Keyora Astaxanthin EP-27: The Ovarian Time Machine: The Physics Of Halting Reproductive Aging
By Keyora Research Notes Series
This article contributes to Keyora’s ongoing scientific documentation series, which systematically outlines the conceptual foundations, mechanistic pathways, and empirical evidence informing our research and development approach.
ORCID: 0009–0007–5798–1996

The Transition To Female Sovereignty
Moving Beyond The Male Parameters To Forensically Examine The Biophysical Architecture Of The Ovarian Reserve.
The clinical verdict on male reproductive restoration is complete.
We documented the objective efficacy of thermodynamic shielding and lipidomic reconfiguration in the spermatozoon.
We observed the precise cessation of membrane degradation when exposed to targeted nutritional intervention.
We must now shift our forensic lens.
We turn to the Female Sovereignty module. The female reproductive axis presents a vastly different biophysical landscape. It is governed by strict, unforgiving timelines. It operates on finite cellular reserves established before birth.
However, the underlying physical laws remain identical. Cellular viability is dictated by mitochondrial function and ATP throughput. It is governed by phospholipid membrane integrity.
We must establish a clinical, objective framework to evaluate ovarian health. The oocyte is not an unsolvable biological mystery.
It is a highly complex, energy – demanding biological machine.
It requires structural reinforcement. It demands architectural support against the relentless friction of time.

1. The End Of The Male Module
Applying Established Biophysical Principles To A New Cellular Environment.
The transition from continuous spermatogenesis to the finite ovarian reserve demands a fundamental recalibration of our therapeutic parameters.
The foundational physics of lipid peroxidation, however, do not change.
An unprotected lipid membrane will inevitably oxidize.
We must apply our rigorously established biophysical principles to this new, highly guarded cellular environment.
I. The Universal Threat:
Reactive oxygen species do not discriminate by gender.
Oxidative stress is a universal cellular disruptor.
A rogue, unpaired electron will tear through an oocyte membrane just as violently as it shreds a sperm membrane.
The physics of a radical chain reaction are absolute and mathematically predictable. They require immediate, precise neutralization at the molecular level.
II. The Membrane Vulnerability:
Both spermatozoa and oocytes rely heavily on highly unsaturated phospholipid bilayers. These complex lipid structures provide the essential fluidity required for the mechanical fusion of fertilization.
However, this dense concentration of double bonds creates an inherent, unavoidable vulnerability.
Both gametes are uniquely susceptible to catastrophic lipid peroxidation. The resulting structural collapse and loss of membrane potential is identical in both biological contexts.
III. The Shift In Focus:
We now transition our focus from the continuous, highly regenerative production cycle of the testes to the finite, steadily depleting reserve of the ovaries.
The male reproductive system operates as an ongoing biological assembly line.
The female system operates as a heavily fortified, static vault.
We are no longer repairing a dynamic factory. We are deploying structural defenses to protect a dormant fortress.
IV. The New Clinical Objective:
The clinical objective is no longer to rebuild a continuous biological supply line from scratch.
The primary objective is to preserve, fortify, and optimize an existing, highly vulnerable biological reserve.
We must deploy an advanced thermodynamic shield to protect the remaining cellular inventory.
We must aggressively mitigate the biophysical decline of the surrounding follicular architecture to prevent premature apoptosis.

