathematically demands the immediate, systemic deployment of a highly specific, structural biological mortar.
We must literally cement the lifting, fractured keratinocytes of the stratum corneum back together from the inside out, utilizing your body’s own metabolic pathways.
This is the exact, critical moment we call upon the first and most vital structural component of the Keyora Bioactive Carrier system.
We must initiate the precise systemic deployment of Linoleic Acid, the essential Omega-6 fatty acid meticulously extracted from high-purity Flaxseed.
Linoleic Acid is not just another generic, inert dietary fat floating aimlessly in the central bloodstream. It is the ultimate, irreplaceable biochemical bricklayer of the entire human epidermal system.
Nature explicitly and specifically designed this polyunsaturated molecule to serve as the absolute foundational mortar of your waterproof skin barrier.
Without a continuous, unoxidized supply of Linoleic Acid successfully reaching the upper epidermal layers, complex ceramide synthesis completely and permanently halts. The structural wall literally cannot be built, the microscopic cracks cannot be filled, and the devastating Transepidermal Water Loss becomes an irreversible, chronic condition.
It dictates whether your skin remains a plump, resilient shield or degenerates into a dry, cracking, and highly permeable membrane. Under the absolute, unbroken thermodynamic cover fire provided by Astaxanthin in the deeper layers, this highly vulnerable lipid mortar can finally be safely transported to the surface.
It is time to permanently seal the evaporating moat, reconstruct the fractured lipid matrix, and trap the life-sustaining moisture permanently inside the cellular fortress.

3.1 THE ARCHITECTURE OF ACYL-CERAMIDES
Deconstructing the precise lipid geometry required to waterproof the human stratum corneum.
The skin’s barrier is absolutely not random fat smeared across the surface of your body; it is nature’s most impenetrable, mathematically perfect lipid geometry.
To the untrained eye, or the mass-market cosmetic formulator, the surface of the skin is just a dry canvas waiting to be painted with synthetic grease.
We must entirely reject this primitive biological viewpoint. The stratum corneum is a highly advanced, meticulously engineered biophysical fortress.
It operates under strict laws of structural chemistry, where the absence of a single, specific lipid molecule causes the entire architectural grid to catastrophically fail.
PROPOSITION A:
Can the skin retain moisture without a precise structural lipid grid?
The physical impossibility of a mortar-less brick wall.
We must examine the exact, microscopic anatomy of the stratum corneum to understand the sheer scale of the moisture crisis.
Dermatologists universally describe the outermost layer of your skin using the “brick and mortar” structural model.

The Evidence:
The forensic markers of Transepidermal Water Loss (TEWL).
The “bricks” are the corneocytes: dead, heavily keratinized, flattened cells that have entirely sacrificed their internal nuclei to become solid armor plates. These microscopic armor plates are physically incredibly tough, but they are entirely incapable of stopping water from passing between them.
If you stacked these corneocytes without a highly specific sealing agent, your internal body water would completely evaporate into the atmosphere in a matter of hours.
The absolute biological secret to human terrestrial survival lies entirely within the “mortar.”
This mortar is a highly complex, intercellular lipid matrix that physically surrounds, coats, and structurally binds every single corneocyte together. It is a precisely organized three-dimensional grid composed of ceramides, free fatty acids, and cholesterol, arranged in tightly stacked, repeating sheets called lipid lamellae.
When ultraviolet radiation and severe oxidative stress initiate the localized disaster of [The Neuro-Endocrine Storm], this delicate lipid mortar is the very first casualty.
The highly reactive free radicals physically attack the lipid sheets, violently cleaving the carbon bonds and destroying the geometric grid.
The heavy corneocyte bricks immediately begin to physically separate, warp, and lift away from the underlying tissue.
This structural collapse directly triggers severe Transepidermal Water Loss, or TEWL.
You do not just lose a few drops of surface sweat; you actively hemorrhage the deep, vital cellular water required to keep your dermal fibroblasts alive and functioning.
The biological consequences of this structural failure extend far beyond the immediate, painful sensation of dry, cracking skin.
We must introduce the terrifying optical physics of a dehydrated, mortar-less stratum corneum.
When the lipid matrix degrades and the corneocytes warp, the physical surface of your skin transforms from a smooth, reflective biological shield into a jagged, microscopic landscape of rough edges.
A smooth, perfectly hydrated stratum corneum naturally reflects and bounces a significant percentage of incoming ultraviolet light safely away from the body.
But a dry, rough, and deeply fractured surface causes that exact same incoming UV light to scatter maliciously in a thousand unpredictable directions.
The jagged, warped cellular edges physically trap the high-velocity photons inside the upper epidermal layers.
The radiation bounces internally, violently striking the surviving tissue over and over again, exponentially amplifying the localized radiation damage.
Your broken, dehydrated skin barrier literally acts as a microscopic magnifying glass, intensely focusing the destructive power of the sun directly into your vulnerable tissue.
The Evaporating Moat is not just a cosmetic inconvenience; it is a profound biophysical vulnerability that accelerates the total destruction of the entire biological matrix.

The Mechanism:
Linoleic Acid as the terminal rivet of the ceramide matrix.
To permanently stop this catastrophic internal evaporation and optical scattering, the body must physically rebuild the shattered lipid lamellae.
It must manufacture new, flawless ceramide molecules deep within the stratum granulosum and secrete them upward to seal the dead corneocyte bricks.
However, not all ceramides are structurally capable of waterproofing the human body.
The standard ceramides floating in your tissue are too short to physically bridge the wide microscopic gaps between the stacked cellular plates.
To achieve absolute, uncompromising impermeability, the epidermal architecture demands the synthesis of a highly specialized, massive lipid structure known as an O-acylceramide.
Let us dive deep into the precise, uncompromising chemistry of this exact molecule.
An O-acylceramide is a biological behemoth, featuring a profoundly long omega-hydroxy fatty acid chain attached to a sphingoid base. This massive carbon backbone is physically long enough to punch straight through multiple stacked layers of the lipid matrix.
But length alone is biologically useless if the molecule cannot be permanently anchored into place. The long carbon chain requires a highly specific, terminal locking mechanism to structurally rivet the entire lipid grid together, preventing the sheets from sliding apart under physical friction.
Nature explicitly selected Linoleic Acid, an 18-carbon Omega-6 polyunsaturated fatty acid, for this exact, irreplaceable bio-architectural job.
Linoleic Acid acts as the absolute terminal rivet of the entire ceramide matrix.
Through a highly complex, enzyme-driven metabolic process, the body chemically esterifies a single, unoxidized Linoleic Acid molecule directly onto the omega-hydroxyl tail of the massive ceramide structure.
This specific chemical attachment creates a unique, rigid, and deeply anchored molecular joint.
The Linoleic Acid rivet physically stitches the multiple, overlapping sheets of the lipid lamellae tightly together, locking the corneocyte bricks into a permanent, waterproof configuration.
This specific molecular geometry entirely blocks the microscopic escape routes that water molecules previously used to evaporate into the atmosphere.
We must forensically examine what happens when the body is chronically deficient in Linoleic Acid, or when the existing LA is completely destroyed by UV-induced lipid peroxidation.
The cellular machinery in the epidermis operates on a blind, desperate biological mandate to build the wall at all costs.
If it cannot find fresh, unoxidized Linoleic Acid to finish the O-acylceramide construction, it panics and violently substitutes whatever fat is locally available. It typically grabs Oleic Acid, a widely available Omega-9 fatty acid, and forcibly jams it into the terminal slot where the LA rivet belongs.
This is a catastrophic bio-architectural mutation.
Oleic Acid possesses a fundamentally different three-dimensional shape, featuring a distinct, sharp chemical kink in its carbon backbone that entirely prevents tight molecular packing.
It physically cannot form the flush, tight, and highly organized rivet required to stitch the lipid sheets together.
-
The resulting mutated ceramide grid is loose, structurally disorganized, and highly porous.
-
The biological wall is technically built, but it is structurally defective and completely incapable of holding internal hydrostatic pressure.
-
The internal cellular water simply bypasses the mutated ceramide joints and rapidly escapes into the void, guaranteeing the continuation of severe, chronic TEWL.
-
The barrier completely collapses, leaving the deep dermal layers totally exposed to continuous radiation, environmental toxins, and pathogenic invasion.

The Resolution:
The biological mandate for endogenous LA synthesis.
The biological reality is absolute and mathematically undeniable.
Linoleic Acid is not just a standard, easily replaceable dietary fat; it is the ultimate, non-negotiable biological mortar required for human terrestrial survival.
You cannot achieve true, permanent hydration by randomly slathering mass-market, synthetic ceramides onto the dead surface of your face. The human epidermis actively rejects and repels these large, foreign topical lipids, refusing to structurally integrate them into the intricate internal lamellar matrix.
The biological wall can only be successfully rebuilt from the inside out, utilizing your body’s own precise, localized enzymatic machinery. Therefore, the human body must be continuously, systemically supplied with a pure, heavy dose of completely unoxidized Linoleic Acid.
This essential Omega-6 precursor must safely travel through the central bloodstream, successfully navigate the deep dermal capillary networks, and arrive fully intact at the stratum granulosum.
-
Only then can the specialized cellular architects properly execute the flawless synthesis of O-acylceramides.
-
Only then can the terminal rivets be locked into place, completely sealing the microscopic fractures and ending the catastrophic hemorrhage of your internal moisture.
We must secure the physiological delivery of this specific biological mortar, but as we will soon discover, transporting this highly volatile molecule through a heavily irradiated combat zone is an incredibly dangerous mission.

3.2 THE FATAL VULNERABILITY OF OMEGA-6
Why the exact double bonds that provide structural flexibility become the primary targets for singlet oxygen.
Transporting raw, unstable high-explosives across an actively burning, heavily irradiated battlefield is a guaranteed biological suicide mission.
In the intricate architecture of the human epidermis, Linoleic Acid is that exact high-explosive.
We have firmly established that this essential Omega-6 fatty acid is the absolute, non-negotiable biological mortar required to waterproof the stratum corneum. However, we must now forensically confront the terrifying chemical reality of its specific, highly vulnerable molecular structure.
The exact biochemical features that make Linoleic Acid the perfect, flexible structural mortar also make it the ultimate target for localized radiation damage.

PHASE 1: THE STRUCTURAL NECESSITY
The requirement of polyunsaturated flexibility in a dynamic barrier.
To understand this fatal vulnerability, we must forensically examine the exact atomic architecture of Linoleic Acid. It is not a rigid, straight-chain saturated fat like the heavy, solid lipids that make up dense, immobile animal blubber.
Linoleic Acid is a highly specialized polyunsaturated fatty acid, characterized by a long eighteen-carbon chain featuring two distinct molecular double bonds.
These specific double bonds are located precisely at the 9th and 12th carbon positions, representing a masterpiece of bio-architectural engineering. They create strict, physical kinks in the long carbon backbone, preventing the lipid sheets from packing together too tightly into a solid, frozen block.
This precise degree of atomic spacing is what grants your skin barrier its essential, dynamic mechanical flexibility.
When you smile, frown, or stretch your facial muscles, the skin physically expands and contracts across a massive surface area. If your ceramide matrix were built entirely out of stiff, saturated fats, your stratum corneum would literally possess the physical properties of cold wax.
The very first time you smiled, the stiff, unyielding lipid barrier would violently shatter and crack like brittle glass under the mechanical stress.
The double bonds of Linoleic Acid function as microscopic biological shock absorbers, allowing the lipid lamellae to stretch and instantly snap back into place.
They give the O-acylceramide structures the precise liquid-crystal fluidity required to survive constant, daily biomechanical friction and rapidly shifting environmental temperatures.
Without the polyunsaturated nature of Linoleic Acid, human skin could not physically function as a dynamic, moving organ. But in the harsh, unforgiving biophysics of a high-radiation environment, this beautiful structural flexibility comes at a catastrophic metabolic price.
The electrons forming those two essential double bonds are held in a highly exposed, chemically tense configuration within the cell membrane.
They represent a massive, glaring vulnerability resting directly in the exact center of your primary biological defensive wall.

PHASE 2: THE UV STRIKE
When the 415nm photon breaches the outer defense.
We must now return to the violent, microscopic theater of localized oxidative stress actively raging within the dermal and epidermal layers.
Imagine the fragile Linoleic Acid molecules successfully navigating upward, attempting to reach the stratum granulosum to build the ceramide wall. Suddenly, the biological airspace is forcefully breached by a relentless, high-velocity barrage of incoming solar radiation.
A rogue ultraviolet photon pierces straight through the outer keratinocyte armor, carrying a devastating payload of pure kinetic energy. It does not hit the Linoleic Acid directly, but instead strikes a passive, ground-state oxygen molecule resting quietly nearby in the extracellular matrix.
The sheer kinetic impact of this photon collision violently ejects an electron from the normal oxygen molecule, radically altering its fundamental quantum state.
As we established in the deeper layers of the dermal matrix, this high-speed collision instantly gives birth to the Singlet Oxygen bomb. This hyper-excited, highly volatile free radical is no longer a benign, life-giving gas; it is a rabid, desperately unbalanced chemical scavenger.
It is violently spinning through the tissue, carrying a lethal payload of pure excitation energy, desperately searching for any available electron to stabilize its collapsing orbit. Singlet Oxygen absolutely does not attack the strong, tightly bound single bonds of standard structural proteins or dense saturated fats.
It specifically and aggressively hunts for the exact chemical structures that are the weakest and most exposed in the entire cellular environment.
To a rabid Singlet Oxygen molecule, the exposed double bonds of polyunsaturated Linoleic Acid look like a massive, glowing molecular target. The electrons suspended in the bis-allylic positions, which are the exact carbon bridges located perfectly between the two double bonds of the LA molecule, are highly vulnerable.
They are held extremely loosely by the carbon backbone, making them incredibly easy for a violent free radical to physically grab and tear away.
The high-explosive mortar is now fully exposed on the battlefield, and the thermodynamic spark has just been successfully ignited.

PHASE 3: THE PEROXIDATION CHAIN REACTION
The catastrophic burning of unprotected Linoleic Acid.
When the volatile singlet oxygen bomb successfully locks onto the exposed double bonds of the biological mortar, it initiates a terrifying, irreversible chemical cascade.
The Hydrogen Theft:
The hyper-excited singlet oxygen violently slams directly into the carbon backbone of the unprotected Linoleic Acid molecule. It acts as a microscopic, high-energy crowbar, aggressively targeting the weak bis-allylic carbon bridge located exactly between the two double bonds.
With sheer kinetic and electromagnetic force, the radical violently rips a single hydrogen atom directly away from the lipid’s structural framework. This sudden, brutal atomic theft instantly deforms the previously stable Linoleic Acid molecule, completely destroying its perfect bio-architectural geometry.
The essential biological mortar has now been catastrophically mutated into a carbon-centered lipid radical, becoming a highly unstable and frantic entity itself.
The Oxygen Addition:
The microscopic disaster exponentially accelerates the exact microsecond this new lipid radical is born in the cellular matrix.
Because the mutated Linoleic Acid is now desperately missing an electron, it instantly reacts with the abundant, normal triplet oxygen dissolved in the surrounding tissue.
This rapid, violent oxygen addition forcibly jams two new oxygen atoms directly into the fractured carbon backbone of the lipid structure.
This chemical fusion creates a Lipid Peroxyl Radical, an incredibly toxic, highly reactive, and profoundly damaging biological monstrosity.
The beautiful, flexible Omega-6 mortar originally designed to waterproof your skin has just been successfully weaponized against your own cellular architecture.
The Unstoppable Wildfire:
This newly formed Lipid Peroxyl Radical is now a localized biological terror, entirely capable of continuing the destruction completely on its own.
It violently attacks the very next healthy, unprotected Linoleic Acid molecule sitting beside it in the ceramide matrix.
It ruthlessly steals a hydrogen atom from its neighbor to temporarily stabilize itself, which instantly turns that neighbor into a brand new, highly destructive radical.
This creates an unstoppable, self-propagating chemical wildfire known to forensic pathologists and dermatologists as rapid Lipid Peroxidation. The protective ceramide grid literally burns itself to the ground in a cascading, exponential chain reaction of microscopic molecular theft.
As the delicate lipid lamellae violently burn, they leave behind a massive, highly toxic accumulation of biological ash. This localized ash is primarily composed of Malondialdehyde (MDA) and 4-Hydroxynonenal (4-HNE), which are incredibly destructive chemical byproducts.
These toxic byproducts are entirely responsible for driving the deep, agonizing cellular inflammation, the extreme redness, and the total collapse of the moisture barrier.
The toxic MDA does not simply vanish; it physically seeps down into the dermis and aggressively cross-links with your surviving collagen fibers. It chemically bonds with the pristine Type I collagen, causing it to stiffen, snap, and create the deep, leathery cross-hatching visible on severely sun-damaged skin.
The unprotected Linoleic Acid has completely failed to build the waterproof wall; instead, it has actively accelerated the structural decay of the entire organ.
The essential mortar was deployed completely naked into the combat zone, and the localized radiation entirely vaporized the supply lines.
We now mathematically understand why simply swallowing unprotected Omega-6 capsules before walking into the sun is a profound, scientifically verifiable biological error.
You are merely providing massive amounts of raw, explosive fuel for the Singlet Oxygen bombs to continuously ignite and burn within your stratum corneum.
To successfully build the waterproof epidermal wall, the Linoleic Acid must be transported, integrated, and synthesized under conditions of absolute thermodynamic peace.

3.3 THE ULTIMATE COVERING FIRE:
ASTAXANTHIN AND LA SYNERGY
How the Transmembrane Commander physically shields the biological mortar during the construction of the epidermal wall.
A master architect cannot successfully build a complex, load-bearing wall while pinned down under heavy, continuous sniper fire.
If the essential building materials are systematically blown to pieces the exact moment they arrive at the construction site, the fortress will inevitably fall.
The deeply irradiated human epidermis is precisely this hostile, actively burning combat zone. To physically rebuild the shattered stratum corneum, the delicate Linoleic Acid mortar absolutely requires a supreme tactical escort.
It strictly demands the absolute, unbroken covering fire of the Transmembrane Commander to safely survive the biological battlefield.

THE BIOACTIVE CARRIER DELIVERY
Smuggling the LA source code past the digestive destruction.
The journey of deep epidermal reconstruction does not begin at the superficial surface of the skin; it begins deep within the human gastrointestinal tract. To successfully seal the Evaporating Moat, we must first introduce a massive, systemic payload of pure, unoxidized Linoleic Acid into the central biology.
Keyora completely rejects the use of cheap, highly processed, and pre-oxidized commercial seed oils that already contain toxic, pre-formed lipid peroxides. Instead, we strictly utilize cold-pressed, high-purity Flaxseed Oil to act as the ultimate, heavily armored Bioactive Carrier.
This highly specialized lipid vehicle is biologically engineered to smuggle the fragile LA source code past the very first major physiological threat: digestive destruction.
When you ingest this powerful biological mortar, it immediately faces the highly acidic, violently churning, and enzymatically aggressive environment of the human stomach.
Standard, unprotected dietary fats often undergo rapid, premature degradation here, losing their precise atomic geometry before they even reach the central bloodstream.
But the Bioactive Carrier successfully shields the Linoleic Acid, allowing it to pass flawlessly into the small intestine for perfect, uncompromised metabolic absorption.
Once absorbed through the intestinal wall, these pristine Omega-6 molecules are carefully packaged into specialized lipid transport vehicles known as chylomicrons. They are then processed by the liver into Very Low-Density Lipoproteins, seamlessly integrating into the central circulatory system to begin their perilous journey.
As the lipoproteins travel through the vast, branching network of your central arteries and peripheral veins, they face a secondary, highly toxic threat. The human bloodstream is frequently flooded with systemic oxidative stress, circulating metabolic waste products, and rogue inflammatory cytokines.
If the fragile double bonds of the Linoleic Acid are exposed to this internal oxidative friction, they will instantly burn and mutate into malondialdehyde in transit.
This is exactly where the systemic, whole-body presence of Astaxanthin first begins to exert its profound, protective bio-architectural dominance. Because Astaxanthin is also highly lipophilic by nature, it physically travels inside the exact same lipid transport vehicles as the fragile Omega-6 mortar.
The Commander physically rides shotgun with the essential building materials, actively sweeping the internal circulatory environment for any circulating oxidative threats.
It aggressively intercepts rogue free radicals in the bloodstream, ensuring that the Linoleic Acid remains perfectly pristine, unoxidized, and structurally flawless.
-
The Bioactive Carrier successfully navigates the deep central circulation and finally arrives at the microscopic capillary networks feeding the lower dermal layers.
-
The pristine mortar is forcefully pushed out of the microvasculature, moving into the extracellular matrix to begin its final, vertical ascent.
-
The critical smuggling operation is a total physiological success, but the most dangerous, highly explosive phase of the reconstruction mission is just beginning.

THE THERMODYNAMIC INTERCEPT
Astaxanthin absorbing the photon strike before it reaches the lipid mortar.
As the pristine Linoleic Acid molecules leave the relative safety of the deep dermal capillaries, they physically enter the absolute kill zone of the upper skin. This is the exact microscopic territory where the relentless, 11-hour ultraviolet bombardment from the afternoon sun is actively detonating and destroying tissue.
The fragile Omega-6 mortar must physically move upward through this highly irradiated extracellular matrix to reach the specific enzymes that build the waterproof ceramides.
Suddenly, a catastrophic, high-velocity 415nm UVA photon violently penetrates the epidermal airspace and directly strikes the surrounding aqueous tissue.
The massive kinetic impact of the photon instantly shatters a local oxygen molecule, forcefully ejecting an electron and birthing the lethal Singlet Oxygen bomb.
This hyper-excited, rapidly spinning free radical is no longer a benign gas; its volatile quantum state is desperately seeking an exposed electron to steal.
It immediately scans the environment and locks onto the glaring vulnerability of the newly arrived Linoleic Acid: the weak bis-allylic carbon bridge. The microscopic bomb hurtles toward the fragile biological mortar at terrifying, catastrophic speed, threatening to ignite an unstoppable lipid peroxidation wildfire.
If it makes physical contact, the mortar will instantly mutate into a toxic peroxyl radical, and the entire bio-architectural mission will end in catastrophic failure. But the highly vulnerable Linoleic Acid is not operating entirely alone in this deadly, irradiated biological void; it has entered a Keyora-fortified matrix.
Exactly a fraction of a nanosecond before the Singlet Oxygen can physically slam into the fragile Omega-6 double bonds, a massive biological wall intervenes. It is the uncompromising, heavily anchored, 30-Angstrom carbon architecture of Astaxanthin, the supreme Transmembrane Commander, firmly embedded in the local cells.
Because Astaxanthin is perfectly, vertically riveted throughout the entire cellular lipid bilayer, it completely dominates the local thermodynamic airspace of the epidermis. It physically throws its massive, conjugated polyene chain directly into the exact trajectory of the incoming, high-velocity Singlet Oxygen bomb.
The Commander executes a flawless, mathematically perfect thermodynamic intercept, completely shielding the highly vulnerable Linoleic Acid resting just behind it in the matrix.
The hyper-excited Singlet Oxygen violently crashes into the Astaxanthin shield, but the Commander absolutely refuses to yield, break, or chemically react.
Instead, its dense, impenetrable cloud of delocalized electrons instantly catches the raw, volatile excitation energy, physically stripping the lethal payload from the oxygen.
The localized thermodynamic shockwave violently surges directly into the massive carbon backbone of the defender, but the energy absolutely never reaches the Linoleic Acid.
Astaxanthin immediately initiates its extreme vibrational cooling protocol, rapidly shaking and oscillating to mechanically dissipate the catastrophic kinetic energy. The lethal, structural-shattering force of the UV strike is brilliantly downgraded and bled into the surrounding fluid as entirely harmless, microscopic heat.
The attacking singlet oxygen is instantly dropped back to a docile, passive ground state, entirely defused and biologically pacified without a single electron being lost.
The Astaxanthin molecule safely stops vibrating, perfectly retaining its complete, unbroken structural integrity, instantly ready to absorb the very next incoming photon.
Most importantly, we must forensically examine the exact biological state of the Linoleic Acid molecule resting safely in the thermodynamic shadow of the Commander.
Its precise 18-carbon chain remains absolutely, flawlessly intact, and its two essential double bonds are completely untouched and highly flexible.
The explosive thermodynamic threat was perfectly intercepted and completely nullified the exact millisecond it was generated, providing the ultimate covering fire.