2. The Misconception Of Inevitable Decline
Rejecting The Abstract Concept Of The Biological Clock.
The mainstream clinical dialogue surrounds ovarian aging with an impenetrable veil of fatalism. It is spoken of in hushed, defeated tones in consultation rooms worldwide. This represents a profound scientific failure of imagination.
We must strip away the emotion and the psychological weight. We must examine the raw, unforgiving physics of follicular senescence.
I. The Clinical Dogma:
Traditional reproductive medicine often treats ovarian aging as an abstract, unavoidable ticking clock. It is viewed as an absolute, untouchable chronological certainty.
Patients are routinely told their ovarian reserve is simply depleted due to age. The actual biophysical mechanism of this depletion is rarely explained or investigated. It is passively accepted as a fixed biological destiny.
II. The Keyora Rejection:
The Keyora research protocol firmly rejects this abstract, unscientific fatalism. Cellular aging is not a mystical process. It is a measurable sequence of physical and biochemical degradation.
What standard clinical medicine loosely terms age – related decline, Keyora explicitly defines as [The Follicular Oxidative Cascade].
We track the exact chronological loss of mitochondrial efficiency.
We measure the exact, cumulative accumulation of intracellular reactive oxygen species.
III. The Microenvironmental Reality:
The functional decline in oocyte quality is heavily, if not entirely, influenced by the progressive deterioration of the surrounding ovarian microenvironment.
The surrounding granulosa cells act as the critical life support and nutrient filtration system for the oocyte.
When these supporting somatic cells suffer extensive oxidative damage, the central oocyte is functionally starved.
The surrounding follicular fluid transitions from a nourishing incubator into a highly toxic, inflammatory bath.
IV. The Potential For Modulation:
Because this reproductive decline is fundamentally driven by physical, thermodynamic variables, it is entirely subject to biophysical intervention. It can be objectively modulated, slowed, and mitigated through targeted nutritional engineering.
We possess the molecular tools to restore the antioxidant capacity of the follicular fluid.
We can structurally reinforce the mitochondrial walls of the granulosa cells, thereby preserving the oocyte within.

3. The Oocyte Vulnerability
The Unique Biophysical Demands Of The Female Gamete.
To effectively engineer a protocol to protect the oocyte, we must first deeply understand its extreme physiological burden.
It is by far the largest single cell in the human body.
It is concurrently the most metabolically and energetically demanding.
This unique combination creates a highly precarious bioenergetic reality that demands constant defense.
I. The Extended Lifespan:
Human oocytes remain arrested in the dictyate stage within the ovaries for decades. This prolonged state of biological stasis exposes them to twenty, thirty, or forty years of cumulative environmental stress.
Every systemic inflammatory spike, every exposure to heavy metals or toxic load, leaves a microscopic biochemical scar. The female gamete must survive a lifetime of intense oxidative crossfire before it is ever deployed for ovulation.
II. The Mitochondrial Density:
The mature oocyte contains significantly more mitochondria than any other somatic or germ cell in the human body. A single, healthy mature oocyte can house well over one hundred thousand individual mitochondria.
While necessary for energy, this density creates a massive potential for endogenous oxidative leakage. It is the biological equivalent of a heavy engine running continuously for decades, generating constant cellular exhaust.
III. The Energy Requirement:
The sequential processes of meiotic maturation, fertilization, and early embryonic cleavage require extraordinary, uninterrupted volumes of ATP synthesis.
The critical chromosomal separation phase is highly dependent on instantaneous energy availability. A single, microscopic drop in mitochondrial voltage during this phase leads directly to chromosomal non – disjunction and aneuploidy.
The oocyte simply cannot tolerate bioenergetic power failures.
IV. The Target Established:
The fundamental reality is clear. The oocyte is highly dense in mitochondria and entirely dependent on pristine, uninterrupted cellular energy production. It requires an absolute, unyielding thermodynamic protection protocol.
We must now turn our attention to examine the specific environmental and physiological factors that threaten this delicate system. We must precisely map the exact coordinates of oxidative failure within the ovary.
Keyora Insight:
The ovarian reserve is not an abstract hourglass running out of sand. It is a biological vault under constant thermodynamic siege.
By shifting our focus from the chronological age of the patient to the objective biophysical integrity of the follicular microenvironment, we reclaim control. We stop passively watching the clock, and we begin actively reinforcing the shield.