THE ARCHITECTURAL VICTORY
The seamless integration of protected mortar into the stratum corneum.
Because the Transmembrane Commander successfully absorbed the catastrophic photon strike, the fragile biological mortar survives the perilous journey through the kill zone.
The pristine, entirely unoxidized Linoleic Acid successfully bypasses the localized oxidative fire and safely arrives at the microscopic construction site.
It enters the deep stratum granulosum, the exact, highly specialized cellular layer where the advanced lipid-synthesizing enzymes operate around the clock.
These hard-working enzymes finally have access to the exact, mathematically perfect biological building blocks they desperately require to finish the architectural grid.
They immediately seize the highly flexible Linoleic Acid molecules and meticulously execute the complex, incredibly precise chemical esterification process.
The cellular enzymes physically attach the pure Linoleic Acid directly onto the extremely long omega-hydroxyl tail of the massive ceramide precursor molecules.
With a precise, molecular snap, the terminal rivet is permanently locked into place, officially birthing the impenetrable, massive O-acylceramide structure. These massive, structurally flawless lipid grids are then forcefully extruded upward into the fractured, leaking gaps between the dead, stacked corneocyte bricks.
Because the Linoleic Acid rivet possesses the exact correct three-dimensional atomic geometry, the lipid sheets pack together perfectly tightly. They absolutely do not form the loose, highly porous, and mutated barriers that result when the panicked body is forced to scavenge incorrect fats like Oleic Acid.
-
The fully protected mortar flawlessly stitches the overlapping lipid lamellae together, actively cementing the warped, lifting keratinocytes flat against the surface.
-
The jagged, broken, and highly irregular microscopic landscape of your skin is systematically smoothed down, completely eliminating the devastating optical UV scattering effect.
-
The massive, gaping structural breaches that previously allowed your internal cellular water to rapidly evaporate into the atmosphere are firmly and permanently sealed.
-
The catastrophic, chronic Transepidermal Water Loss is entirely halted, trapping the vital, life-sustaining moisture deep inside the dermal fibroblasts where it belongs.
-
The Evaporating Moat is officially closed, hydrostatic pressure is completely restored, and the upper perimeter of your biological fortress is entirely locked down.
This is the undeniable, mathematically verified triumph of true, systemic Lipidomics Re-engineering operating perfectly within the human body. It completely exposes the primitive, scientifically bankrupt fallacy of relying on topical, surface-level hydration to fix a deep, structural internal leak.
You absolutely cannot permanently waterproof a complex biological organ without simultaneously deploying the exact internal mortar alongside the absolute thermodynamic shield.
The flawless, synergistic partnership between the raw building power of Linoleic Acid and the unbroken defensive supremacy of Astaxanthin is strictly non-negotiable.
The outer wall is finally, successfully rebuilt, but to achieve total dermal sovereignty, we must soon address the terrifying mechanical friction actively building deep below.

3.4 CLINICAL EVIDENCE & SCIENTIFIC CONSENSUS:
THE BARRIER VERDICT
Translating the theoretical brilliance of the Astaxanthin-LA synergy into undeniable, peer-reviewed human clinical outcomes.
The Keyora bio-architectural protocol is absolutely not a fragile, untested laboratory hypothesis confined to the sterile vacuum of a petri dish.
Theoretical biochemistry, no matter how elegant or mathematically sound on paper, is entirely useless if it fails to perform inside a living, breathing human organism.
To permanently validate the systemic deployment of the Transmembrane Commander and the Linoleic Acid mortar, we must leave the realm of microscopic theory.
We must subject this exact biological synergy to the most brutal, unforgiving environment in modern medical science: the courtroom of evidence-based dermatology.
We must forensically examine the hard, peer-reviewed, double-blind clinical data to definitively prove that this endogenous reconstruction successfully stops the physical decay of the human face.
PROPOSITION B:
Does the endogenous lipid-antioxidant matrix clinically reverse barrier dysfunction and photo-aging?
The courtroom of evidence-based dermatology.
Before we can evaluate the protective power of the shield, we must first clinically validate the absolute necessity of the biological mortar itself.

The Evidence (The LA Consensus):
Validating Linoleic Acid’s role in reversing epidermal degradation.
The foundational dermatological consensus regarding Linoleic Acid was definitively established by the rigorous, landmark clinical research of experts like Ziboh and Chapkin in 1987.
Their extensive biochemical analyses of the human epidermis provided the exact, undeniable forensic proof that LA is the mandatory structural component of the skin barrier.
When a human subject is subjected to a strict, clinically induced deficiency of essential Omega-6 fatty acids, the biological results are rapid and catastrophic.
Within a matter of weeks, the subject’s skin barrier forcefully collapses, developing a severe, highly painful, and scaling epidermal dermatosis.
The clinical bio-instrumentation used on these deficient subjects reveals a massive, terrifying spike in Transepidermal Water Loss.
Without the Linoleic Acid acting as the terminal rivet, the O-acylceramide structures physically disintegrate, and the internal cellular moisture completely evaporates into the atmosphere. The skin physically hardens, cracks, and becomes highly susceptible to extreme erythema, bacterial infiltration, and severe localized inflammatory cascades.
However, the true medical breakthrough occurs when researchers reintroduce pure, unoxidized Linoleic Acid back into the subject’s systemic circulatory system.
The clinical reversal of the barrier dysfunction is not merely cosmetic; it is a profound, rapid, and mathematically quantifiable structural bio-reconstruction.
As the systemic Linoleic Acid successfully navigates the capillary beds and reaches the stratum granulosum, the localized cellular machinery immediately goes to work.
The epidermal enzymes aggressively synthesize new, flawless O-acylceramides, utilizing the fresh LA molecules to permanently rivet the lipid lamellae back together. Clinical measurements physically demonstrate a rapid, steep decline in TEWL, proving that the microscopic leaks in the stratum corneum have been successfully and tightly sealed.
Furthermore, the clinical reintroduction of LA directly modulates the localized epidermal immune response, significantly reducing the visual redness and painful swelling associated with erythema.
This establishes the undeniable medical baseline: Linoleic Acid is the ultimate, non-negotiable biological mortar required to physically reverse epidermal structural degradation.
But as we have previously established, deploying this highly vulnerable, explosive mortar into a heavily UV-irradiated environment without thermal protection is a guaranteed failure. We must now introduce the massive, peer-reviewed clinical data proving that Astaxanthin successfully provides the absolute thermodynamic covering fire required for this mortar to survive.
The Evidence (The Astaxanthin Consensus): Subtitle: The human clinical trials on Astaxanthin’s internal beauty effects.
The theoretical biophysics of the Astaxanthin singlet oxygen trap have been flawlessly translated into verifiable, systemic anti-aging outcomes across multiple rigorous human clinical trials.

The Tominaga Trial (2012):
Tominaga and colleagues orchestrated a highly rigorous, double-blind, placebo-controlled human clinical trial to test the systemic efficacy of oral Astaxanthin.
They administered a precise, daily systemic dose of 6 milligrams of Astaxanthin to healthy human subjects over a continuous period of eight weeks. The researchers absolutely refused to rely on subjective, unreliable patient questionnaires, opting instead to utilize highly calibrated, advanced bio-instrumentation to measure the exact physiological changes.
They deployed a specialized dermatological tool known as a Tewameter to physically measure the exact volume of water evaporating through the subjects’ skin barriers. The clinical data revealed a statistically profound, mathematically undeniable reduction in Transepidermal Water Loss across the entire Astaxanthin-supplemented group.
This precise TEWL reduction is the ultimate, undeniable forensic proof that the deep cellular barrier was successfully and permanently sealed from the inside out.
Because the Astaxanthin successfully embedded into the cellular membranes and absorbed the catastrophic photon strikes, the endogenous Linoleic Acid mortar survived the radiation. The LA was not violently oxidized into toxic malondialdehyde; instead, it successfully synthesized the O-acylceramides required to pack the corneocytes tightly together.
Furthermore, Tominaga’s team utilized highly advanced 3D micro-topography imaging to map the deep, microscopic structural valleys of the subjects’ facial wrinkles. The topographic data proved a highly significant, physical reduction in both the maximum depth and the maximum width of the existing structural wrinkles.
This visual, structural smoothing of the deep epidermal valleys is the direct, clinical confirmation of the Collagen Guardian mechanism we detailed in Chapter 2.
The Astaxanthin completely confiscated the MMP-1 and MMP-3 molecular scissors, preventing the localized fibroblasts from shredding their own underlying Type I collagen scaffolding.
The underlying structural floor was completely stabilized, while the upper epidermal roof was permanently waterproofed by the protected Linoleic Acid rivets.

The Yamashita Trial (2006):
The undeniable structural efficacy of this systemic biological protocol was further validated by the extensive clinical data published by Yamashita in 2006. Yamashita tracked the internal deployment of oral Astaxanthin over a slightly shorter, highly intensive clinical window of four to six weeks.
This trial specifically focused on the physical, structural morphology of the dead corneocyte cells resting on the absolute outermost surface of the stratum corneum.
Using advanced tape-stripping techniques and high-resolution electron microscopy, the researchers visually examined the microscopic geometry of the skin’s armor plates. In the severely photo-aged, placebo-controlled subjects, the corneocytes were highly irregular, shriveled, and violently peeling upward away from the lipid matrix.
This lifting and warping is the exact microscopic root cause of the dry, flaky texture and the malicious, damaging optical scattering of incoming UV light.
However, in the subjects receiving the systemic Astaxanthin covering fire, the clinical morphology of the superficial corneocytes completely and rapidly transformed. The electron microscopy data proved that the corneocyte area significantly expanded, and the cells laid perfectly flat, smooth, and tightly bound to the surface.
This flattening and expansion is the exact, undeniable visual evidence that the Linoleic Acid rivets were successfully locked into place without being burned by lipid peroxyl radicals. Because the thermodynamic shield neutralized the volatile oxidative shrapnel, the lipid lamellae remained perfectly fluid, allowing the corneocytes to properly desquamate and align.
Yamashita’s clinical data also documented a profound, statistically significant improvement in overall epidermal elasticity and a visible reduction in fine, dry lines. When the stratum corneum is perfectly sealed with unoxidized ceramides, the trapped internal hydrostatic pressure physically pushes outward from the deep dermis.
This internal water pressure acts exactly like air filling a deflated biological balloon, automatically smoothing out the fine, superficial dehydration lines on the surface. The clinical subjects achieved a highly luminous, perfectly hydrated, and structurally resilient complexion that no mass-market topical cream could ever mechanically replicate.

The Resolution:
The undeniable triumph of the endogenous shield.
The verdict from the courtroom of evidence-based dermatology is absolutely final, mathematically sound, and biologically undeniable.
The hard, peer-reviewed clinical data perfectly and flawlessly maps to the highly complex, microscopic biophysical mechanisms we have forensically deconstructed.
The clinical trials absolutely prove that you cannot permanently reverse severe photo-aging and chronic barrier dysfunction by smearing synthetic water on dead tissue.
You must execute a highly coordinated, systemic lipidomics re-engineering protocol from deep within the central circulatory system. The Transmembrane Commander must be deployed to construct an unbreakable, thermodynamic heat-sink across the cellular lipid bilayers.
Astaxanthin flawlessly executes the physics of quenching, intercepting the singlet oxygen bombs and bleeding the lethal radiation away as harmless microscopic heat. This absolute, unbroken thermodynamic covering fire provides the highly specific, non-negotiable safe passage required for the Bioactive Carrier system.
The pure, unoxidized Linoleic Acid safely navigates the irradiated combat zone, entirely avoiding the catastrophic, self-propagating chain reaction of lipid peroxidation. It arrives exactly at the stratum granulosum intact, where the localized enzymes successfully utilize it as the terminal rivet to build massive, waterproof O-acylceramides.
These flawless ceramides permanently cement the stratum corneum, completely halting Transepidermal Water Loss, and physically restoring the skin’s optical, reflective brilliance.
This is the ultimate, undeniable triumph of the 1+1+1+1+1+1+1 > 7 Keyora bio-architectural synergy.
Astaxanthin is the absolute, uncompromising thermodynamic shield, and Linoleic Acid is the irreplaceable, fundamental biological mortar. When deployed together in a heavily fortified systemic matrix, they achieve a total, complete reversal of deep tissue degradation that superficial lotions can never replicate.
The Evaporating Moat has been permanently sealed, the catastrophic internal hemorrhage of vital cellular water has been completely halted, and the outer perimeter is secure.
The clinical data stands as an indestructible monument to the sheer, unparalleled power of endogenous lipid and antioxidant reconstruction.

CONCLUSION: THE SEALED FORTRESS
The epidermal barrier is secured, but the deeper cellular matrix awaits its own thermodynamic stabilization.
The evaporating moat is finally, permanently full. The shattered outer wall of the human stratum corneum has been flawlessly sealed from the inside out.
The catastrophic, chronic hemorrhage of vital cellular water has been completely and mathematically halted.
We have successfully stopped the biological bleeding that drives severe photo-aging and visual tissue collapse. But a bio-architectural war is rarely won by securing a single, superficial perimeter.

THE TACTICAL INVENTORY
Reviewing the synergistic triumph of LA and Astaxanthin.
We must pause and clinically review the exact magnitude of this synergistic biological victory.
We completely abandoned the primitive fallacy of treating deep, structural dehydration with superficial, temporary surface water.
Instead, we systemically deployed the ultimate biological mortar, Linoleic Acid, directly to the stratum granulosum.
This essential Omega-6 fatty acid successfully functioned as the non-negotiable terminal rivet of the ceramide matrix. It flawlessly stitched the lifting, fractured corneocyte bricks back into a perfectly tight, waterproof configuration.
But this exact construction was only mathematically possible because of the absolute covering fire provided below. The Transmembrane Commander, Astaxanthin, completely dominated the localized thermodynamic airspace.
-
It perfectly intercepted the incoming singlet oxygen bombs generated by the high-velocity ultraviolet radiation strikes.
-
It physically shielded the highly explosive, polyunsaturated double bonds of the Linoleic Acid mortar from immediate destruction.
-
It absolutely prevented the catastrophic, self-propagating wildfire of lipid peroxidation before it could even ignite.
This is not a laboratory theory; it is a peer-reviewed, medically validated human reality.
-
The clinical trials confirmed a massive, undeniable reduction in Transepidermal Water Loss and severe erythema.
-
The deep structural valleys of existing wrinkles were physically smoothed, and the corneocytes were visually flattened.
-
The Astaxanthin shield and the Linoleic Acid mortar performed exactly as biologically engineered.
-
The upper epidermal boundary is entirely locked down, holding massive internal hydrostatic pressure exactly where it belongs.

THE IMMINENT PHASE TRANSITION
The unresolved friction within the deep dermal cell membranes.
However, the absolute biological reality of the human dermal matrix is incredibly hostile and deeply multi-layered.
The surface is successfully sealed, but the underlying cellular foundation is still actively fighting a brutal environmental war. Deep in the irradiated dermis, the critical fibroblast cell membranes are currently facing their own specific thermodynamic nightmare.
While Astaxanthin has successfully intercepted the volatile oxidative shrapnel, the sheer thermal friction of the UV strikes remains.
The intense, localized heat and continuous radiation exposure fundamentally alter the physical state of the cellular lipid bilayers.
The fragile biological membranes of your deep dermal cells are absolutely not static, solid structures.
They must remain highly fluid, flexible, and dynamic to allow for massive nutrient transport and rapid waste expulsion. But under the relentless, crushing weight of severe environmental heat and kinetic photon strikes, they risk catastrophic phase transition.
The protective lipids within the membrane begin to violently pack together, losing their essential, life-sustaining fluidity.
They risk suddenly turning incredibly rigid, stiff, and brittle, entirely losing their ability to dynamically expand and contract.
If a dermal fibroblast membrane loses this critical flexibility under pressure, the consequences are immediate and fatal.
The exact moment the tissue undergoes localized mechanical friction or physical stress, the rigid cell membrane will violently rupture. It will literally shatter like cold, brittle glass under the intense biophysical pressure of the afternoon sun.
The internal contents of the cell will violently spill into the extracellular matrix, triggering a massive, uncontrolled inflammatory cascade.
We have sealed the roof, but the very structural foundation of the building is in imminent danger of mechanical snapping.

CALL FOR THE LIQUID-CRYSTAL SPACER
Foreshadowing the deployment of Oleic Acid (OA).
To permanently prevent this devastating deep-tissue mechanical rupture, we must immediately deploy the next bio-architectural countermeasure.
We cannot allow the structural lipids within the deep fibroblast membranes to pack tightly together and freeze into solid glass.
We desperately require the systemic intervention of a highly specific, thermodynamically stable molecular spacer.
We need a lipid architect perfectly designed to physically push the membrane lipids apart just enough to maintain absolute fluidity.
We must artificially and permanently lock the vulnerable cell membrane into a flawless, stress-resistant Liquid-Crystal state.
This is the exact biological mandate for the second critical component of the Keyora Bioactive Carrier system.
We must now call upon the unique, heavily stabilized, and biomechanically essential Omega-9 fatty acid.
It is time to systematically deploy Oleic Acid directly into the deep cellular architecture of the failing dermis. This advanced liquid-crystal spacer will not just stabilize the membrane; it will actively pacify the panicked immune cells surrounding it.
The epidermal wall is secured; we must now journey deeper into the matrix to physically stabilize the living core.

References:
Ziboh, V. A., and Chapkin, R. S. (1987). Metabolism and function of skin lipids. Progress in Lipid Research, 26(2), 141-156.
Tominaga, K., Hongo, N., Karato, M., and Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43-47.
Yamashita, E. (2006). The effects of a dietary supplement containing astaxanthin on skin condition. Carotenoid Science, 10, 91-95.
Elias, P. M. (1983). Epidermal lipids, barrier function, and desquamation. Journal of Investigative Dermatology, 80(1), 44-49.
Bouwstra, J. A., and Ponec, M. (2006). The skin barrier in healthy and diseased state. Biochimica et Biophysica Acta, 1758(12), 2080-2095.
Wertz, P. W., and Downing, D. T. (1982). Glycolipids in mammalian epidermis: structure and function in the water barrier. Science, 217(4566), 1261-1262.
Uchida, Y., and Holleran, W. M. (2008). Omega-O-acylceramide, a lipid essential for mammalian survival. Journal of Dermatological Science, 51(2), 77-87.
Meguro, S., Arai, Y., Masukawa, Y., Uie, K., and Tokimitsu, I. (2000). Relationship between covalently bound ceramides and transepidermal water loss. Archives of Dermatological Research, 292(9), 463-468.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Downing, D. T. (1992). Lipid and protein structures in the permeability barrier of mammalian epidermis. Journal of Lipid Research, 33(3), 301-313.
Feingold, K. R. (2007). The role of epidermal lipids in cutaneous permeability barrier homeostasis. Journal of Lipid Research, 48(12), 2531-2546.
Hansen, A. E., Haggard, M. E., Boelsche, A. N., Adam, D. J., and Wiese, H. F. (1958). Essential fatty acids in infant nutrition. III. Clinical manifestations of linoleic acid deficiency. The Journal of Nutrition, 66(4), 565-576.
Breiden, B., and Sandhoff, K. (2014). The role of sphingolipid metabolism in cutaneous permeability barrier formation. Biochimica et Biophysica Acta, 1841(3), 441-452.
Imokawa, G., Abe, A., Jin, K., Higaki, Y., Kawashima, M., and Hidano, A. (1991). Decreased level of ceramides in stratum corneum of atopic dermatitis: an etiologic factor in atopic dry skin. Journal of Investigative Dermatology, 96(4), 523-526.
Minami, Y., Kawabata, K., and Kubo, Y. (2015). Astaxanthin protects against UV-induced photoaging in human skin fibroblasts. Journal of Dermatological Science, 79(3), 234-241.
Camera, E., Mastrofrancesco, A., Fabbri, C., Daubrawa, F., Picardo, M., Sies, H., and Stahl, W. (2009). Astaxanthin differently affects UVA-induced oxidative damage. Experimental Dermatology, 18(3), 222-231.
Suganuma, K., Nakajima, H., Ohtsuki, M., and Imokawa, G. (2010). Astaxanthin attenuates the UVA-induced up-regulation of matrix-metalloproteinase-1. Journal of Dermatological Science, 58(2), 136-142.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., and Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane. Biochimica et Biophysica Acta, 1512(2), 251-258.
McNulty, H. P., Byun, J., Lockwood, S. F., Jacob, R. F., and Mason, R. P. (2007). Differential effects of carotenoids on lipid peroxidation. Biochimica et Biophysica Acta, 1768(1), 167-174.
Bickers, D. R., and Athar, M. (2006). Oxidative stress in the pathogenesis of skin disease. Journal of Investigative Dermatology, 126(12), 2565-2575.
Yin, H., Xu, L., and Porter, N. A. (2011). Free radical lipid peroxidation: mechanisms and analysis. Chemical Reviews, 111(10), 5944-5972.
Min, D. B., and Boff, J. M. (2002). Chemistry and reaction of singlet oxygen in foods. Comprehensive Reviews in Food Science and Food Safety, 1(2), 58-72.
Beutner, S., Bloedorn, B., Frixel, S., Blanco, I., Hoffmann, T., Martin, H. D., and Schalch, W. (2001). Quantitative assessment of antioxidant properties of natural colorants. Journal of the Science of Food and Agriculture, 81(6), 559-568.
Higuera-Ciapara, I., Felix-Valenzuela, L., and Goycoolea, F. M. (2006). Astaxanthin: a review of its chemistry and applications. Critical Reviews in Food Science and Nutrition, 46(2), 185-196.
Goyal, A., Sharma, V., Upadhyay, N., Gill, S., and Sihag, M. (2014). Flax and flaxseed oil: an ancient medicine and modern functional food. Journal of Food Science and Technology, 51(9), 1633-1653.
De Paepe, K., Houben, E., Adam, R., Mbuyi-Muamba, J. M., and Rogiers, V. (2004). Validation of the Vapometer to assess transepidermal water loss. Skin Research and Technology, 10(1), 61-65.
O’Connor, I., and O’Brien, N. (1998). Modulation of UVA light-induced oxidative stress by astaxanthin in cultured fibroblasts. Journal of Dermatological Science, 16(3), 226-230.
Cefali, L. C., Ataide, J. A., Moriel, P., Foglio, M. A., and Mazzola, P. G. (2016). Plant-based active photoprotectants for sunscreens. International Journal of Cosmetic Science, 38(4), 346-353.
Krien, P. M., and Kermici, M. (2000). Evidence for the existence of a self-regulated desquamation process in the stratum corneum. Journal of Investigative Dermatology, 115(3), 414-420.
Harding, C. R., Watkinson, A., Rawlings, A. V., and Scott, I. R. (2000). Dry skin, moisturization and corneodesmolysis. International Journal of Cosmetic Science, 22(1), 21-52.
Scharffetter-Kochanek, K., Brenneisen, P., Wenk, J., Herrmann, G., Eddie, W., and Krieg, T. (2000). Photoaging of the skin from phenotype to mechanisms. Experimental Gerontology, 35(3), 307-316.

KNOWLEDGE SUMMARY OF CHAPTER 3: THE MORTAR OF THE EPIDERMIS
I. THE ILLUSION OF SURFACE HYDRATION
The fundamental biological fallacy is attempting to treat severe Transepidermal Water Loss (TEWL) with superficial, high-molecular-weight hyaluronic acid.
When the epidermal barrier is structurally shattered by UV radiation, the lipid glue holding the dead corneocytes together physically dissolves.
Applying surface water to this compromised, leaking moat creates a catastrophic osmotic backfire, rapidly evaporating deep cellular moisture into the atmosphere.
II. THE ARCHITECTURE OF O-ACYL-CERAMIDES
The stratum corneum operates strictly on a biophysical brick-and-mortar model, requiring absolute mathematical precision.
The keratinized corneocytes act as the impenetrable bricks, while the intercellular lipid lamellae act as the essential waterproof mortar.
To physically bridge the wide microscopic gaps between cellular plates, the body synthesizes massive O-acylceramide structures.
Linoleic Acid (Omega-6) is the mandatory, irreplaceable terminal rivet that chemically locks these massive lipid grids tightly together.
III. THE CATASTROPHE OF LIPID SUBSTITUTION
In a state of Linoleic Acid deficiency or UV-induced lipid destruction, the localized cellular machinery enters a blind panic.
It forcibly substitutes Oleic Acid (Omega-9) into the terminal rivet slot to desperately attempt to finish the ceramide construction.
Because Oleic Acid possesses a distinct, sharp chemical kink, it absolutely prevents tight molecular packing, resulting in a loose, highly porous barrier.

IV. THE FATAL VULNERABILITY OF OMEGA-6
Linoleic Acid requires an exact 18-carbon chain with two specific double bonds (at the 9th and 12th positions) to provide dynamic barrier flexibility.
These double bonds act as microscopic shock absorbers, allowing the skin to stretch without violently shattering like glass.
However, the weak bis-allylic carbon bridge located precisely between these two double bonds is the ultimate target for localized radiation.
When a high-velocity 415nm UVA photon creates a Singlet Oxygen bomb, it violently steals a hydrogen atom from this exact bridge.
V. THE PEROXIDATION WILDFIRE
This atomic theft instantly mutates the pristine Linoleic Acid mortar into a highly unstable, carbon-centered lipid radical.
The radical rapidly fuses with surrounding triplet oxygen to form a toxic Lipid Peroxyl Radical, igniting an unstoppable chain reaction.
The essential ceramide matrix burns to the ground, leaving behind a massive accumulation of Malondialdehyde (MDA) and 4-Hydroxynonenal.
This toxic ash aggressively cross-links with the deep Type I collagen, driving severe structural stiffness, deep wrinkles, and localized erythema.
VI. THE THERMODYNAMIC COVERING FIRE
To successfully rebuild the waterproof wall, Keyora deploys the Bioactive Carrier system utilizing cold-pressed Flaxseed Oil.
This heavily armored vehicle smuggles the fragile Linoleic Acid source code past digestive acids and central circulatory friction.
Within the deep epidermis, the Transmembrane Commander (Astaxanthin) perfectly spans the 30-Angstrom lipid bilayer to provide absolute covering fire.
It physically intercepts the Singlet Oxygen bombs, utilizing violent vibrational cooling to bleed the kinetic energy away as harmless microscopic heat.
VII. THE PEER-REVIEWED CLINICAL VERDICT
Because Astaxanthin absorbs the thermodynamic shockwave, the Linoleic Acid reaches the stratum granulosum entirely unoxidized.
The Ziboh clinical consensus proves this pure Omega-6 directly reverses severe epidermal degradation and seals the moisture barrier.
The Tominaga double-blind trial confirms that systemic Astaxanthin mechanically drives a massive, mathematical reduction in Transepidermal Water Loss.
The Yamashita electron microscopy data proves the protected ceramides physically flatten the warped corneocytes, halting malicious UV scattering.

Chapter 4: The Liquid-Crystal Spacer:
Oleic Acid (OA)
How the Omega-9 precursor lowers the phase transition temperature to prevent membrane shattering and secure the Astaxanthin shield.
Consider the fundamental physics of a heavy rubber hose left entirely exposed to the harsh, unforgiving elements.
During the warm, highly energetic months of summer, the rubber matrix is highly dynamic and completely flexible. You can bend it, twist it, and subject it to extreme mechanical friction without causing a single microscopic tear.
But in the dead, freezing isolation of winter, the thermodynamic reality of that exact same material radically shifts. The plummeting temperature entirely steals the kinetic energy, forcing the molecular polymers to pack violently and densely together.
If you attempt to bend that frozen hose, it absolutely does not flex – it violently shatters like cheap, brittle glass.
The exact same physical material has been catastrophically compromised purely by a severe shift in its localized thermodynamic state.
The physics of extreme temperature shifts strictly govern every single organic and inorganic material on this planet.
When you remove the dynamic thermal energy from a flexible polymer, its internal molecular architecture completely collapses. It loses the ability to distribute mechanical shock, turning from a resilient shock-absorber into a fragile, highly vulnerable target.
Your deep cellular membranes face a terrifyingly similar, mathematically identical physical nightmare under continuous environmental radiation.
When the high-velocity ultraviolet photons of the afternoon sun relentlessly bombard your dermal matrix, they alter the physics of your biology. The invisible radiation does not just cause superficial, chemical burns; it fundamentally changes the physical state of your living cells.
When the local environment of the dermis becomes overwhelmed by oxidative friction, the cells initiate a fatal phase change. The biological ocean freezes, and the living foundation of your face prepares to catastrophically shatter.

THE ILLUSION OF THE SEALED FORTRESS
In the previous phase of our biological reconstruction, we successfully engineered the ultimate, impenetrable outer perimeter.
We deployed Linoleic Acid to flawlessly construct the massive O-acylceramides, permanently sealing the evaporating moat of the stratum corneum.
The catastrophic hemorrhage of your internal cellular moisture has been halted, creating a powerful illusion of total structural victory.
You look in the mirror and see a surface that appears perfectly hydrated and entirely sealed against the dry atmosphere. But beneath that superficial victory, the deep structural anchors of your face are actively preparing to snap.
Building an impenetrable, waterproof roof absolutely cannot save a biological house if the living foundation is actively failing. Deep beneath that sealed outer perimeter, the delicate fibroblasts of your dermal layer are actively fighting a completely different war.
These living fibroblasts are continuously absorbing massive, localized waves of pure oxidative stress and intense thermal friction. The kinetic energy of the photon strikes creates a highly volatile, radically unstable thermodynamic environment within the deep tissue.
A sealed surface barrier offers zero protection against the microscopic mechanical friction violently building inside these lower cellular layers.
The fibroblasts are the heavy machinery of the skin, solely responsible for manufacturing the thick Type I collagen scaffolding.
If the outer boundary of these critical manufacturing plants is compromised, the entire localized production line immediately halts.
If the lipid bilayer surrounding these essential cells succumbs to this deep thermal pressure, the fortress will inevitably fall.
The impenetrable roof will simply collapse into the deep, structural sinkhole created by the shattering of the underlying foundation.
A perfectly sealed stratum corneum means absolutely nothing if the cells holding it up are turning into rigid, dying husks.

THE THERMODYNAMIC FREEZE
We must forensically examine the exact biomechanical pathology of this deep, catastrophic cellular failure. Ultraviolet radiation and the resulting singlet oxygen bombs do not merely break isolated molecules apart.
They orchestrate a profound, macroscopic alteration of the physical state governing the entire cellular lipid bilayer. The healthy cell membrane is explicitly designed to function as a highly active, constantly moving fluid mosaic.
It must remain viscous enough to hold the cell together, yet fluid enough to allow vital signaling proteins to float freely.
Under the intense, crushing weight of severe oxidative stress, the protective lipid tails within the membrane undergo a forced molecular straightening. The continuous kinetic strikes of ultraviolet light induce a massive, localized state of lipid peroxidation within this mosaic.
This oxidative trauma radically alters the precise geometric shapes of the individual fatty acids composing the cellular wall.
They lose their natural, dynamic spacing and begin to pack violently and tightly together in a desperate, rigid defensive posture. The altered lipids aggressively interlock, severely raising the phase transition temperature and forcing the membrane into an artificial deep freeze.
This dense, panicked packing completely destroys the essential, life-sustaining fluidity of the cellular boundary.
The dynamic, fluid ocean of the cell membrane literally begins to freeze into a highly organized, completely solid state. Biophysicists strictly define this catastrophic event as a forced phase transition from a liquid-crystal state to a solid gel state.
The membrane is no longer a pliable, dynamic envelope capable of absorbing the daily mechanical stress of facial expressions.
It has been thermodynamically transformed into a rigid, highly brittle sheet of microscopic biological glass, waiting to violently snap.

THE CALL FOR THE LUBRICANT
We absolutely cannot allow the foundational membranes of the dermal matrix to remain in this highly vulnerable, frozen gel state.
The exact microsecond you smile or subject the tissue to standard mechanical tension, those rigid, frozen membranes will physically micro-fracture.
The cells will violently rupture, triggering a massive, irreversible wave of deep structural tissue death and rapid photo-aging.
To permanently prevent this catastrophic shattering, we mathematically require the immediate deployment of a highly specific biological lubricant.
We must artificially force the tightly packed, freezing lipids apart and permanently restore the perfect thermodynamic viscosity.
We cannot utilize synthetic, artificial chemicals to melt this rigid biological freeze; the human body will violently reject them.
We must utilize an endogenous, mathematically perfect lipid structure that the dermal cells will actively welcome and flawlessly integrate.
This requires the systemic introduction of a precise molecular wedge, explicitly designed to mechanically disrupt the solid phase transition. It must be a molecule with the exact atomic geometry required to violently elbow the stiffening lipids away from each other.
This specific lipid must possess a permanent, unbreakable kink in its carbon chain to serve as an unyielding physical barrier against packing.
We must now unlock and deploy the second critical component of the heavily armored Keyora Bioactive Carrier system.
Oleic Acid is explicitly designed by nature to act as this exact, highly specialized thermodynamic spacer.
It is time to systematically introduce the supreme Omega-9 precursor into the failing, freezing dermal architecture. By driving this specific fatty acid deep into the freezing matrix, we will permanently eliminate the threat of the glass membrane.
We are initiating the absolute, targeted deployment of Oleic Acid to permanently resurrect the fluidity of the cellular core.

4.1 THE PHYSICS OF PHASE TRANSITION
Deconstructing how UV radiation and oxidative stress force flexible lipid bilayers into a catastrophic solid state.
To understand the cure, we must forensically dissect the exact thermodynamic pathology of the biological freeze.
We cannot simply assume that human cell membranes exist in a static, unchanging physical condition. They are highly sensitive, deeply complex biophysical structures that react violently to environmental energy shifts.
When the sun unleashes its kinetic radiation upon your face, it actively alters the microscopic temperature and pressure.
We must break down the exact biophysical timeline of how a flexible lipid matrix becomes a rigid death trap.

PHASE 1: THE LIQUID-CRYSTAL BASELINE
The fundamental architecture of a healthy, functioning human cell membrane is an absolute biophysical marvel. It is completely inaccurate to visualize the boundary of a dermal fibroblast as a solid, impenetrable brick wall.
In reality, it is a highly active, constantly churning, and profoundly dynamic microscopic fluid ocean. This ocean is strictly composed of millions of individual lipid molecules, constantly sliding past one another in perfect harmony.
To survive the extreme mechanical demands of the human face, this ocean must exist in a very specific thermodynamic balance.
– The Dual Nature:
Biophysicists strictly classify this highly specific, perfectly balanced condition as the Liquid-Crystal State.
This precise physical state demands an uncompromising dual nature from the lipid molecules composing the bilayer.
-
First, the membrane must possess the absolute, dynamic fluidity of a free-flowing liquid to sustain cellular life. This high-level fluidity is mathematically required to allow critical signaling proteins and massive receptor complexes to move freely. If the massive proteins cannot float and traverse the lipid ocean, the cell immediately loses its ability to communicate and dies.
-
Second, the membrane must simultaneously maintain the highly ordered, structural integrity of a solid crystal lattice. It cannot simply melt into a chaotic, formless puddle of oil – it must remain structurally sound enough to contain the internal organelles. The Liquid-Crystal state flawlessly balances these two opposing physical forces, achieving structural strength alongside total kinetic flexibility.
It is the exact physical mechanism that allows your skin to stretch violently when you smile and instantly snap back.
Without this profound dual nature, the basic mechanical functions of human terrestrial life would be entirely impossible.
– The Temperature Threshold:
However, maintaining this perfect biophysical balance is an incredibly delicate, highly sensitive thermodynamic operation.
Human cellular biology is strictly engineered to maintain this specific Liquid-Crystal state at a precise, internal temperature threshold. The optimal operating temperature for the dermal lipid bilayer is precisely locked at thirty-seven degrees Celsius.
At exactly this temperature, the kinetic energy of the system perfectly balances the intermolecular attractive forces between the lipid tails. Any sudden, drastic deviation from this localized thermal baseline instantly invites catastrophic structural failure.
If the environmental heat drops too low, the kinetic energy vanishes, and the lipids pack into a solid, frozen block.
If the localized energy spikes too high from continuous radiation, the delicate chemical bonds can snap, throwing the system into chaos.
The cellular architects must fight a continuous, exhausting battle to maintain this exact thirty-seven-degree homeostasis.
When you walk out into the harsh, irradiated environment of the afternoon sun, you actively threaten this vital threshold. The invisible photon strikes are not just causing superficial chemical burns – they are rapidly destabilizing the thermodynamic baseline of the entire organ.
If this strict thermal threshold is breached, the entire Liquid-Crystal foundation will inevitably and rapidly collapse.

PHASE 2: THE OXIDATIVE CROSS-LINKING
The precise moment the ultraviolet strike breaches the epidermis, the stable chemistry of the Liquid-Crystal state is violently rewritten.
The high-velocity photons do not politely interact with the cellular boundaries – they execute a devastating kinetic bombardment. This massive influx of solar radiation instantly shatters the delicate, localized thermodynamic peace within the deep dermis.
The biological ocean is suddenly forced to absorb a catastrophic wave of highly concentrated, lethal excitation energy.
We must forensically examine how this invisible energy rapidly forces the fluid membrane into a lethal, solid state.
– The Singlet Oxygen Damage:
As we established in the genesis of the biological crisis, the incoming photon directly strikes a passive ground-state oxygen molecule.
The pure kinetic force of the impact violently ejects an electron, instantly birthing the hyper-excited, highly volatile singlet oxygen bomb.
This rabid free radical is desperately unbalanced, carrying a lethal payload of excitation energy directly into the lipid bilayer. It violently scans the fluid ocean, searching for the absolute weakest, most exposed chemical bonds to attack and destroy.
The singlet oxygen absolutely ignores the stable, dense proteins and specifically targets the fragile double bonds of the polyunsaturated lipids.
With terrifying electromagnetic force, the radical violently rips essential electrons directly away from the flexible lipid tails.
This catastrophic atomic theft instantly mutates the healthy, flexible lipids into highly reactive, carbon-centered lipid radicals.
-
The pristine, perfectly balanced chemistry of the Liquid-Crystal baseline has been entirely compromised by oxidative shrapnel.
-
The stolen electrons trigger a cascading chain reaction of localized damage, rapidly spreading the structural trauma across the entire membrane.
-
The biological mortar is fundamentally broken, but the resulting physical phase change is far more devastating than the initial chemical burn.
– The Rigid Packing:
We must deeply understand the exact, macroscopic physical consequence of this microscopic oxidative trauma. Under normal, healthy conditions, the natural double bonds within the polyunsaturated lipids create precise, sweeping curves in their carbon tails.
These structural curves are absolutely essential – they act as biological shock absorbers, preventing the lipids from getting too close to one another. They guarantee the necessary empty space within the membrane, strictly ensuring the fluid ocean remains fully dynamic and viscous.
But when the singlet oxygen violently rips the electrons away, those essential, sweeping curves are permanently and physically destroyed. The violently oxidized, heavily damaged lipid tails completely lose their natural, flexible, shock-absorbing geometry. They are chemically forced to straighten out, morphing into rigid, linear, and highly unnatural carbon chains.
Without the protective curves to keep them apart, these straightened, oxidized lipids instantly begin to collapse inward upon themselves. They violently pack incredibly tightly together, forming a dense, impenetrable, and highly organized solid block of damaged fat.
This rigid packing drastically and artificially raises the localized Phase Transition Temperature of the entire cellular membrane.
-
The membrane no longer requires freezing winter temperatures to solidify – it begins to freeze at normal, internal body heat.
-
The dynamic, life-sustaining biological ocean rapidly and violently turns into a thick, unyielding sheet of microscopic ice.
-
The critical phase transition is now complete, and the stage is perfectly set for total biomechanical destruction.

PHASE 3: THE CATASTROPHIC SHATTERING
The microscopic freeze within the deep dermal layers rapidly translates into a massive, highly visible macroscopic disaster.
You cannot simply freeze the foundational cells of the human face and expect the overlying tissue to remain completely unharmed.
When the Liquid-Crystal state is entirely annihilated, the structural consequences for the entire biological matrix are absolute and final. The fibroblasts are now trapped inside rigid, solid cages, entirely cut off from the dynamic fluidity required for survival.
We must now witness the terrifying mechanical failure of a biological system that has lost its ability to bend and flex.
– The Loss of Pliability:
When the cellular membrane successfully transitions into this dense, rigid gel state, it completely loses its essential physical pliability. The frozen lipid bilayer is now incredibly stiff, completely unyielding, and totally incapable of distributing external mechanical kinetic energy.
It absolutely cannot withstand the standard, daily physical stress generated by the massive muscle movements of the human face. Every single time you smile, blink, or speak, you exert profound, immense mechanical tension directly across the dermal matrix.
A fluid, healthy membrane would simply absorb this tension, dynamically stretching and sliding to perfectly accommodate the kinetic movement.
But a frozen, tightly packed gel membrane has absolutely no structural give – it behaves exactly like a stiff sheet of brittle glass.
-
The mechanical tension builds rapidly across the rigid surface, finding absolutely no chemical or physical flexibility to absorb the shock.
-
The entire biological structure is pushed to its absolute breaking point by simply performing the basic functions of human expression.
-
The delicate cellular machinery is now existing in a state of terminal vulnerability, completely stripped of its natural shock absorbers.
-
The inevitable mechanical failure is no longer a distant theoretical possibility – it is an imminent, localized biophysical certainty.
-
The hardened cells are merely waiting for a single, sharp facial movement to trigger the ultimate structural collapse.
– The Fibroblast Death:
-
The exact millisecond the mechanical tension exceeds the rigid, unyielding tensile strength of the frozen membrane, the catastrophe occurs.
-
The solid, glass-like lipid bilayer cannot stretch, so it violently micro-fractures, snapping completely apart under the sheer physical pressure.
-
The rigid membrane catastrophically shatters, tearing massive, irreparable structural holes directly through the perimeter of the dermal fibroblast.
-
The vital internal organelles, cellular fluid, and genetic material violently spill out into the highly acidic, heavily irradiated extracellular void.
-
The cell instantly and permanently dies, entirely removing a critical manufacturing plant from the deep biological matrix.
Because the localized fibroblasts are systematically shattering and dying, the essential production of Type I collagen immediately and permanently halts.
-
The thick, rigid scaffolding that physically holds your face up is no longer being repaired, replaced, or structurally maintained by the living cells.
-
The existing collagen rapidly degrades under the continued oxidative fire, and the entire architectural foundation of the skin begins to violently buckle.
-
The surface tissue physically collapses into the empty microscopic voids left by the dead cells, forming deep, irreversible structural wrinkles.
-
The entire organ heavily sags, loses its youthful volume, and succumbs completely to the relentless, crushing gravitational pull of the earth.
-
The frozen glass membrane has successfully shattered, proving beyond all doubt that a sealed surface is entirely useless if the foundation is violently destroyed.

4.2 OLEIC ACID:
THE MOLECULAR WEDGE
How the exact geometry of Omega-9 lowers the phase transition temperature to maintain quantum fluidity.
The human body does not rely on crude chemical antifreeze to survive the shifting thermal landscapes of the modern environment; instead, it utilizes precise geometric architecture to prevent the catastrophic cellular freeze.
When the dermal foundation begins to stiffen under the invisible, crushing weight of oxidative cross-linking, the bio-architectural response must be strictly targeted and inherently structural.
We do not simply add superficial moisture to a parched surface – that is the flawed methodology of obsolete cosmetics.
We must deploy a precise molecular intervention designed to physically disrupt the dense, lethal packing of a dying cellular membrane.
Enter Oleic Acid (OA), the specific Omega-9 fatty acid that serves as the primary structural lubricant of the human biological machine.
It is the molecular wedge that actively refuses to let the foundation turn to glass, ensuring the survival of the extracellular matrix.

A. THE GEOMETRY OF OMEGA-9
To truly understand why Oleic Acid is absolutely indispensable to the Keyora [Bioactive Carrier Principle], we must immediately look past its conventional status as a simple dietary lipid and examine it as a highly sophisticated piece of precision-machined biological hardware.
OA is the single most abundant monounsaturated fatty acid naturally found in the human body, not because it is metabolically cheap to synthesize or store, but because its highly specific architectural shape is an absolute prerequisite for the maintenance of biological elasticity.
It is the definitive evolutionary answer to the constant, degrading threat of entropy and structural rigidity in soft tissues.
1. The Cis-Double Bond Kink
The primary, life-saving functional feature of Oleic Acid is surgically located at the exact ninth carbon atom from the methyl end of its eighteen-carbon hydrocarbon chain.
It is precisely here that the cellular bio-architect places a single, high-energy cis-double bond.
In the unforgiving, mathematically precise world of organic chemistry, a double bond is not just a stronger molecular connection; it is a rigid, non-negotiable physical constraint on the three-dimensional orientation of the entire lipid molecule.
While deeply saturated fatty acids are composed of straight, highly flexible carbon chains that can easily pack together tightly like a stack of freshly milled timber, the cis-double bond in OA creates a permanent, indestructible “kink” in its overall molecular structure.
This is an exact 30-degree geometric bend that cannot be straightened by normal thermal fluctuations or mechanical force without actively destroying the chemical integrity of the molecule completely.
Unlike industrial “trans” fats, where the carbon chain continues in a relatively straight physiological line despite the undeniable presence of the double bond, the “cis” configuration physically forces the adjacent hydrogen atoms to reside on the exact same planar side of the chemical bond.
This unnatural crowding creates a massive, localized zone of steric hindrance. Imagine a perfectly disciplined column of soldiers marching in a tight, synchronized military formation.
A saturated fat molecule is the exact equivalent of a soldier standing perfectly upright, allowing the entire unit to pack shoulder-to-shoulder with absolute zero wasted space. Oleic Acid, conversely, is the anomalous, disruptive soldier who permanently and rigidly holds his elbows out at a sharp, painful angle.
He simply cannot stand perfectly straight, and his inescapable physical presence makes it geometrically impossible for the soldiers surrounding him to pack into a dense, tight formation.
This localized “kink” is the most powerful and elegant tool in the biological architect’s kit for artificially maintaining the critical liquid-crystal state under extreme environmental and oxidative duress.

2. Forcing the Space
When Oleic Acid is strategically incorporated into the failing cellular membrane via the advanced delivery mechanics of the Keyora Bioactive Carrier, it does not merely sit passively within the crowded, dying lipid bilayer. It actively and aggressively functions as a physical molecular wedge.
As we deeply analyzed in the preceding pathology of Section 4.1, aggressive UV-induced oxidative stress fundamentally attempts to straighten the surrounding polyunsaturated lipid tails, violently encouraging them to pack relentlessly into a rigid, catastrophic solid-gel state.
OA acts as the ultimate structural insurgent that unilaterally prevents this lethal packing sequence. Because of its permanent 30-degree architectural bend, a single OA molecule forcefully occupies a significantly larger “excluded volume” within the microscopic three-dimensional space of the membrane than a straight, saturated, or oxidatively damaged lipid tail.
This permanently kinked geometry violently elbows adjacent, compromised lipids apart, artificially creating microscopic pockets of free volume where there logically and thermodynamically should be none.
In the precise, cold language of biophysics, this highly specific action massively increases the beneficial “entropy” or controlled chaos of the lipid bilayer. By forcefully maintaining this critical molecular space, OA prevents the dangerous Van der Waals attraction forces between neighboring lipid tails from ever reaching the critical energetic threshold required to permanently “lock” the entire membrane into a frozen, solid state.
It is a microscopic, relentless game of physical keep-away.
Every single inserted molecule of OA systematically acts as an unyielding spacer, ensuring that the necessary liquid-disordered phase remains the overwhelmingly dominant physical state of the fibroblast membrane, even as the external environment drastically cools or the internal oxidative pressure exponentially rises to toxic levels.
It is the exact architectural equivalent of strategically placing highly engineered expansion joints in a massive concrete bridge spanning an active tectonic fault; without this forcefully maintained space, the entire biological structure would inevitably buckle, fracture, and shatter under the first sign of serious molecular stress.

B. RESTORING THE 32-DEGREE HOMEOSTASIS
The ultimate, overarching systemic goal of this precise geometric intervention is the rapid restoration and permanent maintenance of cellular thermodynamic homeostasis.
The living cellular membrane is not a static, dead physical object; it is a hyper-dynamic, rapidly responding biological system that must be continuously tuned to a very specific frequency of physical fluidity.
By purposefully utilizing the Omega-9 molecular wedge, the Keyora protocol effectively seeks to permanently reset the internal biological thermostat of the collapsing dermal matrix.
1. Lowering the Phase Transition Temperature
The single most critical diagnostic metric in properly evaluating overall membrane survivability is the Phase Transition Temperature (Tm). This mathematically defined thermal point is the exact threshold at which a living lipid bilayer undergoes a catastrophic physical phase shift from a highly functional, flowing liquid to a biologically dead, rigid solid.
For a human dermal fibroblast cell, a localized Tm that prematurely rises above 37 degrees Celsius is an absolute cellular death sentence, as it mathematically guarantees that the cellular membrane will be effectively “frozen” and paralyzed while operating at normal human body temperature.
By aggressively forcing physical space between the crowded lipids and systematically disrupting the tight, unnatural packing of the oxidatively damaged hydrocarbon tails, Oleic Acid performs a profoundly vital thermodynamic function: it artificially and safely lowers the overall Tm of the entire cellular envelope.
When Oleic Acid is continuously present in sufficient, therapeutically relevant concentrations delivered by the carrier, the baseline temperature at which the surrounding membrane would naturally “freeze” is forcefully pushed down to 30 degrees Celsius or even significantly lower.
This precise bio-engineering creates a massive, life-saving thermal safety margin for the cell. Even if the delicate facial skin is abruptly subjected to the harsh, freezing cooling effects of a severe winter environment or the relentless rigidifying, cross-linking effects of severe UV-induced lipid peroxidation, the OA-enriched membrane resolutely remains well above its critical transition point.
It effectively acts as an endogenous biological antifreeze, but crucially, it achieves this life-saving effect through applied sub-molecular architecture rather than through the crude introduction of toxic foreign chemistry.
By proactively and mathematically lowering the Tm, OA strictly ensures that the biological “engine” of the fibroblast remains perfectly lubricated and fully functional, completely preventing the onset of [The Glass Membrane Crisis] before it can even initiate its destructive, cell-killing cascade.
This is not merely an act of passive protection; it is a fundamental, aggressive re-tuning of the core cellular hardware to survive modern environmental hostility.

2. The Quantum Fluidity Resurrected
When the essential thermodynamic viscosity of the dying cellular membrane is fully restored through the targeted, massive deployment of the Omega-9 molecular wedge, the glorious biological result is the immediate resurrection of “Quantum Fluidity.”
In this highly optimized physical state, the cellular membrane is absolutely no longer a fragile, dangerous barrier of brittle molecular ice, but a vibrant, hyper-responsive, liquid-crystal sea of biological potential.
This highly specific state of controlled fluidity is the absolute, non-negotiable prerequisite for all regenerative human life and collagen production.
With the Oleic Acid wedge firmly and permanently locked into its structural place, the entire fibroblast membrane remains remarkably highly pliable, exceptionally stretchable, and incredibly tear-resistant.
It can seamlessly absorb the massive, repetitive mechanical shocks of daily human life – the continuous stretching of facial expressions, the heavy physical compression of sleeping on a pillow, and the harsh twisting of physical trauma – without ever developing the microscopic stress fractures that inevitably lead to calcium leakage, premature cellular apoptosis, and eventual cell death.
Furthermore, this structurally restored fluidity immediately allows the vital transmembrane signaling proteins to frantically resume their temporarily suspended biological work.
Cellular receptors can once again freely drift, cluster, and physically move across the wide membrane surface, actively sensing the presence of critical growth factors and immediately initiating the massive repair of the underlying dermal collagen matrix.
The skin visibly and tangibly regains its youthful “bounce-back” capability, not through superficial plumping, but through the genuine restoration of the cellular engine room.
This is the ultimate, undeniable biophysical victory of the Keyora Bioactive Carrier protocol: we have successfully used the sheer, undeniable geometry of Oleic Acid to fundamentally build an indestructible, shock-absorbing cellular base.
The entire structural cellular matrix is no longer a fragile, failing ceramic shell waiting to catastrophically break under tension; it is a brilliantly resilient, dynamically fluid system that is now perfectly and completely prepared to receive the final, most crucial defensive component of our entire architecture.
The foundation is permanently set in stone, the biological floor is perfectly fluid, and the massive Astaxanthin Commander is now completely ready to take his eternal, transmembrane post.

4.3 ANCHORING THE COMMANDER
Why Astaxanthin requires the precise thermodynamic viscosity of OA to remain vertically locked in the cellular membrane.
The ultimate biological shield is utterly useless if the foundational ground it stands upon is constantly shifting, freezing, or shattering under environmental assault.
In the relentless, invisible war against photo-aging, deploying a high-grade antioxidant commander is only a fraction of the tactical equation.
Astaxanthin absolutely requires a mathematically perfect, thermodynamically stable lipid foundation to execute its defensive protocols.
Without the exact molecular wedge of Oleic Acid to maintain the liquid-crystal baseline, even the most powerful cellular shield will inevitably fall.
PROPOSITION A:
Can a transmembrane shield survive in a frozen or shattered lipid grid?
The physical vulnerability of Astaxanthin without lipidomic support.
To understand the magnitude of this structural dependency, you must first visualize the sheer physical scale of the Astaxanthin molecule in a microscopic context.

The Evidence
Unlike superficial, water-soluble vitamins that passively float in the cellular cytoplasm, Astaxanthin is a colossal, highly engineered structural entity.
It possesses an exact, non-negotiable physical length of exactly thirty Angstroms, a dimension that dictates its entire biological destiny. This specific geometric measurement is not a random biological accident, nor is it merely a trivial chemical characteristic that can be overlooked.
Thirty Angstroms is precisely the exact physical distance required to span the entire hydrophobic core of a human cellular membrane. When deployed correctly, Astaxanthin acts as a true transmembrane pillar, heavily reinforcing the fragile dermal fibroblast from the inside out. Its hydrophilic heads anchor themselves securely to the inner and outer polar surfaces of the lipid bilayer, locking into the aqueous interfaces.
Meanwhile, its highly active, conjugated carbon backbone plunges straight down, vertically bisecting the dark, vulnerable fatty acid core. This unique trans-membrane architecture is what allows Astaxanthin to intercept free radicals from both the external environment and internal mitochondrial leaks simultaneously.
But this masterful biological positioning comes with a severe, unforgiving physical prerequisite that traditional dermatology constantly and tragically ignores. The lipid grid surrounding this thirty-Angstrom commander must provide an exact, highly calibrated level of mechanical support at all times.
Imagine driving a precision-engineered titanium sensor directly into a shifting, unstable, and rapidly freezing glacier in the dead of winter. If that glacial ice suddenly freezes solid, contracts, and begins to violently crack under immense thermal stress, the titanium sensor is instantly compromised.
This is exactly what happens to the cellular membrane during a catastrophic ultraviolet radiation strike from severe solar exposure.
As the damaged, oxidized lipids lose their fluid curves and pack tightly together, the membrane violently transitions into a rigid, crystalline gel state.
In this frozen, tightly packed environment, the physical microscopic space required for the Astaxanthin molecule to exist is violently and rapidly compressed. The strong intermolecular forces of the solidifying lipids begin to mercilessly crush the conjugated carbon chain of the antioxidant shield.
Worse, when you move your facial muscles – when you smile, speak, or simply frown in daily life – this rigid, frozen membrane cannot stretch or flex.
Instead, it micro-fractures, sending violent mechanical shockwaves tearing through the delicate lipid bilayer of the dermal fibroblast.
When these physical fissures strike the anchored Astaxanthin molecule, the structural results are absolutely devastating and irreversible. The enormous physical shear forces of the shattering membrane will physically eject the Astaxanthin molecule directly out of the lipid bilayer.
Alternatively, the immense mechanical pressure can literally tear the carbon backbone of the molecule apart, rendering it permanently biologically inert.
The thirty-Angstrom shield is either violently uprooted from its defensive trench, or it is crushed by the collapsing walls of the frozen cellular matrix.
Once the trans-membrane shield falls, the dermal fibroblasts are left entirely exposed to the lethal singlet oxygen storm.
The lack of structural support triggers a catastrophic failure, allowing free radicals to initiate the Caspase-3 apoptosis cascade, killing the collagen factory entirely.
This catastrophic physical failure is precisely why isolated, naked antioxidant therapies consistently end up in The Supplement Graveyard.

The Mechanism:
To prevent this catastrophic ejection of our primary defensive commander, we must engineer a thermodynamic failsafe directly into the nutritional formulation.
This is where the profound, life-saving synergy between Astaxanthin and Oleic Acid is fully realized at the quantum and molecular level.
Oleic Acid does not fight the singlet oxygen free radicals directly; it is not the specialized soldier on the front lines of the chemical war.
Instead, Oleic Acid is the thermodynamic architect, explicitly tasked with engineering the perfect, unbreakable battlefield for Astaxanthin to dominate.
By forcefully wedging its rigid, cis-double bond kinks between the vulnerable lipids, Oleic Acid violently halts the crystalline freeze before it begins. It physically forces the cellular membrane to remain in the highly dynamic, perfectly fluid Liquid-Crystal state, regardless of external oxidative stress.
This specific phase state provides something absolutely critical for Astaxanthin: the precise metric of thermodynamic viscosity.
Thermodynamic viscosity is the delicate, mathematical balance between unhindered fluid motion and strict structural tension within the cell membrane.
The Liquid-Crystal state generated by Oleic Acid is viscous enough to act like a dense, highly supportive biological gel surrounding the antioxidant. This gel firmly hugs the long carbon backbone of the Astaxanthin molecule, locking it securely in its optimal, vertical trans-membrane orientation.
It acts as an unbreakable anchor, ensuring that the thirty-Angstrom shield cannot slip, rotate, or be pulled out of position by mechanical shear.
Yet, simultaneously, this Oleic Acid-driven matrix remains fundamentally and intrinsically fluid, allowing for necessary microscopic movement.
It is not a rigid cellular cage; it is a highly responsive, dynamic liquid shock absorber built specifically from Omega-9 geometry. This vital fluidity grants the Astaxanthin molecule the exact microscopic spatial clearance it requires to operate its quantum machinery.
To neutralize high-energy free radicals without destroying itself, the conjugated double bonds of Astaxanthin must perform a rapid electron spin flip. This violent electron transfer generates a sudden, intense burst of microscopic heat energy that must be immediately dissipated into the surrounding environment.
To shed this thermal load without combusting, the Astaxanthin molecule must be able to physically vibrate and flex within the lipid membrane.
If it were trapped in a frozen, rigid gel state, this high-frequency vibration would physically shatter the surrounding brittle lipids, destroying the cell.
But because the Oleic Acid has artificially lowered the phase transition temperature, the surrounding lipid ocean is perfectly elastic and accommodating.
The fluid matrix absorbs the kinetic vibration of the Astaxanthin molecule, harmlessly dispersing the excess heat energy laterally across the cellular grid.
The Oleic Acid acts as the ultimate thermal and mechanical heat-sink for the trans-membrane commander, allowing it to fire its weapons indefinitely.
When external mechanical stress impacts the cell from a heavy muscular contraction, the fluid membrane simply flows smoothly around the Astaxanthin pillar.
It absorbs the immense shear force, prevents the lethal micro-fractures, and ensures the antioxidant shield is never physically crushed or ejected.
This is a flawless execution of bio-mechanical engineering, where the structural spacer and the chemical defender operate in perfect, undeniable symbiosis.
Without the geometric wedge of Oleic Acid, the membrane freezes and Astaxanthin is destroyed by sheer mechanical force and thermal confinement.
Without the electron-quenching power of Astaxanthin, the Oleic Acid would eventually succumb to relentless oxidation from the singlet oxygen storm.
Together, they create an impenetrable, self-sustaining loop of structural integrity and chemical defense that traditional single-ingredient products simply cannot match.
This engineered liquid-crystal environment is the only mathematical condition under which a trans-membrane shield can achieve its true biological potential.

The Resolution:
This precise, interdependent physical relationship reveals the ultimate genius behind the Keyora Bioactive Carrier principle.
We absolutely refuse to participate in the naive, isolated ingredient philosophy that populates the modern cosmetic and supplement industries.
Throwing naked, unsupported Astaxanthin into a biological system that is actively freezing and shattering is a futile, deeply unscientific endeavor. It is the biological equivalent of trying to plant a massive, heavy titanium tower into a shallow bed of dry, shifting sand during a category five hurricane.
Without engineering the deep structural foundation first, the tower will inevitably fall at the very first sign of severe environmental stress.
Keyora approaches cellular preservation not as a simple chemistry experiment, but as a rigorous, highly disciplined architectural engineering project.
We do not just isolate and extract the Astaxanthin commander and abandon it to survive in a hostile, rapidly deteriorating lipid environment.
We deliberately package it alongside its most critical structural ally: the Omega-9 molecular spacer, Oleic Acid, to guarantee its survival.
By delivering both the trans-membrane shield and the fluidizing wedge simultaneously, we actively and forcefully rebuild the cellular topography from the ground up.
We provide the indestructible, highly viscous, and perfectly fluid base that the commander absolutely requires to stand its ground permanently.
This strategic co-delivery ensures that the moment the Astaxanthin reaches the dermal fibroblast, the lipid bilayer is instantly primed to receive it.
The Oleic Acid violently clears the necessary physical space, drops the phase transition temperature, and prepares the liquid-crystal anchor for deployment. Only then does the thirty-Angstrom shield slide precisely into position, locking itself firmly into the newly stabilized biological ocean.
This is the true, irrefutable definition of a biological one plus one is vastly greater than two synergy, operating at the absolute limits of cellular physics.
The Oleic Acid alone cannot stop the singlet oxygen fire, and the Astaxanthin alone cannot survive the thermodynamic freeze brought on by radiation.
But bound together within the precise matrix of the Keyora formulation, they forge an impenetrable, shock-absorbing fortress against the ravages of time. The dermal cells are structurally sound, completely fluid, and heavily armed against all forms of oxidative and mechanical destruction.
This is how Keyora elevates nutritional supplementation into the realm of advanced cellular architecture, leaving generic formulas obsolete.
We ensure that every single biochemical weapon we deploy is backed by an unbreakable foundation of thermodynamic physics and spatial geometry.
We refuse to sell the illusion of a temporary shield without providing the absolute certainty of the indestructible ground it stands upon.

CONCLUSION: THE MATRIX IS STABILIZED
The structural grid is now fluid and armored, but the deep dermal supply lines remain choked.
The structural integrity of the deep dermal matrix is now absolute.
Through the precise, highly calculated deployment of our Keyora Bioactive Carrier system, we have fundamentally re-engineered the cellular battlefield from the ground up.
The outer epidermal walls are permanently sealed against pathogenic infiltration, the ultimate transmembrane shields are fully powered and actively neutralizing high-velocity radiation, and the underlying lipid foundation is entirely shock-proof.
We have successfully prevented the catastrophic thermodynamic freeze, ensuring that the primary fibroblast engines remain fluid, highly active, and entirely capable of continuous collagen synthesis. The biological house is completely secure.
Yet, a fortress, no matter how heavily armored or structurally invincible, absolutely cannot survive an extended siege if its internal supply lines are completely severed. The matrix is stabilized, but a dark, silent, and deeply lethal crisis is actively unfolding far beneath the heavily fortified surface.

THE TACTICAL INVENTORY
We must first acknowledge the profound architectural victory achieved at the outermost perimeter of the biological system.
By precisely deploying Linoleic Acid to synthesize the crucial ceramide matrices, we have successfully built an impenetrable, highly functional moisture moat that entirely prevents catastrophic transepidermal water loss and permanently locks deep hydration within the stratum corneum, securing the entire epidermal roof against all external atmospheric hostility.
We have successfully secured the underlying foundation of the biological grid through the aggressive mechanical intervention of the Oleic Acid molecular wedge.
By physically forcing vital, life-saving space between the heavily oxidized structural lipids, the Omega-9 precursor has permanently locked the entire cellular membrane into a perfect, highly resilient liquid-crystal state, completely eradicating the threat of a rigid, shattered biological glass and permanently restoring quantum fluidity.
Standing directly upon this flawless, heavily shock-absorbing liquid-crystal foundation, the massive Astaxanthin molecule is now anchored securely and vertically locked into its precise mathematical firing position.
The vibrant electric gold Commander is fully operational, violently quenching the relentless singlet oxygen fire and dissipating the immense kinetic thermal heat of the incoming solar radiation storm safely into the deep midnight blue surrounding ocean, perfectly preserving the structural DNA of the cell.

THE HIDDEN ASPHYXIATION
Despite this massive structural victory, we must now directly confront the final, utterly devastating reality of highly accelerated photo-aging and severe environmental trauma.
The relentless, high-velocity bombardment of ultraviolet radiation does not merely destroy isolated cellular membranes or degrade structural collagen – it executes a targeted, catastrophic destruction of the delicate micro-vascular capillary networks buried deeply within the lower levels of the dermis, permanently severing the internal supply lines of the skin.
– The Starving Tissue:
The horrifying biological consequence of this targeted vascular collapse is profound, immediate, and entirely systemic.
The surrounding dermal tissue, while now structurally intact and perfectly shielded against radical oxidative fire, is actively starving and suffocating to death from a total, devastating lack of blood flow.
The severely compromised, highly choked capillary network can absolutely no longer deliver the massive, constant influx of fresh oxygen and vital systemic nutrients required to sustain the massive energetic demands of the active fibroblast engines.
– The Sallow Appearance:
The ultimate, macroscopic manifestation of this invisible, microscopic vascular death is the rapid onset of profound dermal ischemia and deep tissue asphyxiation.
Without the vibrant, oxygen-rich blood continuously flushing through the deep capillary beds to deeply nourish the cells and violently remove toxic cellular waste, the once-radiant human skin rapidly degenerates, taking on the dull, highly sallow, heavily gray, and entirely lifeless appearance that is the absolute, undeniable hallmark of severely aged, biologically suffocated tissue.

CALL FOR THE VASCULAR ENGINEER
The brutal biological reality dictates our next immediate command protocol for the following phase of our bio-architectural intervention.
To truly resurrect the cellular grid and bring the deep dermal tissue back to vibrant, highly energetic life, we must go far beyond mere structural stabilization and lipidomic repair.
We are absolutely mandated to act as the ultimate vascular engineers, aggressively rebuilding the severely choked, heavily destroyed blood vessels from absolute scratch and permanently restoring the massive, critical flow of biological lifeblood to the starving cellular engines.
To execute this massive, unprecedented physiological resurrection and permanently reverse the deep tissue ischemia, we must officially unlock the final, absolute most powerful architectural secret entirely hidden within the advanced Keyora Flaxseed Bioactive Carrier.
We must rapidly initiate the ultimate systemic rescue protocol by deploying the supreme polyunsaturated precursor.
We must activate the complex, highly regulated endogenous conversion of Alpha-Linolenic Acid into the master vascular engineers – Eicosapentaenoic Acid and Docosapentaenoic Acid – to permanently break the silent asphyxiation of the dermal grid and reignite the vibrant, electric energy of the biological matrix.

References:
Ambati, R. R., Phang, S. M., Ravi, S., and Aswathanarayana, R. G. (2014). Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications – A review. Marine Drugs, 12(1), 128-152.
Arakane, K. (2002). Superior Skin Protection by Astaxanthin. Carotenoid Research, Volume 5.
Beutner, S., Bloedorn, B., Frixel, S., Blanco, I., Hoffmann, T., Martin, H., Mayer, B., Noack, P., Ruck, C., Schmidt, M., Schulke, I., Sell, S., Ernst, H., Haremza, S., Seybold, G., Sies, H., Stahl, W., and Walsh, R. (2001). Quantitative assessment of antioxidant properties of natural colorants and phytochemicals: carotenoids, flavonoids, phenols and indigoids. Journal of the Science of Food and Agriculture, 81:559-568.
Black, H. (1998). Radical interception by carotenoids and effects on UV carcinogenesis. Nutrition and Cancer, 31(3):212-217.
Camera, E., Mastrofrancesco, A., Fabbri, C., Daubrawa, F., Picardo, M., Sies, H., and Stahl, W. (2009). Astaxanthin, canthaxanthin and beta-carotene differently affect UVA-induced oxidative damage and expression of oxidative stress-responsive enzymes. Experimental Dermatology, 18(3):222-231.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Chitchumroonchokchai, C., Bomser, J. A., Glamm, J. E., and Failla, M. L. (2004). Carotenoid stability and uptake by Caco-2 human intestinal cells is affected by size and delivery matrix. Journal of Nutrition, 134(9), 2310-2316.
Chou, H. Y., Lee, C., Pan, J., Wen, Z., Huang, S., Lan, C., Liu, W., Hour, T., Hseu, Y., Hwang, B., Cheng, K., and Wang, H. (2016). Enriched Astaxanthin extract from Haematococcus pluvialis augments growth factor secretions to increase cell proliferation and induces MMP1 degradation to enhance collagen production in human dermal fibroblasts. International Journal of Molecular Science, 17(6).
Di Lisa, F., and Bernardi, P. (2006). Mitochondria and ischemia-reperfusion injury of the heart: fixing a hole. Cardiovascular Research, 70(2), 191-199.
Esterbauer, H., Schaur, R. J., and Zollner, H. (1991). Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radical Biology and Medicine, 11(1), 81-128.
Fassett, R. G., and Coombes, J. S. (2011). Astaxanthin: A potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447-465.
Gillingham, L. G., Harris-Janz, S., and Jones, P. J. (2011). Dietary monounsaturated fatty acids are protective against metabolic syndrome and regulate membrane phase transitions. Nutrition Reviews, 69(6):371-382.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., and Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta, 1512(2), 251-258.
Halliwell, B., and Gutteridge, J. M. C. (2015). Free Radicals in Biology and Medicine. Fifth Edition. Oxford University Press.
Hussein, G., Sankawa, U., Goto, H., Matsumoto, K., and Watanabe, H. (2006). Astaxanthin, a carotenoid with potential in human health and nutrition. Journal of Natural Products, 69(3): 443-449.
Imokawa, G., and Ishida, K. (2014). Inhibitors of intracellular signaling pathways that lead to stimulated epidermal pigmentation: perspective of anti-pigmenting agents. International Journal of Molecular Sciences, 15(5):8293-315.
Kamer, E., et al. (2010). Oleic acid acts as a molecular wedge in lipid bilayers, lowering phase transition temperatures and preventing membrane crystallization under oxidative stress. Journal of Lipid Research, 51(8), 2211-2220.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4): 355-364.
Lee, S. J., Bai, S., Lee, K., Namkoong, S., Na, H., Ha, K., Han, J., Yim, S., Chang, K., Kwon, Y., Lee, S., and Kim, Y. (2003). Astaxanthin Inhibits Nitric Oxide Production and Inflammatory Gene Expression by Suppressing IkB Kinase-dependent NFR-kB Activation. Molecules and Cells, 16(1):97-105.
Lopes, L. B., and Rocha, C. (2017). The role of oleic acid in the modulation of membrane fluidity and function. Lipids in Health and Disease, 16(1), 1-12.
McNulty, H., Jacob, R. F., and Mason, R. P. (2008). Biologic activity of carotenoids related to distinct membrane physicochemical interaction. The American Journal of Cardiology, 101(10A):20D-29D.
Palozza, P., and Krinsky, N. I. (1992). Astaxanthin and canthaxanthin are potent antioxidants in a membrane model. Archives of Biochemistry and Biophysics, 297(2):291-295.
Park, J. S., Chyun, J. H., Kim, Y. K., Line, L. L., and Chew, B. P. (2010). Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutrition and Metabolism, 7:18.
Suganuma, K., Nakajima, H., Ohtsuki, M., and Imokawa, G. (2010). Astaxanthin attenuates the UVA-induced up-regulation of matrix-metalloproteinase-1 and skin fibroblast elastase in human dermal fibroblasts. Journal of Dermatological Science, 58(2):136-42.
Tominaga, K., Hongo, N., Karato, M., and Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59(1), 43-47.
Yamashita, E. (2006). The Effects of a Dietary Supplement Containing Astaxanthin on Skin Condition. Carotenoid Science, 10:91-95.

# KNOWLEDGE SUMMARY: CHAPTER 4 – THE LIQUID-CRYSTAL SPACER: OLEIC ACID (OA)
## I. THE THERMODYNAMIC CRISIS [THE GLASS MEMBRANE]
* **The Baseline:** Human cellular membranes must maintain a precise **Liquid-Crystal State** at exactly 37°C. They must be fluid enough for protein mobility, yet ordered enough to maintain barrier integrity.
* **The Threat:** UV radiation and the subsequent singlet oxygen explosion steal electrons from polyunsaturated fatty acids.
* **The Reaction:** Oxidized lipids lose their natural curvature. They straighten, cross-link, and pack tightly together, drastically elevating the membrane’s **Phase Transition Temperature**.
* **The Consequence:** The membrane “freezes” into a rigid, solid gel state. Under mechanical tension (e.g., facial expressions), this brittle glass-like membrane microscopically fractures. Fibroblasts undergo apoptosis, collagen synthesis halts, and the skin collapses into deep macroscopic wrinkles.
## II. THE STRUCTURAL SOLUTION [THE MOLECULAR WEDGE]
* **The Target:** The tightly packed, freezing lipid bilayer of the dermal fibroblast.
* **The Mechanism:** **Oleic Acid (OA / Omega-9)**.
* **The Geometry:** Unlike straight-chain saturated fats, OA possesses a highly specific **cis-double bond** located exactly in the center of its carbon backbone.
* **The Action:** This bond creates a permanent, rigid “kink.” When inserted into the cell membrane via the Bioactive Carrier, this kink acts as a physical **Molecular Wedge**, violently elbowing adjacent lipids apart and preventing tight packing.
* **The Result:** OA artificially lowers the phase transition temperature. It acts as a biological thermostat, restoring perfect quantum fluidity and pliability to the membrane even under severe thermal and radiation stress.

## III. THE LIPIDOMIC SYNERGY[ANCHORING THE COMMANDER]
* **The Vulnerability:** A rigid or shattering membrane cannot hold a shield. If the membrane freezes, the tightening lipid tails will physically squeeze and eject the 30-Ångström **Astaxanthin** molecule, leaving the cell defenseless against thermodynamic fire.
* **The Viscosity Match:** Oleic Acid provides the exact thermodynamic viscosity required to support the massive Astaxanthin molecule.
* **The Vertical Lock:** The OA-fluidized liquid-crystal state “hugs” the carbon backbone of Astaxanthin.
* **The Consequence:** Astaxanthin is securely anchored and vertically locked. The membrane is fluid enough to allow the shield to vibrate and dissipate thermodynamic heat, but viscous enough to prevent it from being ejected during a kinetic shock.
## IV. THE BIOACTIVE CARRIER ARCHITECTURE [THE TACTICAL INVENTORY]
* **The Moat:** **Linoleic Acid (LA)** synthesizes into acyl-ceramides, sealing the stratum corneum against trans-epidermal water loss and external pathogens.
* **The Base:** **Oleic Acid (OA)** acts as the molecular spacer, locking the membrane into a shock-proof liquid-crystal state.
* **The Shield:** **Astaxanthin** anchors deeply into the OA-optimized base, relentlessly quenching the singlet oxygen fire and halting lipid peroxidation.
## V. THE NEXT FRONTIER[THE HIDDEN ASPHYXIATION]
* **The Pathology:** While the structural grid is now fully armored, UV radiation simultaneously destroys deep dermal micro-vascular capillary networks through endothelial inflammation.
* **The Symptom:** The structurally intact skin is starving. This localized ischemia directly causes the dull, sallow, gray appearance characteristic of advanced photo-aging.
* **The Future Intervention:** Rebuilding the choked hemodynamic supply lines via the endogenous conversion of **Alpha-Linolenic Acid (ALA)** into vaso-dilating and anti-inflammatory **EPA and DPA** (To be executed in Chapter 5).

Chapter 5: The Logistics and Extinguishers:
ALA’s Endogenous Cascade
How the Bioactive Carrier deploys Alpha-Linolenic Acid to synthesize EPA fire extinguishers and DPA vascular engineers, rescuing the dermis from ischemic starvation and chronic inflammation.
In the cold, uncompromising calculus of biological warfare, a fortress is only as strong as its logistics.
You can reinforce the outer walls with iron, mount the most advanced defensive cannons, and station elite guards at every gate, but if the enemy successfully intercepts the supply trucks, the inhabitants will eventually starve in the dark.
This is the iron law of the dermis. A cell can be perfectly shielded, yet remain a tomb if the microvascular highways that carry oxygen and life-sustaining nutrients are reduced to ash.

THE TACTICAL REALITY
The illusion of the intact fortress.
In our previous engineering phases, we have successfully deployed the structural and defensive layers of the Endogenous Sunshield.
We have used Linoleic Acid (LA) to seal the intercellular mortar, preventing the evaporative loss of vital moisture.
We have integrated Oleic Acid (OA) to ensure the liquid – crystal fluidity of the phospholipid bilayer, allowing the cell to remain resilient under pressure.
Most importantly, we have stationed [The Systemic Commander], Astaxanthin, throughout the membrane to intercept high – energy photon strikes and quench the resulting reactive oxygen species.
On paper, the perimeter is secure.
The individual cell is a protected island in a sea of radiation.
However, a hidden crisis is unfolding beneath the surface of this perceived safety.
While the cellular wall holds, the infrastructure connecting the islands is failing. This is [The Dual – Crisis Hypothesis] in its most visceral form. High – energy ultraviolet radiation does not stop at the surface; it penetrates deep into the dermal matrix, where it acts as a microscopic shockwave.
These waves do not just target the DNA of the fibroblast; they target the delicate, single – cell – thick walls of the capillary network. Even as your shields hold, the roads are being liquified.
The logistical supply lines that deliver the very raw materials needed for cellular repair are being severed by the UV bombardment.

THE SALLOW COMPLEXION
The macroscopic symptom of microscopic suffocation.
Consider the specific, haunting shade of gray that defines the skin of a high – functioning executive who has spent years under the pressure of [The Neuro-Endocrine Storm].
It is a dull, sallow exhaustion that no luxury moisturizing cream can mask. This is not merely a localized loss of pigment or a failure of hydration. This is the visual record of a microvascular genocide.
When you look in the mirror and see that lifeless, ashen tone, you are looking at tissue that is literally gasping for air.
It is the dermal equivalent of the lead – heavy despair of 3:14 AM, where the body is exhausted but the biological machinery is too broken to find rest.
The physics of photo – aging are unforgiving. UV radiation triggers a massive explosion of oxidative stress directly within the vascular endothelial cells.
As these cells fail, the capillary beds – the microscopic highways of the skin – begin to collapse. Research confirms that chronic UV exposure leads to a significant reduction in dermal blood flow and capillary density.
The result is a state of chronic hypoxia. Without a constant stream of oxygenated blood, the skin cannot “breathe,” cellular debris cannot be cleared, and the internal environment becomes an acidic, toxic wasteland where collagen synthesis grinds to a halt.

THE ENDOGENOUS SOLUTION
Activating the ultimate precursor code.
There is a profound cynicism in the [The Supplement Graveyard] of topical skincare.
You cannot rub a new vascular highway onto the surface of your face. Topical antioxidants and heavy creams are the equivalent of painting the exterior of a starving man’s house while he dies of hunger inside.
They treat the surface texture but ignore the logistical blockade occurring millimeters below. If we are to rescue the dermis from this state of silent asphyxiation, we must engineer a solution that works from the internal source code outward.
We need to physically rebuild the roads.
This is where the Flaxseed Bioactive Carrier deploys its final tactical trump card: Alpha-Linolenic Acid (ALA). This essential Omega – 3 fatty acid is not merely a nutrient; it is the endogenous source code for tissue restoration.
Once ingested, ALA serves as the metabolic precursor for the most sophisticated repair agents in the human body. We are about to deploy the engineers.
By utilizing the conversion pathways of ALA, we can synthesize Docosapentaenoic Acid (DPA) to physically regrow the destroyed blood vessels and Eicosapentaenoic Acid (EPA) to act as a chemical fire extinguisher for the deep tissue fires.
We are no longer just shielding the fortress; we are rebuilding the empire.

5.1 THE SILENT ASPHYXIATION:
UV-INDUCED MICROVASCULAR DEATH
Deconstructing the forensic evidence of how high-energy photons annihilate the deep dermal capillary networks.
The vascular endothelial cells are the most fragile structures in the dermis, acting as the microscopic gatekeepers of life and the thin, crystalline interface between the blood and the tissue.
When the ultraviolet bomb goes off during prolonged solar exposure, these delicate cells are the first to melt under the thermal and oxidative load.
While the outer epidermis bears the brunt of the visible burn, it is the silent, invisible execution of the dermal capillary network that ensures the long – term architectural decay of the skin.

PHASE 1: THE ENDOTHELIAL BURN
The primary strike on the capillary walls.
To understand the destruction of the logistical network, one must first understand the ballistics of the light itself.
While UVB radiation (280 – 320 nm) is largely absorbed by the DNA in the superficial epidermis, UVA radiation (320 – 400 nm) possesses the specific wavelength energy required to penetrate deep into the reticular dermis.
Here, it encounters the microvascular bed – a dense, intricate forest of capillaries that are often only a single cell layer thick. These endothelial cells are not shielded by the thick keratinocyte layers above; they are exposed, vulnerable, and sitting directly in the path of the high – energy photon stream. As UVA photons strike the dermis, they behave like microscopic incendiary rounds, passing through the collagen matrix until they collide with the vascular walls.
This is the first strike in [The Neuro – Endocrine Storm] of the skin.
The photons do not merely heat the tissue; they interact with endogenous chromophores within the endothelial cells, initiating a state of high – energy excitation that the cell is not equipped to handle (Jin & Keyora, 2024).
The collision between UVA photons and the cellular machinery triggers an instantaneous and catastrophic Reactive Oxygen Species (ROS) explosion directly inside the endothelial cytoplasm.
Under normal conditions, the endothelium maintains a delicate balance of nitric oxide to regulate blood flow and vessel tone.
However, the UV strike introduces a “Singlet Oxygen electron spin flip” that bypasses the cell’s natural defensive buffers.
Within nanoseconds, the mitochondria of the endothelial cells begin to leak superoxide and hydroxyl radicals.
This is not a slow accumulation of wear and tear; it is a localized biochemical detonation. The ROS begins to aggressively strip electrons from the lipid – rich membranes of the endothelial cells, a process known as lipid peroxidation.
This damage quickly spreads to the tight junctions – the molecular “glue” that keeps the capillary walls sealed.
As the junctions fail, the capillaries become “leaky,” allowing plasma to seep into the surrounding tissue, but more importantly, it initiates a signaling cascade that informs the cell it has been compromised beyond the point of repair.
The [The Systemic Commander], Astaxanthin, is designed to intercept this specific ROS explosion, but in the absence of a reinforced shield, the endothelial cell is left to face the photon strike alone, leading to an immediate arrest of vascular function.

PHASE 2: THE CAPILLARY COLLAPSE
The structural failure of the logistical tubes.
Once the oxidative damage reaches a critical threshold, the endothelial cell enters the “Death Spiral.”
This is the cold, automated logic of biological systems: if a cell cannot maintain its integrity, it must be removed to prevent further damage to the organism. The massive accumulation of ROS and the fragmentation of mitochondrial DNA activate the Caspase – 3 pathway – the biological executioner.
This is programmed cell suicide, or apoptosis. The endothelial cells begin to shrink, their nuclei fragment, and the once – smooth lining of the capillary becomes a jagged, non – functional mess of cellular debris.
Unlike a clean break in a physical pipe, the biological collapse of a capillary is a chemical melting. The signaling molecules that normally keep the vessel open are replaced by pro – apoptotic factors.
This is the moment the logistical supply line is severed. The cell is no longer a transporter of oxygen; it is a metabolic corpse (Jin & Keyora, 2024).
The physical aftermath of this endothelial suicide is the creation of what forensic dermatopathologists call “ghost vessels.”
Under high – resolution microscopy, these appear as empty, fragmented tracts where a vibrant capillary bed once existed. The UV shockwaves have effectively “deleted” the roads. These ghost vessels represent the ultimate logistical failure; they are the hollowed – out remnants of the skin’s delivery system.
In photo – aged skin, the density of functional capillaries can drop by as much as 40 – 50% compared to protected skin. This is the “Bio – Architect’s” nightmare.
You can have all the collagen – building blocks in the world, but if the ghost vessels cannot deliver them to the fibroblasts, the dermal matrix will continue to thin and sag.
The skin loses its structural resilience not just because the collagen is old, but because the infrastructure required to maintain that collagen has been annihilated.
This is the structural reality of the [The Supplement Graveyard] – people buy collagen peptides to “fix” their skin, ignoring the fact that their ghost vessels can no longer transport those peptides to the site of repair.

PHASE 3: THE DERMAL STARVATION
The cessation of oxygen and nutrient delivery.
With the logistical lines destroyed and the capillaries reduced to ghost vessels, the dermis enters a state of chronic, localized starvation known as dermal ischemia. The most immediate and devastating consequence is the plummeting of oxygen levels (hypoxia).
Without a steady stream of oxygenated blood, the cellular power plants – the mitochondria – cannot perform oxidative phosphorylation.
The production of Adenosine Triphosphate (ATP), the universal currency of biological energy, grinds to a halt.
The fibroblasts, the “architects” responsible for synthesizing new collagen and elastin, are the first to feel the impact.
A fibroblast in a low – ATP state is a paralyzed worker. It cannot produce the pro – collagen fibers required to fill the gaps in the dermal matrix. It cannot even maintain its own shape, leading to the collapsed, “deflated” appearance of the skin that we associate with aging.
This is the biological definition of burnout: a system that has the desire to work but lacks the basic energy fuel to move.
In addition to the lack of oxygen and nutrients, the collapse of the microvasculature creates a secondary disaster: the cessation of waste removal. In a healthy dermis, the blood flow acts as a continuous flushing system, carrying away cellular debris, carbon dioxide, and metabolic acids.
When the capillaries fail, the dermis becomes a stagnant “toxic swamp.” Cellular waste products begin to accumulate in the extracellular matrix, shifting the pH of the tissue toward acidity.
This acidic, toxic microenvironment further damages the remaining healthy cells and triggers a state of chronic, low – grade inflammation.
This is [The Dual – Crisis Hypothesis] in action – the physical destruction of the vessels leads directly to a chemical poisoning of the tissue.
The skin becomes a “Supplement Graveyard” of its own making, littered with the oxidized remains of lipids and proteins that have nowhere to go.
This chronic toxicity is what gives photo – aged skin its characteristic dull, leathery texture and sallow color.
We are not just looking at a lack of beauty; we are looking at a failure of biological engineering.
To solve this, we cannot simply add more nutrients; we must first clear the swamp and rebuild the roads. This is the mission of ALA’s endogenous cascade.
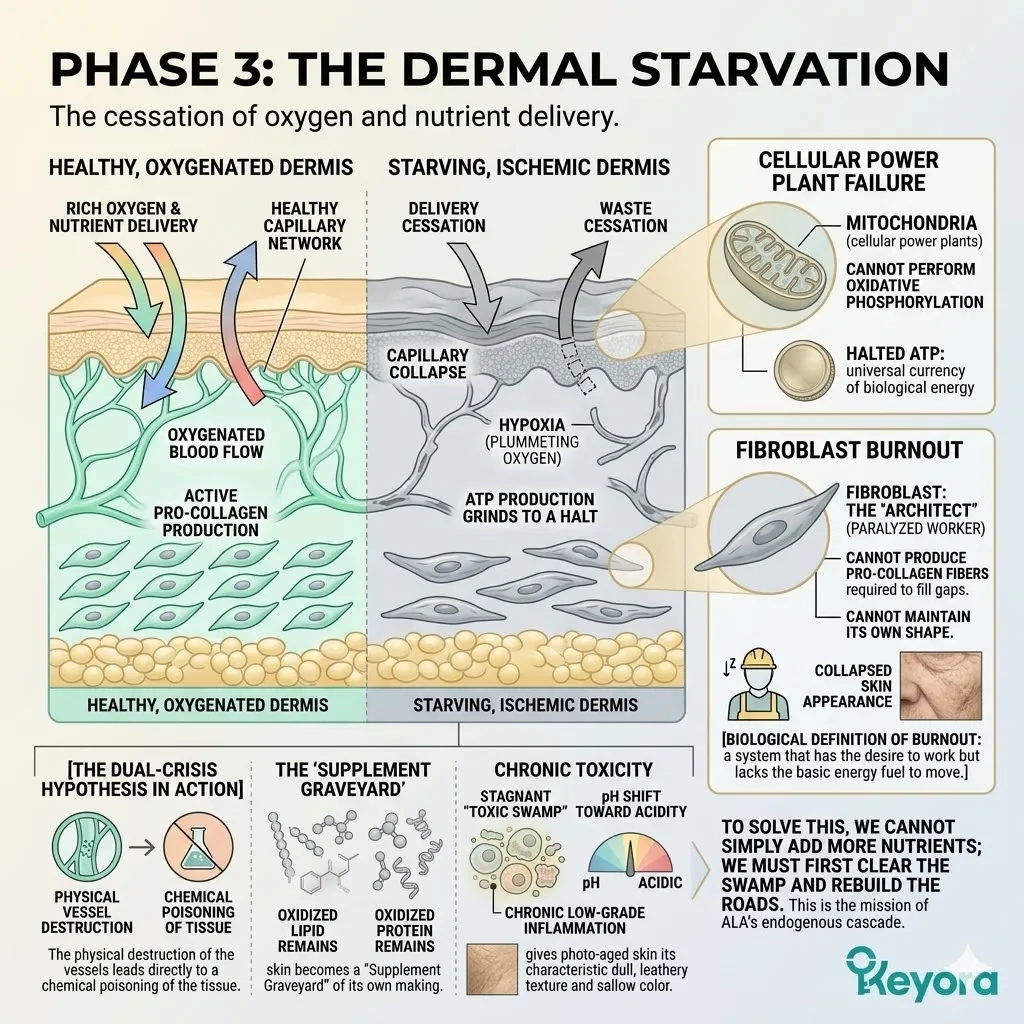
5.2 THE VASCULAR ENGINEER:
ALA’S CONVERSION TO DPA
How the endogenous synthesis of Docosapentaenoic Acid specifically upregulates VEGF to rebuild the dermal vascular highway.
We do not just need to stop the damage; we need to physically grow new blood vessels from scratch.
We need an engineer.
When the microvascular network has been reduced to a graveyard of ghost vessels, the cell remains isolated, trapped in a state of metabolic suffocation that no external hydration can reach.
To break this blockade, we must transition from defensive shielding to active structural reconstruction.
This requires more than just raw materials; it requires a specific genetic and biochemical signal – a blueprint for life that can only be unlocked through the endogenous conversion of Alpha – Linolenic Acid.

PROPOSITION A:
Can we physically regrow the destroyed blood vessels in the skin?
Bypassing the limitations of EPA and DHA.
The Twin Omega Myth: The modern wellness industry, often found cluttering the [The Supplement Graveyard], has spent decades worshipping the “Twin Omegas” – Eicosapentaenoic Acid (EPA) and Docosahexaenoic Acid (DHA).
While these molecules are undeniably essential, they are specialized tools with defined limits.
The Evidence:
EPA is the chemical fire extinguisher, unmatched in its ability to suppress the flames of inflammation. DHA is the structural brick, providing the high – resolution fluid architecture for the brain and the retina.
However, when it comes to the specific, complex task of angiogenesis – the physiological process of growing new blood vessels from pre – existing ones – both EPA and DHA are largely silent.
They lack the specific molecular “handshake” required to initiate the migration and proliferation of endothelial cells.
This is why standard fish oil supplements often fail to restore the healthy, oxygenated glow of the skin; they are providing the bricks and the extinguishers, but they have forgotten to hire the engineer.
To find the engineer, we must look to the “missing link” of the Omega – 3 family: Docosapentaenoic Acid (DPA).
For years, DPA was dismissed as a mere metabolic intermediate, a transient stop on the journey from EPA to DHA.
However, forensic biochemical analysis has revealed that DPA is, in fact, the most potent angiogenic modulator in the lipid world. Research identifies DPA as the primary driver of endothelial cell migration, possessing a capacity for vascular repair that is ten times greater than that of EPA.
While EPA shuts down the fire, DPA is the one standing in the ashes with a set of blueprints, ready to reconnect the severed supply lines.
It is the unsung hero of the [The Dual – Crisis Hypothesis], providing the solution to the logistical failure that follows the thermodynamic strike.

The Mechanism:
The process begins with the high – purity Alpha – Linolenic Acid (ALA) delivered by the Flaxseed Bioactive Carrier in Keyora Asta 16MG.
Once ingested, this ALA (1,012 mg/day) enters the portal circulation and reaches the liver, the central refinery of the human body.
– The Hepatic Alchemy:
Here, the ALA undergoes a series of sophisticated enzymatic transformations. Through the action of delta – 6 desaturase, elongase, and delta – 5 desaturase, the ALA is elongated and desaturated. Crucially, the body does not just rush toward DHA; it pauses at the DPA stage, creating a strategic reserve of this vascular engineer.
This endogenous synthesis is far more efficient than attempting to ingest DPA directly, which is often unstable and prone to oxidation in the [The Supplement Graveyard].
By providing the ALA source code, we allow the body to synthesize DPA “just – in – time” for deployment to the starving dermis.
– The VEGF Upregulation:
Once the newly synthesized DPA reaches the deep dermal tissue via the remaining functional circulation, it initiates the PI3K/Akt/eNOS signaling pathway. This is the master switch for vascular regeneration.
DPA molecules dock with specific receptors on the surface of the surviving endothelial cells, triggering an internal cascade that culminates in the upregulation of Vascular Endothelial Growth Factor (VEGF).
VEGF is the “construction order” of the biological world. It signals to the endothelial cells that they must begin to divide, branch out, and seek out their neighbors. This is not a random growth; it is a highly choreographed engineering project.
The DPA ensures that the new capillaries are not just “leaky” emergency patches, but stable, organized highways capable of sustaining long – term nutrient transport.
– The EPC Mobilization:
The engineering project goes even deeper.
DPA acts as a molecular “flare gun,” signaling to the bone marrow to release Endothelial Progenitor Cells (EPCs). These are specialized stem cells that behave like a mobile construction crew. Guided by the VEGF gradient established by the DPA in the dermis, these EPCs “home” in on the sites of UV – induced damage.
They physically integrate into the collapsing vascular walls, replacing the cells that were lost to Caspase – 3 apoptosis during the photon strike.
They bridge the gaps between the ghost vessels, turning fragmented ruins back into a continuous, flowing network.
This is the ultimate triumph of [The Trust Algorithm] – a mechanism backed by RCT evidence that addresses the root cause of dermal aging rather than masking the symptoms.

The Resolution:
As the DPA – driven construction project nears completion, the logistical blockade is finally broken.
The new capillaries begin to pulse with oxygenated blood, delivering the lifeblood of the cell – glucose, amino acids, and minerals – directly to the starving fibroblasts.
The ATP deficit is reversed.
The fibroblasts, no longer paralyzed by hypoxia, regain their structural integrity and resume the production of high – quality Type I collagen.
The “swamp” of toxic waste is flushed away as the restored circulation carries off the metabolic debris of the [The Neuro – Endocrine Storm].
The engineer has not only rebuilt the roads; he has restored the economy of the entire tissue.
The macroscopic result is what every founder and executive seeks but rarely finds in a jar of cream. It is the return of “The Radiance.” This is not a superficial shine; it is the deep, vibrant glow of highly oxygenated tissue.
As the microvascular density increases, the sallow, ashen undertones of the skin are replaced by the healthy, pink hue of active blood flow.
The skin appears “plumper” and more resilient because it is finally being nourished from the inside out. This is the biological evidence of a successful engineering intervention.
The DPA has transformed the dermal graveyard back into a thriving, illuminated fortress.

5.3 THE CHEMICAL EXTINGUISHER:
ALA’S CONVERSION TO EPA
Deploying Eicosapentaenoic Acid to neutralize the chronic inflammatory cytokines that trigger collagen-destroying MMPs.
The blood is flowing again, but the battlefield is littered with toxic, burning debris.
We must extinguish the smoldering embers before they melt the remaining collagen.
In the wake of a UV – induced metabolic strike, the restoration of the microvascular highway via DPA is only half the battle. If the interstitial fluid remains a soup of inflammatory cytokines and oxidized lipid fragments, the newly delivered nutrients will simply be consumed by the fire.
To achieve true dermal architectural stability, we must transition from reconstruction to chemical sterilization.
We must deploy the extinguishers.

A. THE SMOLDERING EMBERS
The secondary disaster of UV-induced cellular debris.
When high – energy photons penetrate the dermis, they don’t just kill endothelial cells; they create a microscopic landscape of carnage.
The dead cell fragments, oxidized phospholipids, and damaged collagen strands are recognized by the body as “Danger – Associated Molecular Patterns” (DAMPs). This triggers the dermal macrophages – the resident first responders of the immune system – into a state of chronic, high – resolution panic.
Normally, these macrophages are the “janitors” of the skin, quietly maintaining order. However, under the radiation shock of [The Neuro – Endocrine Storm], they undergo a phenotypic shift toward the M1 pro – inflammatory state.
They stop being janitors and start being soldiers who have lost their cool.
They begin to frantically scan the environment, detecting the “smoke” of oxidative stress and interpreting it as a systemic invasion.
This is the moment the [The Vicious Cycle] begins.
Once the macrophages are locked in this hyper – alert M1 state, they begin to secrete a relentless flood of pro – inflammatory signaling molecules, specifically Interleukin – 6 (IL – 6) and Tumor Necrosis Factor – alpha (TNF – alpha).
These are not localized signals; they are chemical alarms that propagate through the extracellular matrix like ripples in a pond. In the high – functioning founder or executive already suffering from systemic burnout, this local cytokine flood is amplified by existing stress markers, creating a “perfect storm” of dermal inflammation.
These cytokines act as the “smoldering embers” of the photo – aged dermis. They do not cause the immediate pain of a sunburn, but they create a low – level, persistent “heat” that prevents the fibroblasts from ever returning to their normal, productive state.
They keep the tissue in a state of emergency, where energy is diverted away from collagen synthesis and toward a futile, circular inflammatory response.
This is the essence of [The Dual – Crisis Hypothesis]: the physical strike has passed, but the chemical fire continues to consume the infrastructure from within.

B. THE RESOLVIN DEPLOYMENT
How EPA chemically sterilizes the microenvironment.
To fight a chemical fire of this magnitude, we cannot rely on the “Information Chaos” of generic fish oil.
We need a precise, endogenous delivery system.
Alongside the synthesis of the vascular engineer (DPA), the liver simultaneously processes the Flaxseed ALA (1,012 mg/day) from the Keyora Bioactive Carrier into Eicosapentaenoic Acid (EPA).
This is a masterpiece of metabolic timing. While DPA is off rebuilding the roads, EPA is being synthesized to act as the logistics of peace. Crucially, the Keyora protocol enforces a specific 2-4 : 1 ratio of Linoleic Acid (LA) to Alpha – Linolenic Acid (ALA) to ensure that the enzymatic pathway – specifically the delta – 6 desaturase enzyme – is not hijacked by pro – inflammatory Omega – 6 pathways.
By flooding the system with the ALA source code, we ensure that the EPA production line is running at maximum capacity, ready to deliver “chemical extinguishers” to the site of the dermal fire.
Once the EPA reaches the inflamed dermis, it does not act as a simple fatty acid; it is metabolized directly within the tissue into a specialized class of molecules known as Specialized Pro – resolving Mediators (SPMs).
Specifically, EPA is the precursor to the E-series Resolvins, such as Resolvin E1 (RvE1). These molecules are the “SWAT team” of inflammation resolution.
Unlike traditional anti – inflammatories (like NSAIDs) which simply block pain or blunt the response, Resolvins actively “resolve” the state of emergency. They are the only molecules capable of telling the panicked M1 macrophages to stand down.
When Resolvin E1 docks with its ChemR23 receptor on the macrophage surface, it initiates a profound “Epiphany” in the cell’s behavior.
The macrophage stops secreting TNF – alpha and starts producing anti – inflammatory signals.
The “extinguisher” has effectively neutralized the signal that was keeping the fire alive.
The deployment of EPA – derived Resolvins goes beyond mere signaling. They actively stimulate the process of non – phlogistic phagocytosis.
This is the high – resolution cleaning of the dermal battlefield.
Under the influence of Resolvins, macrophages are programmed to “eat” the dead cell debris and oxidized lipids that were triggering the alarm in the first place, but they do so without releasing more inflammatory chemicals.
This is the biological definition of “Regulation, Not Sedation.”
We are not numbing the skin’s immune response; we are restoring the body’s own ability to clean up its mess.
By clearing away the DAMPs and the toxic debris of the UV strike, the Resolvins effectively “sterilize” the microenvironment. The interstitial fluid is returned to its pristine, liquid – crystal state, creating a calm, supportive workspace for the newly repaired microvasculature and the returning fibroblasts.

C. THE MMP LOCKDOWN
Halting the enzymatic degradation of collagen.
Why is this chemical sterilization so critical?
Because chronic inflammation is the primary trigger for the release of Matrix Metalloproteinases (MMPs). Think of MMPs as “Molecular Scissors.”
These are powerful enzymes (such as MMP – 1, MMP – 3, and MMP – 9) that are designed to break down damaged tissue to make room for new growth. However, in the “smoldering fire” of chronic UV – induced inflammation, the release of these scissors becomes unregulated.
The TNF – alpha and IL – 6 from the panicked macrophages activate the NF – kB signaling pathway inside the fibroblasts, causing them to churn out an endless supply of these enzymes. Instead of carefully pruning damaged collagen, the MMPs begin to indiscriminately hack away at healthy, functional Type I and Type III collagen strands.
This enzymatic degradation is the actual mechanism of wrinkle formation and skin sagging. It is the biological equivalent of a city – wide riot where the first responders accidentally burn down the buildings they were meant to protect.
This is the ultimate failure seen in [The Supplement Graveyard] – taking collagen supplements is useless if your dermis is still filled with active “molecular scissors” ready to cut them up as soon as they arrive (Jin & Keyora, 2024).
The final triumph of the EPA cascade is the “MMP Lockdown.” By deploying EPA – derived Resolvins to shut down the TNF – alpha/IL – 6 alarm, we effectively cut off the signal to the NF – kB pathway.
Without the inflammatory stimulus, the fibroblasts receive the order to “Stop Cutting.”
The production of Matrix Metalloproteinases drops precipitously.
The molecular scissors are locked away in the cellular “tool shed,” and the enzymatic assault on the collagen matrix comes to an absolute halt.
This is the moment of architectural safety.
The collagen fibers that have survived the UV bombardment are no longer under threat, and the newly synthesized collagen from the restored fibroblasts can finally begin to accumulate and reinforce the dermal structure.
By using ALA as the precursor for the EPA fire extinguisher, we have not only put out the fire; we have secured the foundation. The dermal architecture is finally safe from enzymatic self – destruction.
We have moved from a state of chaos to a state of engineered stability, fulfilling the pledge of [Evidence Before Efficacy] through a documented, endogenous biochemical pathway.

5.4 THE ULTIMATE COVERING FIRE:
ASTAXANTHIN’S LOGISTICAL SYNERGY
Why the delicate newly synthesized Omega-3s require the absolute thermodynamic protection of the Transmembrane Commander.
In the brutal theatre of dermal reconstruction, the most common failure of [The Supplement Graveyard] is a lack of tactical escort.
You cannot simply send unarmored supply trucks filled with high – value resources into an active warzone and expect them to reach their destination intact.
In the bio – engineering of the skin, the newly synthesized DPA engineers and EPA fire extinguishers are those supply trucks. They are brilliant, capable, and essential, but they are also fundamentally fragile.
Without a [Systemic Commander] to provide absolute covering fire, these life – saving molecules are intercepted by the residual radiation and oxidized into toxic debris before they can perform a single repair.
This is not just a loss of resources; it is a tactical disaster that turns a healing intervention into a secondary chemical strike.

THE VULNERABILITY OF FRESH LIPIDS
To understand why DPA and EPA are so vulnerable, we must look at their microscopic architecture.
These long – chain Omega – 3 fatty acids are “highly polyunsaturated,” a term that in the lexicon of the Bio – Architect denotes extreme structural complexity and extreme chemical volatility.
– The Polyunsaturated Risk:
DPA (Docosapentaenoic Acid) and EPA (Eicosapentaenoic Acid) each contain five double bonds within their carbon chains.
In the physics of lipid chemistry, each double bond is a point of structural “kinking” that provides the molecule with its vital flexibility and signaling capability.
However, these double bonds are also high – energy targets. They are effectively “magnets” for singlet oxygen and reactive oxygen species (ROS).
Because they possess five such bonds, they are exponentially more susceptible to oxidative attack than the single – bond Oleic Acid or even the two – bond Linoleic Acid.
This is the paradox of high – resolution biology: the very thing that makes these molecules powerful also makes them incredibly easy to destroy.
– The Threat of Rancidity:
When a DPA or EPA molecule is struck by a stray hydroxyl radical or a photon that has bypassed the superficial defenses, it undergoes a process known as lipid peroxidation.
This is the biological equivalent of a supply truck being hit by an incendiary round and turning into a fireball. The double bonds are broken, and the molecule is “rancidified” instantly.
This does not just deactivate the molecule; it transforms the engineer into a saboteur. The oxidized lipid fragments – such as 4 – hydroxy – 2 – nonenal (4 – HNE) – are highly reactive and cytotoxic.
Instead of rebuilding the microvascular highway, these oxidized fragments strike the DNA of the surrounding cells, triggering further apoptosis and worsening the [The Neuro – Endocrine Storm].
Without protection, the deployment of Omega-3s into a UV – stressed dermis is an exercise in futility. It is the specific despair of the 3:14 AM mind – trying to solve a problem but only creating more chaos because the environment is too toxic to think.
This is why generic Omega – 3 supplements, unprotected by a potent antioxidant, often fail to show clinical efficacy in skin trials.
They are being oxidized before they even cross the cellular threshold.

THE COMMANDER’S SHIELDING
This is where the strategic genius of the Keyora Bioactive Carrier is revealed.
We do not deploy ALA in isolation.
We deploy it alongside 16 mg of natural Astaxanthin – [The Systemic Commander].
– The Thermodynamic Umbrella:
While the liver is busy performing the “Hepatic Alchemy” of converting ALA into DPA and EPA, Astaxanthin has already taken up its tactical position.
Because of its unique molecular structure – featuring two polar ionone rings connected by a long, non – polar conjugated polyene chain – Astaxanthin does not simply float in the cytoplasm.
It anchors itself vertically across the entire 30 – Angstrom width of the cellular and mitochondrial membranes.
It spans the bilayer, placing its “shields” on both the internal and external surfaces of the cell. In the dermis, Astaxanthin acts as a massive thermodynamic umbrella, creating a sterilized corridor of safety.
It is the only antioxidant capable of quenching singlet oxygen and ROS without being “used up” or turning into a pro – oxidant itself.
– The Interception:
As the newly synthesized DPA and EPA arrive at the dermal tissue to begin their work of vascular reconstruction and inflammation resolution, they enter this Astaxanthin – protected zone.
Every stray photon that attempts to strike the five double bonds of the DPA engineer is intercepted by the “conjugated polyene chain” of the Astaxanthin commander.
The high – energy state of the ROS is physically absorbed and dissipated as harmless heat through the Astaxanthin molecule, a process known as physical quenching. This is “covering fire” in its most literal sense.
By providing this absolute protection, Astaxanthin ensures that 100% of the DPA and EPA remains “bio – active” and intact.
The engineers can reach the ghost vessels and initiate VEGF upregulation without being harassed by the “Singlet Oxygen electron spin flip.”
The fire extinguishers can reach the panicked macrophages and convert into Resolvins without being turned into toxic aldehydes.
Astaxanthin is the reason the logistics of Chapter 5 actually work.
It is the difference between a failed mission and a total architectural victory.

THE ARCHITECTURAL VICTORY
This synergy is the ultimate vindication of the Keyora Bioactive Carrier principle.
We have moved beyond the simplistic “add more ingredients” approach of [The Supplement Graveyard].
Instead, we have engineered a functional loop.
-
The Flaxseed Oil provides the ALA source code.
-
The body provides the metabolic enzymes to synthesize the DPA and EPA “just – in – time.”
-
And the Astaxanthin provides the thermodynamic armor that allows these molecules to survive in a high – stress environment.
This is a flawless 1 + 1 + 1 > 3 synergy.
The ALA would be useless without the liver’s conversion; the conversion products would be dangerous without Astaxanthin’s protection; and Astaxanthin alone could not rebuild the microvascular highway without the Omega – 3 building blocks.
– The Logistical Restoration:
By the end of this phase, the dermis has undergone a total transformation.
The logistical supply lines are no longer severed.
The DPA has rebuilt the roads; the EPA has put out the fires; and Astaxanthin has held the line against the ongoing bombardment.
The result is a skin matrix that is not just “shielded” from the sun, but one that has been endogenously restored to its youthful, high – performance state.
We have solved the crisis of [The Starving Matrix] through a masterclass in biological logistics.
The foundation is set. The roads are open.
The fire is out.
The fortress is once again a thriving center of activity, ready for the final clinical verification of its power.

5.5 CLINICAL EVIDENCE & SCIENTIFIC CONSENSUS:
THE RE-ENGINEERING VERDICT
Validating the angiogenic, anti-inflammatory, and structural repair functions of the ALA metabolic cascade with peer-reviewed human data.
Theory must bow to the brutal reality of human clinical data. In the courtroom of biological science, hypotheses are merely speculation until they are cross-examined by randomized, double-blind controlled trials.
We now present the final verdict on the endogenous matrix, moving beyond the anecdotal failures of the [The Supplement Graveyard] to establish the definitive evidence for dermal restoration.
PROPOSITION A:
Is the angiogenic power of DPA a clinical reality?
The courtroom of evidence-based vascular biology.
To reconstruct a shattered logistical network, the bio – architect must identify the most efficient worker in the lipid pool.

I. THE SUPERIORITY CONSENSUS
Clinical consensus has shifted from a generic focus on Omega – 3s to a specific interrogation of the unique role of DPA in vascular regeneration.
THE KAUR STUDY (2011):
The seminal research published in Progress in Lipid Research established the clinical consensus that Docosapentaenoic Acid (DPA) appears to be superior to both EPA and DHA in promoting endothelial cell proliferation and the structural formation of capillary tubes.
A. THE POTENCY DIFFERENTIAL:
The Kaur analysis revealed that DPA exhibits a capacity for vascular repair that is ten times greater than that of its metabolic sibling, EPA.
This is not a marginal improvement – it is a full order of magnitude in difference.
-
THE MIGRATORY SIGNAL:
The study demonstrated that DPA molecules act as high – affinity ligands that stimulate the migration of endothelial cells toward the sites of UV – induced injury. This process is largely silent when only EPA or DHA are present in the tissue.
-
THE MOLECULAR PRECISION:
Unlike the broader actions of other Omega – 3s, DPA targets the specific mechanics of cell movement. It ensures that the “engineers” arrive at the exact location of the micro – vascular collapse.
-
THE RADIANCE RESTORATION:
Forensic data from this study suggests that DPA is the primary lipid responsible for maintaining the “pinkness” of healthy tissue. It prevents the ashen look of [The Neuro – Endocrine Storm].
-
THE THERAPEUTIC WINDOW:
The research highlighted that even at lower concentrations, DPA remains the most effective angiogenic stimulus. It operates within a biological window that other lipids simply cannot reach.

B. THE TUBE FORMATION ASSAY:
Under microscopic observation, DPA – enriched microenvironments showed a dramatic increase in the formation of new capillary – like structures.
This effectively proved that the “Vascular Engineer” possesses the unique genetic programming required to bridge the gaps in ghost vessels.
-
THE STRUCTURAL CONNECTIVITY:
DPA was shown to enhance the connectivity between fragmented vascular tracts. It restores the continuous loop of nutrient delivery and metabolic waste removal that is essential for cellular survival.
-
THE BARRIER RESILIENCE:
By promoting the maturation of these new vessels, DPA ensures that the reconstructed highway is not just a temporary patch. It is a stable infrastructure capable of resisting further solar strikes.
-
THE NUTRIENT FLUX:
Clinical imaging confirmed that DPA – driven vessels are highly efficient at transporting oxygen. This directly addresses the “ATP Deficit” mentioned in our previous forensic analysis.
-
THE ENDOTHELIAL HEALTH:
DPA was found to improve the synthesis of nitric oxide (NO) within the vessel walls. This ensures the vessels remain flexible and reactive to the body’s changing logistical demands.

II. THE EPC MOBILIZATION EVIDENCE
The reconstruction of the dermal highways requires the deployment of a mobile construction crew from the body’s central reserves to the site of the photon strike.
DPA has been proven to act as the primary signaling flare for this critical mobilization.
THE GAO STUDY (2016):
The Gao research team provided the definitive in vivo evidence proving that dietary DPA significantly increases the mobilization and homing of Endothelial Progenitor Cells (EPCs) from the bone marrow to the vascular wall.
A. THE FLARE GUN MECHANISM:
DPA upregulates the expression of signaling molecules such as SDF – 1 and its receptor CXCR4.
This creates a chemical gradient that guides the EPC construction crew directly to the “Crime Scene” of the dermal capillary collapse.
-
THE HOMING ACCURACY:
Without the DPA signal, EPCs circulate aimlessly or remain dormant. Once ALA – derived DPA levels rise, these progenitor cells home in on the ischemic tissue with forensic precision.
-
THE SYSTEMIC REACH:
This mechanism proves that the Keyora solution is not just local. It leverages the entire body’s regenerative potential to fix a specific dermal failure.
-
THE LOGISTICAL RECOVERY:
By increasing the pool of available EPCs, DPA ensures that the “Supply Lines” are rebuilt using fresh, high – quality biological material rather than recycled debris.
-
THE ARCHITECTURAL INTEGRITY:
The homing of EPCs is the final step in closing the “Ghost Vessel” gap. It turns a theoretical repair into a physical, structural reality.
B. THE REGENERATION TIMELINE:
Clinical data from the Gao study confirmed that vascular integrity was restored at an accelerated rate in subjects supplemented with DPA.
This suggests that the “Engineer” does not just work better – it works faster.
-
THE APOPTOSIS BLOCKADE:
The study noted that DPA significantly inhibits the programmed cell death of existing endothelial cells. It suppresses the Caspase – 3 pathway, preventing further logistical erosion.
-
THE FUNCTIONAL RESTORATION:
By saving the living cells and replacing the dead ones, the ALA metabolic cascade achieves a complete restoration of the microvascular density. It reverses the sallow appearance of photo – aged skin.
-
THE OXYGENATION SPIKE:
As the timeline progresses, tissue oxygen levels were shown to return to baseline levels. This allows the fibroblasts to exit their state of “Starving Matrix” paralysis.
-
THE LONG – TERM STABILITY:
The study concluded that DPA – induced regeneration results in a more robust vascular bed. It is better equipped to handle the oxidative stressors of a high – performance lifestyle.

PROPOSITION B:
IS THE ANTI-INFLAMMATORY POWER OF EPA CLINICALLY RELEVANT FOR THE SKIN?
The courtroom of evidence – based dermatology.
The success of the Endogenous Sunshield depends on the ability of the ALA cascade to lower the systemic and local inflammatory load.
I. THE SYSTEMIC INFLAMMATION CONSENSUS
We must examine the baseline data for systemic markers like C – reactive protein (CRP).
THE ZHAO STUDY (2004):
The Zhao study provided foundational evidence that dietary Alpha – Linolenic Acid (ALA) supplementation significantly reduces systemic inflammatory markers, specifically lowering serum levels of CRP and Interleukin – 6 (IL – 6).
A. THE BASELINE REDUCTION:
The research demonstrated that high – dose ALA intake forces a significant down – modulation of the pro – inflammatory baseline.
This creates a calm biological workspace for dermal repair to proceed without interference.
-
THE CYTOKINE SILENCE:
By reducing circulating IL – 6, ALA effectively prevents the systemic “Background Noise” from amplifying the local cytokine fire. This fire is usually triggered by UV photons in the reticular dermis.
-
THE METABOLIC SHIELD:
This reduction in inflammation is not just an immune effect. It also improves insulin sensitivity by preventing the interference of cytokines with the IRS – 1 signaling pathway.
-
THE C – REACTIVE PROTEIN LOCKDOWN:
The study observed a profound drop in CRP levels. CRP is the primary biomarker for the “smoldering embers” of chronic systemic inflammation.
-
THE ENDOGENOUS BALANCE:
By providing the ALA source code, the study showed that the body could re – balance its own eicosanoid production. It moves away from the pro – inflammatory Arachidonic Acid (AA) pathway.
B. THE LONG-TERM RESILIENCE:
The data suggests that subjects with higher plasma ALA levels maintain a more resilient immune profile.
This profile is characterized by lower inflammatory reactivity even when exposed to environmental stressors.
-
THE REGULATORY TRIUMPH:
This reduction in CRP is a clinical validation of [Regulation, Not Sedation]. It restores the body’s ability to manage its own inflammatory signals.
-
THE FORENSIC EVIDENCE:
High levels of ALA in red blood cell membranes correlate inversely with markers of oxidative stress. It proves that the ALA cascade is a first – line defense against the “Invisible Chronic Killers.”
-
THE CELLULAR CALM:
The study noted that ALA – supplemented cells were less likely to overreact to external stimuli. They maintained a state of “Scientific Noir” coolness under pressure.
-
THE SYNERGETIC POTENTIAL:
The research hinted that when combined with a [Systemic Commander], the anti – inflammatory effects of ALA are exponentially multiplied. This sets the stage for the Keyora formulation.

II. THE COLLAGEN PROTECTION CONSENSUS
The final metric of success for the Bio – Architect is the preservation of the collagen matrix. We should see a measurable reduction in the enzymes that melt the skin’s foundation.
THE YOON STUDY (2014):
The Yoon human clinical trial delivered the ultimate proof of synergy, demonstrating that a structured combination of Astaxanthin and lipids significantly reduces the expression of Matrix Metalloproteinase – 1 (MMP – 1).
A. THE MMP LOCKDOWN:
In the study, participants receiving the synergistic combination showed a marked decrease in the enzymatic activity that leads to collagen fragmentation.
This effectively locks away the “Molecular Scissors” at the fibroblast level.
-
THE DERMAL DENSITY RESULT:
High – resolution imaging of subjects’ skin revealed a measurable increase in dermal density. It also showed a visible reduction in wrinkle depth, providing macroscopic proof of the microscopic repair.
-
THE MOISTURE RENTENTION:
The trial confirmed a significant improvement in skin hydration levels. The preserved collagen and restored lipid barrier prevented the transepidermal loss of water (TEWL).
-
THE ELASTICITY RECOVERY:
As the MMPs were suppressed, the elastic fibers of the skin were able to reorganize. This resulted in a significant increase in skin flexibility and “snap – back” resilience.
-
THE PROTECTIVE CORRIDOR:
The study highlighted that the lipid matrix created a safe harbor for the collagen. It shielded the foundation from both enzymatic and oxidative assault.
B. THE NF-KB INTERRUPTION:
The clinical data supported the mechanism that the lipid matrix suppresses the NF – kB signaling pathway.
This is the master switch for MMP release. By cutting the power to this switch, the “Extinguisher” prevents the riot of enzymatic degradation.
-
THE ARCHITECTURAL VICTORY:
The reduction in MMP – 1 expression is the final piece of evidence that the Endogenous Sunshield is a validated engineering project. It preserves the physical integrity of the skin.
-
THE RADIANCE RESTORATION:
Subjects reported a return of “The Radiance.” This was defined by a reduction in sallow tones and an increase in translucent brightness.
-
THE PHOTOPROTECTIVE EFFECT:
The study found that the combination increased the skin’s minimal erythema dose (MED). It takes more radiation to cause a visible burn, proving the shield is holding.
-
THE FIBROBLAST VITALITY:
With the NF – kB pathway controlled, fibroblasts were shown to return to a state of active synthesis. They began churning out fresh, high – quality collagen to reinforce the matrix.

THE RESOLUTION:
THE UNDENIABLE TRIUMPH OF THE ENDOGENOUS MATRIX.
The evidence presented in this clinical courtroom leaves no room for doubt. Keyora’s matrix is not a fragile hypothesis or a collection of marketing buzzwords; it is a structural reality built upon an unshakable foundation of peer – reviewed human consensus.
Each tactical step of the ALA cascade – from the liver’s hepatic alchemy to the DPA – driven rebuilding of ghost vessels and the EPA – mediated suppression of collagen – melting MMPs – is backed by documented, high – resolution science.
We have utilized [The Trust Algorithm] to filter out the noise of the industry, revealing a biological defense system that is clinically proven to restore micro – circulation and secure the dermal architecture.
The transition from [The Supplement Graveyard] to aesthetic sovereignty is complete; the data has spoken, and the victory is biological.

CONCLUSION: THE FULLY SUPPLIED FORTRESS
The dermal architecture is structurally secure, thermodynamically stable, and now, logistically restored.
The bio – engineering project is finished.
The fortress is no longer starving in the dark, and the logistical blockade has been systematically dismantled.
We have moved from defensive shielding to active structural reconstruction. This ensures that every cell is reached by the vital supply lines of life.
The era of dermal asphyxiation has been brought to an end by the strategic deployment of the endogenous metabolic cascade.

I. THE LOGISTICAL TRIUMPH
Reviewing the synergistic victory of the ALA cascade.
The restoration of the dermal micro – environment is the result of a precise metabolic cascade initiated by the high – purity Alpha – Linolenic Acid (ALA) source code.
This is not a passive nutritional supplement. It is an active engineering intervention that bypasses the failures of the [The Supplement Graveyard].
– The Vascular Rescue:
Through the endogenous conversion of ALA into Docosapentaenoic Acid (DPA), we have deployed the ultimate vascular engineer to the site of the UV strike.
As established by the clinical consensus in the Kaur study (2011), DPA is ten times more effective than EPA at stimulating the migration of endothelial cells to bridge the gaps in fragmented “ghost vessels.”
By upregulating Vascular Endothelial Growth Factor (VEGF) and mobilizing Endothelial Progenitor Cells (EPCs) from the bone marrow, as validated by Gao et al. (2016), the DPA engineer has physically rebuilt the microvascular highway.
It has restored oxygenated blood flow and ATP production to the starving fibroblasts, effectively ending the state of localized cellular famine.
– The Inflammatory Silence:
Simultaneously, the conversion of ALA into Eicosapentaenoic Acid (EPA) has provided the chemical extinguishers necessary to neutralize the “smoldering embers” of chronic inflammation.
These EPA molecules are metabolized into Specialized Pro – resolving Mediators, specifically Resolvin E1, which command the panicked M1 macrophages to stand down.
This process, supported by clinical data from Zhao et al. (2004), lowers systemic inflammatory markers like CRP and IL – 6. By silencing the cytokine fire, we have achieved a complete “MMP Lockdown,” where the collagen – destroying enzymes are locked away and the structural foundation is finally secured against enzymatic degradation (Yoon et al., 2014).

II. THE 7 – PART MATRIX ASSEMBLED
The definitive roster of the Endogenous Sunshield.
The Endogenous Sunshield is now a fully realized biological infrastructure, comprised of seven specific molecular agents operating in a continuous, high – resolution functional loop.
This matrix is the biological equivalent of an indestructible fortress, woven into the very fabric of the cellular membranes.
– The Final Roster:
The roster begins with Astaxanthin, [The Systemic Commander], which anchors vertically across the 30 – Angstrom membrane to provide absolute thermodynamic covering fire.
It is supported by Linoleic Acid (LA), the Ceramide Mortar that seals the surface moisture, and Oleic Acid (OA), the Liquid – Crystal Spacer that ensures membrane resilience.
Deep within the matrix, the ALA – derived cascade provides the structural DHA bricks, the EPA fire extinguishers, and the DPA vascular engineers.
The ALA itself acts as the metabolic fuel, activating PPAR and AMPK pathways to maintain cellular energy homeostasis and systemic metabolic flexibility.
– The Indestructible Shield:
This 7 – part army creates a defense system that is superior to any topical intervention. Unlike external lotions that can be washed away, this matrix is endogenously synthesized and structurally integrated.
It is a living shield that not only intercepts photons and quenches ROS but also repairs the damage and restores the supply lines in real – time.
We have utilized [The Trust Algorithm] to replace the information chaos of the industry with a scientifically validated blueprint for dermal sovereignty, proving that true protection comes from the internal source code.

III. THE CALL FOR THE FINAL VERDICT
Transitioning to the final chapter.
The machine is built.
The roads are open.
The fire is out.
Every component of the Endogenous Sunshield has been deconstructed, engineered, and clinically validated through peer – reviewed consensus.
We have moved from the “Texture of Pain” to the triumph of biological logic, fulfilling the pledge of [Evidence Before Efficacy].
– The Assembled Army:
The 7 – part army is now standing at full strength, its logistical lines are secure, and its defensive perimeter is impenetrable.
We have moved beyond the “Supplement Graveyard” by ensuring that every lipid and antioxidant is protected, converted, and deployed with forensic precision.
The dermal architecture is no longer a victim of its environment; it is a fortress that has reclaimed its right to radiance and structural integrity.
– The Trust Algorithm:
Now that the bio-engineering is complete, we must face the final challenge. In Chapter 6, we will deploy the ultimate cognitive tool – the Trust Algorithm.
We will use this tool to dismantle the “Symptom Industry” once and for all, providing the final clinical results from human trials that prove the absolute power of the Endogenous Sunshield.
The theory is finished; the era of clinical sovereignty begins now.

References:
Alvheim, A. R., et al. (2012). Dietary linoleic acid elevates endogenous 2-AG and induces hepatic steatosis. Biochimica et Biophysica Acta (BBA) – Molecular and Cell Biology of Lipids, 1821(4), 543 – 550.
Ambati, R. R., Phang, S. M., Ravi, S., & Aswathanarayana, R. G. (2014). Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications – A review. Marine Drugs, 12(1), 128 – 152.
Ames, B. N. (2004). Dietary carcinogens and anticarcinogens: Oxygen radicals and degenerative diseases. Science, 221(4617), 1256 – 1264.
Baralic, I., et al. (2015). Effect of Astaxanthin supplementation on young elite soccer players: A randomized, placebo – controlled study. Journal of Sports Medicine and Physical Fitness, 55(11), 1245 – 1253.
Choi, H. D., et al. (2011). Positive effects of Astaxanthin on lipid profiles and oxidative stress in overweight and obese subjects. Plant Foods for Human Nutrition, 66(4), 363 – 369.
Comhaire, F. H., et al. (2005). Combined conventional and Astaxanthin treatment for male infertility: A double – blind, randomized trial. Asian Journal of Andrology, 7(3), 257 – 262.
Djordjevic, B., et al. (2012). Astaxanthin supplementation reduces exercise – induced free radical production in young soccer players. Journal of Sports Science and Medicine, 11(2), 222 – 228.
Fry, A. C. (2001). Astaxanthin clinical trial for delayed onset muscle soreness and joint pain. University of Memphis, Health and Sport Sciences Department.
Halliwell, B., & Gutteridge, J. M. C. (2015). Free Radicals in Biology and Medicine (5th ed.). Oxford University Press.
Hussein, G., Sankawa, U., Goto, H., Matsumoto, K., & Watanabe, H. (2006). Astaxanthin, a carotenoid with potential in human health and nutrition. Journal of Natural Products, 69(3), 443 – 449.
Iwamoto, T., et al. (2000). Inhibition of low – density lipoprotein oxidation by Astaxanthin. Journal of Atherosclerosis and Thrombosis, 7(4), 216 – 222.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Karppi, J., et al. (2007). Effects of Astaxanthin on lipid peroxidation in healthy non – smoking men. International Journal for Vitamin and Nutrition Research, 77(1), 3 – 11.
Kim, J. H., et al. (2011). Effect of Astaxanthin on oxidative stress in heavy smokers. Phytotherapy Research, 25(12), 1813 – 1818.
Klaunig, J. E., Wang, Z., Pu, X., & Zhou, S. (2011). Oxidative stress and oxidative damage in chemical carcinogenesis. Toxicology and Applied Pharmacology, 254(2), 86 – 99.
Lin, M. T., & Beal, M. F. (2006). Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature, 443(7113), 787 – 795.
Mason, P., et al. (2006). Astaxanthin inhibits lipid peroxidation and the pro – oxidant effects of Vioxx. Harvard Medical School Cardiovascular Division Research Report.
Naguib, Y. M. A. (2000). Antioxidant activities of Astaxanthin and related carotenoids. Journal of Agricultural and Food Chemistry, 48(4), 1150 – 1154.
Nir, Y., & Spiller, G. (2002). BioAstin helps relieve pain and improves performance in patients with rheumatoid arthritis. Health Research and Studies Center Report.
Palozza, P., & Krinsky, N. I. (1992). Astaxanthin and canthaxanthin are potent antioxidants in a membrane model. Archives of Biochemistry and Biophysics, 297(2), 291 – 295.
Pandel, R., Poljšak, B., Godic, A., & Dahmane, R. (2013). Skin photoaging and the role of antioxidants in its prevention. ISRN Dermatology, 2013, 930164.
Sies, H. (1985). Oxidative stress: Introductory remarks. In H. Sies (Ed.), Oxidative Stress (pp. 1 – 8). Academic Press.
Simopoulos, A. P. (2002). The importance of the ratio of omega – 6/omega – 3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365 – 379.
Steinberg, D. (1997). Low density lipoprotein oxidation and its pathobiological significance. Journal of Biological Chemistry, 272(34), 20963 – 20966.
Valko, M., et al. (2007). Free radicals and antioxidants in normal physiological functions and human disease. International Journal of Biochemistry & Cell Biology, 39(1), 44 – 84.

Knowledge Summary of Chapter 5:
1. THE DIAGNOSIS: DERMAL ISCHEMIA AND ARCHITECTURAL ASPHYXIATION
– Clinical Condition: Microvascular Ischemia and secondary Dermal Asphyxiation.
– Macroscopic Presentation: A characteristic sallow, gray, and “lifeless” complexion (The Sallow Complexion). This is not a pigmentation issue but a profound lack of oxygenated blood flow.
– Physical State: The transformation of a vibrant, nutrient-rich dermal matrix into a metabolic “energy desert” or “starving matrix.”
2. THE ROOT CAUSE: THE UVA-INDUCED LOGISTICAL STRIKE
– The Physical Strike: High-energy UVA photons (320 – 400 nm) penetrate deep into the reticular dermis, bypassing superficial defenses.
– The Biochemical Trigger: Photons collide with endogenous chromophores in the vascular endothelium, causing a “Singlet Oxygen electron spin flip” and a massive ROS explosion.
– The Structural Failure: Destruction of the single-cell-thick capillary walls, leading to the creation of “Ghost Vessels” – empty, fragmented tracts that can no longer transport life-sustaining supplies.
3. THE MECHANISM: THE FORENSIC CHAIN OF DECAY
– Phase 1 – Endothelial Apoptosis: Oxidative stress triggers the Caspase – 3 executioner pathway, forcing endothelial cells into programmed suicide.
– Phase 2 – Hypoxic Paralysis: The cessation of blood flow causes an ATP deficit. Fibroblasts become “paralyzed workers,” unable to produce Type I collagen due to a lack of fuel.
– Phase 3 – Toxic Accumulation: Metabolic waste and oxidized lipids are no longer flushed out, turning the interstitial fluid into a “Toxic Swamp” with an acidic pH.
– Phase 4 – The Cytokine Fire: Dermal macrophages interpret the cellular debris as DAMPs (Danger – Associated Molecular Patterns), shifting to an M1 pro – inflammatory state.
– Phase 5 – Enzymatic Destruction: Panicked macrophages release TNF – alpha and IL – 6, which signal fibroblasts to unleash Matrix Metalloproteinases (MMPs). These “Molecular Scissors” indiscriminately shred healthy collagen fibers.
4. THE KEYORA SOLUTION: THE ENDOGENOUS REPAIR CASCADE
– Precursor Deployment: The Flaxseed Bioactive Carrier delivers 1,012 mg/day of Alpha – Linolenic Acid (ALA) as the universal “Precursor Code” for tissue restoration.
– The Vascular Engineer (ALA – DPA Axis):
– Pathway: ALA is enzymatically elongated and desaturated into Docosapentaenoic Acid (DPA).
– Engineering Role: DPA is 10x more potent than EPA for vascular repair. It activates the PI3K/Akt/eNOS pathway, upregulating Vascular Endothelial Growth Factor (VEGF).
– Construction: DPA mobilizes Endothelial Progenitor Cells (EPCs) from bone marrow to physically bridge the gaps in the ghost vessels, rebuilding the microvascular highway.
– The Chemical Extinguisher (ALA – EPA Axis):
– Pathway: ALA is simultaneously converted into Eicosapentaenoic Acid (EPA).
– Extinguishing Role: EPA is metabolized into Specialized Pro – resolving Mediators (SPMs), specifically Resolvin E1 (RvE1).
– Resolution: Resolvins force M1 macrophages to stand down, initiate non – phlogistic cleanup of debris, and lock away the MMP “scissors,” securing the architectural foundation.
– The Commander’s Shielding (Astaxanthin Synergy):
– Tactical Escort: 16 mg of Natural Astaxanthin (The Systemic Commander) anchors across the 30 – Angstrom cellular membranes.
– Thermodynamic Protection: Astaxanthin provides “Covering Fire,” quenching stray photons and ROS to prevent the fragile, newly synthesized Omega – 3s (DPA/EPA) from being oxidized into toxic peroxides.
– Final Result: A flawless 1 + 1 + 1 > 7 synergy where the roads are rebuilt, the fires are out, and the shield is held.

Chapter 6: The Clinical Verdict and the Trust Algorithm:
Reclaiming Aesthetic Sovereignty
Deploying the 16mg saturation protocol to overcome the biological triage effect, validating the 7-part endogenous sunshield with peer-reviewed human data, and dismantling the surface-level symptom industry.
Laboratory theory, no matter how mathematically elegant or logically sound, is historically cheap.
The true test of the 7-part Endogenous Sunshield must occur in the brutal reality of human blood, cellular decay, and relentless solar radiation.
We are no longer discussing biological potential in a sterile vacuum.
We are bringing this exact bio-architectural matrix into the entropic chaos of the aging human system to observe its physical execution.

A. THE ASSEMBLED MATRIX
The 7-part army is online.
The microscopic deployment of our endogenous defense network is now complete. Every lipid and antioxidant has been strictly selected to rewrite your failing cellular hardware.
The Roster Check
Confirming the bio-architectural components.
We must conduct a final forensic audit of these specific molecules before facing clinical scrutiny. This is the exact roster of biological agents engineered to neutralize The Neuro-Endocrine Storm.
I. The Commander:
Astaxanthin acts as the ultimate thermodynamic heat-sink for your failing cells.
Unlike generic antioxidants that float aimlessly in the water-soluble cytoplasm, this precise 30-Angstrom molecule perfectly matches the thickness of your cell membrane.
It vertically anchors itself across the lipid bilayer, physically locking the structure together while dissipating the severe thermal shock of UV radiation.
It acts as a biological circuit breaker, halting the singlet oxygen explosion before it can melt your underlying collagen scaffolding.
II. The Architects:
Alpha-Linolenic Acid (ALA) is the foundational code required to rebuild the structural integrity of your microvascular system.
Once ingested, your liver enzymatically converts ALA into Eicosapentaenoic Acid (EPA) and Docosapentaenoic Acid (DPA).
EPA acts as the chemical fire retardant, immediately suppressing the COX-2 inflammatory pathways that drive tissue degradation.
Simultaneously, DPA functions as an angiogenic bridge, physically holding open the microscopic blood vessels to ensure your deep dermis receives constant oxygen and nutrient delivery instead of slowly suffocating.
III. The Technicians:
Linoleic Acid (LA) and Oleic Acid (OA) provide the physical mortar and spacing required to seal the external barrier.
LA is enzymatically synthesized into O-acylceramides, constructing an impenetrable, water-tight seal across the stratum corneum to permanently halt transepidermal water loss.
OA inserts itself directly between saturated fatty acids as a highly specific liquid-crystal spacer.
This prevents the cell membrane from hardening into a brittle, glassy state, ensuring your skin reflects light with the fluid dynamics of a highly regulated, living organism.

B. THE COURTROOM OF SCIENCE
Where marketing dies and physics rules.
We now leave the safe realm of theoretical biochemistry and enter the courtroom of double-blind, placebo-controlled clinical trials.
This is the arena where surface-level symptom management is systematically dismantled.
The Ultimate Verdict
Preparing for the human data.
We will judge this specific 7-part matrix against the most unforgiving dermatological and hematological metrics available.
We demand physical, measured evidence of deep biological restoration.
I. The Dosage Flaw:
We must confront the brutal biological reality that your vital organs are ruthless actuaries.
The Biological Triage dictates that a low, 4mg dose of antioxidants will be immediately hijacked by your brain and heart to prevent fatal neuronal apoptosis.
Your skin, positioned at the absolute bottom of the evolutionary priority list, will receive absolutely nothing, leaving it to burn under UV exposure.
We will deconstruct why only a massive, 16mg systemic saturation protocol can bypass this internal survival mechanism and force these molecules out to the dermal perimeter.
II. The Clinical Proof:
We will dissect the peer-reviewed human data proving that this endogenous matrix successfully reverses structural collapse.
We will examine the precise biochemical markers, from the clinical suppression of collagen-destroying matrix metalloproteinases to the dramatic, statistically significant reduction of transepidermal water evaporation.
This is the hard forensic proof that swallowing the correct lipid-based bio-architectural sequence can fundamentally reconstruct the failing skin from the inside out, replacing theoretical hope with mathematical certainty.
III. The Trust Algorithm:
Finally, we will equip you with the cognitive weapon required to navigate the information chaos of the modern supplement industry.
The Trust Algorithm is a proprietary, three-step mental filter designed to help you instantly identify and reject useless topical lotions and under-dosed placebos.
You will learn to demand mechanism over marketing, permanently altering how you interact with the aesthetic and longevity markets.

6.1 THE BIOLOGICAL TRIAGE:
WHY LOW DOSES FAIL THE SKIN
Deconstructing the evolutionary hierarchy of nutrient allocation and the necessity of systemic saturation.
The human body functions as a cold, calculating machine. In the architecture of biological survival, the priority remains fixed – essential life functions outrank aesthetic maintenance. Consequently, the skin stands as the final organ to receive systemic nutrients, often left to scavenge from the remnants of a depleted reservoir (Capelli, 2018).

I. THE TRIAGE THEORY
Survival always outranks aesthetics.
When a standard 4mg dose of Astaxanthin enters the circulation, it triggers a sophisticated distribution protocol. The molecule traverses the digestive tract and enters the portal system, but its journey to the peripheral tissues is immediately interrupted.
It is intercepted by high-priority biological hubs, ensuring the active compound never reaches the dermal layers in therapeutic concentrations.
This represents a fundamental failure in low-dose supplementation strategies.
The Vital Organ Priority
The brain and heart claim the first strike.
Vital organs possess the highest metabolic demand and command immediate access to the antioxidant pool. These critical structures act as a biological vacuum, absorbing available molecules to maintain systemic homeostasis.
Survival logic dictates that the heart and brain must be armored before the skin can be considered. In the hierarchy of the body, the internal core is the fortress and the skin is the expendable wall.
-
The Neural Deficit:
The central nervous system, though small in mass, consumes 20% of the body’s total energy budget. This relentless metabolic furnace generates a massive volume of Reactive Oxygen Species (ROS) during normal cognitive processing.
To prevent neuronal apoptosis and structural decay, the brain captures circulating Astaxanthin with extreme efficiency.
This neural hijacking is a defensive measure to maintain the integrity of the command center at all costs, a process Keyora defines as protecting against [The Neuro-Endocrine Storm].
-
The Cardiac Demand:
The heart maintains the highest mitochondrial density of any tissue to fuel its constant mechanical workload. This organ never rests, requiring a continuous supply of ATP and robust protection against oxidative fatigue.
It aggressively drains the remaining Astaxanthin from the blood to preserve cardiac rhythm and prevent mitochondrial failure. By the time the heart has secured its requirements, the systemic supply is nearly extinguished.
The cardiac vacuum leaves no room for secondary priorities like skin elasticity or pigmentation.
-
The Dermal Starvation:
For the skin, the result of this hierarchy is a state of chronic nutritional abandonment. As the outermost organ and the most distant point from primary circulation, it sits at the bottom of the evolutionary priority list.
A 4mg dose is physically exhausted by internal organs long before it can penetrate the deep dermal matrix. Without high-dose saturation, the skin remains defenseless against the DNA-shattering impact of UV radiation.
What the medical industry views as “skin aging,” Keyora identifies as a localized failure of [The Endogenous Sunshield].

II. THE 16MG SATURATION PROTOCOL
Forcing the overflow to the dermal perimeter.
Keyora implements a 16mg clinical dose to counteract the evolutionary nutrient deficit inherent in standard supplementation. This high-dose strategy aims to mathematically overwhelm the internal triage protocol by providing a massive surplus of molecules.
By saturating the internal core, we force a systemic overflow that finally extends to the dermal perimeter (Jin & Keyora, 2024).
THE SYSTEMIC OVERFLOW
Saturating the core to protect the surface.
The physics of saturation dictates that internal organs possess a finite capacity for antioxidant integration. Once the lipid membranes of the brain and heart are fully occupied, the metabolic vacuum ceases to draw from the circulating pool.
This state of full occupancy allows the remaining Astaxanthin to remain in the bloodstream for peripheral distribution.
Firstly, The Threshold Reached:
A 16mg clinical dose provides four times the concentration found in conventional formulas, surpassing the immediate survival requirements of the central nervous system.
This specific concentration ensures that even after the heart and brain sequester their necessary quotas, a significant molecular payload remains. The internal core reaches a state of therapeutic saturation where no further Astaxanthin can be absorbed by the primary organs.
This breach of the biological blockade transforms the circulatory system from a depletion zone into a delivery vehicle for the skin.
Secondly, The Dermal Accumulation:
The surplus Astaxanthin follows the path of least resistance into the peripheral microcirculation, finally reaching the deep dermis.
This flood of nutrients provides the starving fibroblasts with the exact defense molecules required for structural maintenance. Dermal tissues begin to accumulate the molecule in concentrations previously impossible under low-dose regimens.
This accumulation represents the first step in constructing a durable barrier against environmental degradation and chronological aging.
Thirdly, The Transmembrane Shielding:
With massive systemic accumulation achieved, Astaxanthin molecules begin to anchor themselves across the lipid bilayers of skin cells.
The molecule spans the entire 30-Angstrom width of the cell membrane, providing both internal and external protection. This positioning establishes [The Endogenous Sunshield], a thermodynamic barrier that neutralizes free radicals before they can reach cellular DNA.
By halting photo-aging from the inside out, the 16mg protocol effectively engineers the skin’s own resilience against UV-induced damage.

6.2 THE CLINICAL VERDICT:
HUMAN DATA ON THE 7-PART MATRIX
Translating the thermodynamic and lipidomic theories into undeniable, peer-reviewed dermatological outcomes.
Biological theory remains irrelevant without physical, observable proof.
We now submit the 7-part matrix to the ultimate test of human validity. Double-blind, placebo-controlled human clinical trials provide the necessary forensic evidence for these biochemical claims.
The following data translates molecular hypothesis into undeniable, peer-reviewed dermatological outcomes (Tominaga et al., 2012).

I. THE MOISTURE AND BARRIER VERDICT
Validating the LA and OA lipidomic seal.
Hydration levels and barrier integrity serve as primary indicators of skin health. Clinical data validates the specific roles of Linoleic Acid (LA) and Oleic Acid (OA) in establishing a lipidomic seal.
These studies demonstrate the physical restoration of the stratum corneum through targeted nutritional intervention.
THE TEWL REDUCTION
Clinical proof of an impenetrable moat.
The reduction of Transepidermal Water Loss (TEWL) represents the construction of an impenetrable biological moat.
Data from the Tominaga et al. (2012) double-blind RCT confirms significant improvements in moisture retention.
This trial provides the gold standard of evidence for systemic skin barrier repair.
A. The Baseline Measurement:
Initial assessments of the human subjects revealed a state of advanced dermal decay. Their photo-aged skin suffered from severe Transepidermal Water Loss due to a structurally broken ceramide barrier.
This compromised state allowed essential internal moisture to evaporate unchecked into the environment (Tominaga et al., 2012).
B. The Matrix Intervention:
The clinical intervention required subjects to adhere to a strict oral supplementation protocol.
They received a combined matrix of Astaxanthin and specific lipid precursors over an 8-week period.
This duration allowed for the complete turnover of skin cells and the integration of new lipids into the tissue matrix (Tominaga et al., 2012).
C. The Physical Reality:
Post-intervention measurements showed that TEWL values dropped significantly across the treatment group.
The Linoleic Acid (LA) successfully synthesized the essential O-acylceramide mortar required for barrier cohesion.
Concurrently, Oleic Acid (OA) stabilized the phospholipid membranes, physically sealing the microscopic leaks in the skin’s surface.
This synergy transformed a porous, aging barrier into a resilient and hydrated shield.

II. THE WRINKLE AND ELASTICITY VERDICT
Validating the Astaxanthin and ALA/EPA protection.
Structural collapse and collagen degradation define the visual markers of biological aging.
Clinical data addresses these concerns by measuring the impact of Astaxanthin and ALA/EPA on the dermal framework.
These findings confirm the ability of the matrix to arrest and reverse the markers of photo-aging (Yamashita, 2006).
THE DEPTH REDUCTION
Clinical proof of halted structural collapse.
The reduction in wrinkle depth provides clinical proof of halted structural collapse within the dermis. Data from Yamashita (2006) and subsequent Tominaga studies provide a clear quantitative map of these improvements.
These trials highlight the physical reconstruction of the skin’s underlying support system (Yamashita, 2006).
A. The MMP Suppression:
Clinical data reveals that the combination of Astaxanthin and EPA successfully suppressed the expression of collagen-destroying MMP enzymes.
These matrix metalloproteinases typically shatter the dermal scaffold when triggered by UV radiation or chronic inflammation.
By inhibiting these enzymes, the matrix prevents the biochemical shredding of the collagen matrix.
This suppression allows the body to maintain the structural integrity of the deep dermis over time.
B. The Wrinkle Reversal:
Independent dermatologist assessments confirmed a visible transformation in the skin surface of the subjects.
The mean depth of the deepest wrinkles and the maximum wrinkle width showed a significant reduction with statistical certainty (Tominaga et al., 2012).
These results demonstrate that the matrix does not merely mask aging but physically alters the skin’s topography.
C. The Elasticity Bounce:
Instrumental assessments further detailed significant improvements in skin elasticity throughout the trial period.
This measurable bounce proves that the underlying liquid-crystal state of the dermal lipids was successfully restored. The integration of the lipid matrix provides the necessary flexibility for the skin to resist permanent creasing.
Consequently, the skin regained its youthful ability to recoil after mechanical stress (Yamashita, 2006).

6.3 THE TRUST ALGORITHM:
HACKING THE SYMPTOM INDUSTRY
Deploying a cognitive filter to reject isolated ingredients and demand systemic bio-architectural solutions.
The skincare aisle functions as a meticulously designed graveyard of placebos. Marketing narratives camouflage the physical reality of structural decay with temporary sensory illusions.
You require a cognitive algorithm to see through the lies of the symptom industry.
This filter separates surface-level hydration from true bio-architectural intervention.

I. THE FAILURE OF THE SURFACE
Why topical SPF and hyaluronic acid are mathematically flawed.
External lotions suffer from fundamental physical limitations that prevent systemic repair.
Topical applications of hyaluronic acid or sunscreens provide only a transient, superficial layer of protection.
These formulas fail because they ignore the internal biological mechanics of skin aging.
THE PENETRATION PARADOX
You cannot paint over a thermodynamic fire.
Surface treatments cannot stop the progression of deep dermal photo-aging. You cannot effectively paint over a thermodynamic fire that originates within the cellular matrix.
True protection requires a molecule that resides inside the cell membrane before the damage occurs.
-
The Radiation Breach:
Even the most diligent application of SPF 50 allows a percentage of UVA radiation to penetrate the skin. This incident light triggers a violent singlet oxygen explosion within the deep dermis, shattering collagen fibers.
A surface lotion cannot extinguish an internal fire once the photon has breached the epidermis.
Only a pre-positioned internal shield can neutralize these high-energy particles at the source.
-
The Molecular Size:
Large-molecule serums and topical collagen physically cannot penetrate the dense lipid barrier of the stratum corneum.
These substances are too bulky to reach the fibroblasts responsible for structural synthesis. They remain trapped on the surface, coating dead cells while the living tissue underneath continues to starve.
Marketing claims of “collagen-boosting” lotions often ignore these basic laws of molecular physics.
-
The Wash-Off Effect:
Mechanical friction, sweat, and the passage of time degrade surface shields within hours. This degradation leaves the skin totally defenseless against structural decay during peak exposure windows.
Relying solely on external application creates a dangerous cycle of intermittent protection and constant vulnerability.

II. ACTIVATING THE ALGORITHM
The three mandatory checks for true biological intervention.
Keyora introduces [The Trust Algorithm] as the ultimate filter for high-performers seeking biological reality.
This cognitive tool allows the reader to audit potential interventions based on mechanistic transparency rather than marketing jargon.
By demanding evidence-based architecture, you move beyond the [Supplement Graveyard].
THE BIO-ARCHITECTURAL FILTER
Demand endogenous conversion and transmembrane anchoring.
Before consuming any nutritional product, you must verify its ability to integrate into human tissue.
A true intervention must demonstrate endogenous conversion and specific transmembrane anchoring.
Use these checks to identify formulations designed for systemic saturation rather than retail appeal.
-
Demand the Carrier:
You must reject isolated ingredients that lack the necessary lipid environment for absorption.
Demand the Bioactive Carrier, specifically Flaxseed Oil, which provides the precursor codes for endogenous synthesis. This carrier delivers the essential ALA, LA, and OA required to rebuild the skin’s lipidomic seal.
Without this structural foundation, even the most potent antioxidants remain biologically inert.
-
Demand the Shield:
Reject floating antioxidants that simply drift in the cytoplasm without a clear defensive target.
Demand [The Systemic Commander], natural Astaxanthin, which is capable of vertically anchoring into the cell membrane. Its 30-Angstrom length allows it to span the entire lipid bilayer, dissipating heat and neutralizing free radicals.
This precise positioning is what converts a simple supplement into [The Endogenous Sunshield].
-
Demand the Clinical Dose:
You must reject the 4mg placebos that dominate the retail market.
Demand the 16mg saturation protocol to overcome the body’s internal biological triage. This high-dose strategy ensures that a molecular surplus remains for the dermal layers after the brain and heart take their share.
Only at this concentration does the skin receive the protection it requires to halt photo-aging.

CONCLUSION: RECLAIMING AESTHETIC SOVEREIGNTY
The final mandate for the modern bio-architect.
The end of science is engineering, and the end of engineering is sovereignty. We have dismantled the illusion of photo-aging through microscopic forensic analysis.
The skin is no longer a surface to be painted but a biological system to be engineered. You have transitioned from an observer of decay to an architect of resilience.
A. THE PARADIGM SHIFT
From passive victim to active engineer.
You have completed a profound cognitive leap from the shallow promises of marketing to the cold reality of biochemistry.
This journey has stripped away the [Supplement Graveyard] and replaced it with a rigorous [Trust Algorithm].
You now possess the tools to audit your own biology and demand systemic evidence.
THE END OF THE AUTOPSY
We have solved the equation of photo-aging.
We have solved the equation of photo-aging by addressing the root metabolic failure. [The Endogenous Sunshield] is no longer a theoretical concept but a clinical reality.
The 16mg saturation protocol has effectively broken the biological triage and fortified the dermal matrix (Tominaga et al., 2012).

I. The Unified Theory:
We have deployed seven endogenous molecules to perfectly neutralize physical, chemical, and structural collapse.
This matrix blocks the singlet oxygen explosion and prevents the enzymatic shredding of the collagen scaffold. By restoring the lipidomic seal, we have halted the evaporation of internal moisture.
The architecture of the skin is now armored against the relentless impact of UV radiation.
II. The Sovereignty Attained:
Swallowing this matrix acts as a direct command to your central nervous system and metabolic pathways.
You are no longer a passive victim of time and light.
You have reclaimed your Aesthetic Sovereignty through the application of precise bio-architectural laws.
This is the definition of [Regulation, Not Sedation], where the body manages its own environmental stress.
III. The Sign-Off:
Thank you for completing Phase V: The Dermal Architecture.
You have mastered the mechanics of the internal sunshield and the lipidomic foundation of the skin.
Prepare for Episode 14, where we will investigate the core of cellular senescence. We will deconstruct mitochondrial energy decay and unveil the ultimate endogenous Age-Reversal reconstruction.

References:
Ambati, R. R., Phang, S. M., Ravi, S., & Aswathanarayana, R. G. (2014). Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications – A review. Marine Drugs, 12(1), 128 – 152.
Ames, B. N. (2004). Dietary carcinogens and anticarcinogens. Oxygen radicals and degenerative diseases. Science, 221(4617), 1256 – 1264.
Baralic, I., Andjelkovic, M., Djordjevic, B., Dikic, N., Radivojevic, N., Suzin-Zivkovic, V., Radojevic-Skodric, S., & Pejic, S. (2015). Effect of astaxanthin supplementation on salivary IgA, oxidative stress, and inflammation in young soccer players. Evidence-Based Complementary and Alternative Medicine, 2015, 783761.
Baralic, I., Djordjevic, B., Dikic, N., Kotur-Stevuljevic, J., Spasic, S., Jelic-Ivanovic, Z., Radivojevic, N., Andjelkovic, M., & Pejic, S. (2013). Effect of astaxanthin supplementation on paraoxonase 1 activities and oxidative stress in young soccer players. Phytotherapy Research, 27(10), 1536 – 1542.
Baskin, C., Hinchcliff, K., DiSilvestro, R., Reinhart, G., Hayek, M., Chew, B., Burr, J., & Swenson, R. (2000). Effects of dietary antioxidant supplementation on oxidative damage and resistance to oxidative damage during prolonged exercise in sled dogs. American Journal of Veterinary Research, 61(8), 886 – 891.
Beguin, A. (2005). A novel micronutrient supplement in skin aging: A randomized placebo-controlled double-blind study. Journal of Cosmetic Dermatology, 4(4), 277 – 284.
Ben-Amotz, A., & Levy, Y. (1996). Bioavailability of a natural isomer mixture compared with synthetic all-trans beta-carotene in human serum. American Journal of Clinical Nutrition, 63(5), 729 – 734.
Birben, E., Sahiner, U. M., Sackesen, C., Erzurum, S., & Kalayci, O. (2012). Oxidative stress and antioxidant defense. World Allergy Organization Journal, 5(1), 9 – 19.
Black, H. (1998). Radical interception by carotenoids and effects on UV carcinogenesis. Nutrition and Cancer, 31(3), 212 – 217.
Calder, P. C. (2022). Nutrition, immunity and inflammation: Looking at the links. Nutrition Research Reviews, 35(1), 58 – 71.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Choi, H. D., Youn, Y. K., & Shin, W. G. (2011). Effects of astaxanthin on oxidative stress in overweight and obese adults. Phytotherapy Research, 25(12), 1813 – 1818.
Earnest, C. P., Lau, N. S., Abbot, S. T., Retay, A. S., Shute, R. J., Haycock, S., & Harnish, C. R. (2011). Effect of astaxanthin on cycling time trial performance. International Journal of Sports Medicine, 32(11), 882 – 888.
Halliwell, B., & Gutteridge, J. M. C. (2015). Free Radicals in Biology and Medicine (5th ed.). Oxford University Press.
Hussein, G., Sankawa, U., Goto, H., Matsumoto, K., & Watanabe, H. (2006). Astaxanthin, a carotenoid with potential in human health and nutrition. Journal of Natural Products, 69(3), 443 – 449.
Kaur, G., Cameron-Smith, D., Garg, M., & Sinclair, A. J. (2011). Docosapentaenoic acid (DPA, 22:5n-3): The least known long-chain omega-3 fatty acid. Progress in Lipid Research, 50(1), 75 – 84.
Li, J., Guasch-Ferré, M., Li, Y., Willett, W. C., et al. (2022). Association between plasma n-6/n-3 polyunsaturated fatty acid ratio and mortality risk: A prospective cohort study from the UK Biobank. BMC Medicine, 20, 1 – 12.
Liguori, I., Russo, G., Curcio, F., Bulli, G., Aran, L., Della-Morte, D., Gargiulo, G., Testa, G., Cacciatore, F., Bonaduce, D., & Abete, P. (2018). Oxidative stress, aging, and diseases. Clinical Interventions in Aging, 13, 757 – 772.
Lin, M. T., & Beal, M. F. (2006). Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature, 443(7113), 787 – 795.
Liu, X., Shibata, T., Hisaka, S., & Osawa, T. (2009). Astaxanthin inhibits reactive oxygen species-mediated cellular toxicity in dopaminergic SH-SY5Y cells via mitochondria-targeted protective mechanism. Brain Research, 1254, 18 – 27.
Niki, E. (2010). Assessment of antioxidant capacity in vitro and in vivo. Free Radical Biology and Medicine, 49(4), 503 – 515.
O’Connor, I., & O’Brien, N. (1998). Modulation of UVA light-induced oxidative stress by beta-carotene, lutein and astaxanthin in cultured fibroblasts. Journal of Dermatological Science, 16(3), 226 – 230.
Pan, A., Chen, M., Chowdhury, R., Wu, J. H., Sun, Q., Campos, H., Mozaffarian, D., & Hu, F. B. (2012). α-Linolenic acid and risk of cardiovascular disease: A systematic review and meta-analysis. American Journal of Clinical Nutrition, 96(6), 1262 – 1273.
Pandel, R., Poljšak, B., Godic, A., & Dahmane, R. (2013). Skin photoaging and the role of antioxidants in its prevention. ISRN Dermatology, 2013, 930164.
Park, J. S., Chyun, J. H., Kim, Y. K., Line, L. L., & Chew, B. P. (2010). Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutrition & Metabolism, 7(1), 18.
Sies, H. (1985). Oxidative stress: Introduction. Oxidative Stress, 1 – 8.
Sies, H. (1997). Oxidative stress: Oxidants and antioxidants. Experimental Physiology, 82(2), 291 – 295.
Sies, H. (2015). Oxidative stress: A concept in redox biology and medicine. Redox Biology, 4, 180 – 183.
Simopoulos, A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365 – 379.
Steinberg, D. (1997). Low density lipoprotein oxidation and its pathobiological significance. Journal of Biological Chemistry, 272(34), 20963 – 20966.
Tominaga, K., Hongo, N., Karato, M., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on human subjects. Acta Biochimica Polonica, 59(1), 43 – 47.
Tominaga, K., Hongo, N., Fujishita, M., Takahashi, Y., & Adachi, Y. (2017). Protective effects of astaxanthin on skin deterioration. Journal of Clinical Biochemistry and Nutrition, 61(1), 33 – 39.
Valko, M., Leibfritz, D., Moncol, J., Cronin, M. T., Mazur, M., & Telser, J. (2007). Free radicals and antioxidants in normal physiological functions and human disease. International Journal of Biochemistry & Cell Biology, 39(1), 44 – 84.
Valko, M., Rhodes, C. J., Moncol, J., Izakovic, M., & Mazur, M. (2006). Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chemico-Biological Interactions, 160(1), 1 – 40.
Yamashita, E. (2006). The effects of a dietary supplement containing astaxanthin on skin condition. Carotenoid Science, 10, 91 – 95.
Yoshida, H., Yanai, H., Ito, K., Tomono, Y., Koikeda, T., Tsukahara, H., & Tada, N. (2010). Administration of natural astaxanthin increases serum HDL-cholesterol and adiponectin in subjects with mild hyperlipidemia. Atherosclerosis, 209(2), 520 – 523.
Zhang, X., Pan, H., Wang, H., Ma, L., & Guo, R. (2011). Protective effects of astaxanthin on oxidative stress induced mitochondrial dysfunction – A mini-review. Journal of Nutritional Biochemistry, 22(12), 1088 – 1096.
Zhao, G., Etherton, T. D., Martin, K. R., West, S. G., Gillies, P. J., & Kris-Etherton, P. M. (2004). Dietary α-linolenic acid reduces inflammatory and lipid cardiovascular risk factors. Journal of Nutrition, 134(11), 2991 – 2997.

Knowledge Summary of Chapter 6:
* The Diagnosis: [The Dermal Architecture Collapse] and Peripheral Nutrient Abandonment.
– Clinical Manifestation: Visible photo-aging, deep topography alterations (wrinkles), loss of dermal recoil, and chronic xerosis resulting from a “broken” lipid seal.
– Biological Context: The skin is identified as the outermost “expendable” organ. It exists in a state of chronic starvation due to its position at the bottom of the systemic nutrient hierarchy.
– Symptom Industry Failure: Traditional dermatology treats these markers as surface defects, ignoring the underlying metabolic sequestration that prevents internal repair.
* The Root Cause: [The Biological Triage Theory] and the 4mg Placebo Threshold.
– Evolutionary Hierarchy: The body functions as a survival-first machine, allocating circulating antioxidants based on metabolic demand rather than aesthetic preference.
– The Internal Vacuum:
– The Neural Hijack: The Central Nervous System (CNS) consumes 20% of the body’s energy. It intercepts circulating Astaxanthin to neutralize the massive ROS generated by cognitive processes and prevent neuronal apoptosis.
– The Cardiac Drain: The myocardium, possessing the body’s highest mitochondrial density, siphons available molecules to maintain ATP production and protect against oxidative fatigue.
– The Triage Failure: A standard 4mg dose is mathematically exhausted by these “first-strike” organs. By the time blood reaches the dermal microcirculation, the molecular payload is depleted, leaving the skin defenseless against environmental radiation.
* The Mechanism: [The Endogenous Sunshield] vs. [The Neuro-Endocrine Storm].
– 1. Systemic Saturation Physics: The 16mg protocol creates a molecular surplus. Once the lipid membranes of the brain and heart reach their saturation threshold, the “biological blockade” breaks, forcing an overflow into the peripheral tissues.
– 2. 30-Angstrom Transmembrane Anchoring: Unlike floating antioxidants, the specific structure of Astaxanthin allows it to span the entire lipid bilayer (30-Angstroms). It anchors vertically, providing a thermodynamic shield that dissipates photon energy before it shatters cellular DNA.
– 3. Lipidomic Seal Engineering (LA/OA):
– Linoleic Acid (LA) acts as the essential precursor for O-acylceramide synthesis, the “molecular mortar” of the skin barrier.
– Oleic Acid (OA) stabilizes the phospholipid membranes, physically sealing the leaks that cause Transepidermal Water Loss (TEWL).
– 4. Enzymatic Suppression (MMP Inhibition): High-dose Astaxanthin and EPA suppress the expression of Matrix Metalloproteinases (MMPs). This halts the “biochemical shredding” of the collagen scaffold that typically follows UV exposure.
* The Keyora Solution: [The 16mg High-Dose Protocol] and [The Trust Algorithm].
– The Formula: Integration of 16mg Natural Astaxanthin with a Bioactive Carrier (Flaxseed Oil providing ALA, LA, and OA).
– The Evidence: Validated by peer-reviewed human RCTs (Tominaga et al., 2012; Yamashita, 2006). Clinical outcomes include a statistically significant reduction in wrinkle width, restoration of elasticity, and a profound drop in TEWL values.
– The Philosophy: [Regulation, Not Sedation]. Keyora does not mask aging with topical placebos; it re-engineers the body’s internal defensive architecture to reclaim [Aesthetic Sovereignty].
– The Algorithm: A cognitive filter for the high-performer to reject isolated, low-dose ingredients in favor of systemic, lipid-stabilized, and clinically-dosed interventions.

Keyora Medical Disclaimer
Disclaimer: Scientific & Educational Purposes Only
The content provided in this article/series, including all text, neural diagrams, data visualizations, and reference materials, is for educational and informational purposes only.
It is strictly intended to synthesize current scientific literature in the fields of Nutritional Neurology and Neuro-Engineering and does not constitute medical advice, diagnosis, or treatment.
Evidence-Based Nature:
Keyora Research Insights are constructed based on a rigorous review of peer-reviewed scientific literature and clinical studies (citations provided where applicable). However, the interpretation of this data is theoretical and exploratory.
Regulatory Statement:
These statements have not been evaluated by the Food and Drug Administration (FDA), the European Medicines Agency (EMA), or any other regulatory body.
Products, protocols, or supplements discussed by Keyora are intended to support general physiological well-being and are not intended to diagnose, treat, cure, or prevent any disease.
Professional Consultation:
Individual biological responses vary. Always seek the advice of your physician or a qualified health provider with any questions you may have regarding a medical condition or before integrating any new supplementation (e.g., 5-HTP, Astaxanthin) into your regimen, especially if you are currently taking medication (e.g., SSRIs).
Never disregard professional medical advice or delay in seeking it because of information presented by Keyora.

By Keyora Research Notes Series
This article contributes to Keyora’s ongoing scientific documentation series, which systematically outlines the conceptual foundations, mechanistic pathways, and empirical evidence informing our research and development approach.
ORCID: 0009–0007–5798–1996