The Oxidative Reality And The Dietary Variable
Quantifying The Accumulation Of Reactive Oxygen Species And Identifying Dietary Lipid Imbalance As A Key Exacerbating Factor In Ovarian Decline.
To modulate ovarian aging, we must precisely identify the mechanisms of its degradation. The primary driver of this cellular decline is the progressive accumulation of reactive oxygen species within the follicular fluid.
This oxidative stress does not occur in a vacuum. It is heavily influenced by systemic factors.
In the Keyora paradigm, we recognize the modern fifteen to one dietary lipid imbalance not as an absolute determinant, but as a critical environmental variable.
This surplus of omega six fatty acids significantly exacerbates localized inflammation. It heightens the susceptibility of the oocyte to catastrophic lipid peroxidation.
The resulting structural damage operates on a principle of exponential biochemical synergy.
The cumulative biophysical breakdown is mathematically precise.
In the context of lipid degradation, the equation 1+1+1+1+1+1+1 > 7 defines this synergistic cellular destruction.
We must examine this biophysical reality.
We must map the exact coordinates of this structural collapse.

1. The ROS Accumulation
The Progressive Toxicity Of The Follicular Microenvironment.
The ovarian reserve exists within an enclosed fluid ecosystem. This follicular fluid acts as a biological incubator. It transfers essential nutrients to the developing gamete.
However, this fluid is highly susceptible to chemical alteration.
We must analyze the precise origins of this cumulative toxicity.
A. The Endogenous Generation:
Normal ovulation requires intense cellular metabolism. Steroidogenesis naturally generates reactive oxygen species as a continuous chemical byproduct.
The mitochondria produce superoxide anions during normal respiration. This is a baseline physiological process. It is the inescapable cost of cellular energy production.
B. The Exogenous Burden:
External variables forcefully multiply this baseline production.
Environmental toxins penetrate the delicate follicular barrier.
Psychological stress triggers severe neuroendocrine fluctuations.
Chronological aging steadily degrades cellular repair mechanisms. These combined forces dramatically increase the overall oxidative burden over time.
C. The Overwhelmed Defenses:
Biological systems possess finite defensive capacities.
Eventually, the endogenous antioxidant enzymes within the follicular fluid become saturated.
Superoxide dismutase and glutathione peroxidase reach their absolute maximum catalytic velocity. They can no longer neutralize the incoming superoxide anions. The biological shield fractures.
D. The Oxidative Shift:
The physical environment undergoes a rapid phase transition.
The microenvironment shifts from a balanced redox state to a chronic oxidative fire. It becomes a low – grade, relentless inflammatory burn.
The protective incubator is now a highly toxic chemical bath.
The oocyte is left entirely exposed to the elements.

2. The 15-20 : 1 Environmental Variable
How Dietary Lipid Imbalance Exacerbates Cellular Vulnerability.
Oxidative stress requires biological fuel to cause structural damage. The modern dietary lipid ratio provides this highly combustible fuel.
We must objectively explain the mechanics of this environmental variable.
This is not a dietary trend. It is a fundamental biophysical vulnerability that compromises the entire system.
A. The Systemic Inflammation:
The modern human consumes a massive excess of linoleic acid. A fifteen to one ratio of omega six to omega three fatty acids creates constant systemic biological tension.
It acts as a systemic precursor to pro – inflammatory eicosanoids.
It shifts the entire organism into a state of high chemical reactivity.
B. The Localized Impact:
Systemic parameters always dictate local microenvironments.
This systemic inflammatory tone directly penetrates the dense ovarian capsule. It translates into localized low – grade inflammation within the ovarian tissue.
This constant inflammatory signaling forces the surrounding cells to further increase their baseline generation of reactive oxygen species.
C. The Structural Incorporation:
Cells build their structural walls from available dietary materials. Excess arachidonic acid is physically incorporated into the cellular architecture.
It embeds into the plasma membranes of the granulosa cells.
It is forcefully integrated into the delicate membrane of the oocyte itself.
The structural composition is fundamentally altered.
D. The Heightened Susceptibility:
These omega six lipids possess multiple double bonds. These rigid, highly unsaturated structures are chemically unstable.
They are prime biophysical targets for destructive hydroxyl radicals.
They act as highly combustible fuel for the internal oxidative fire. The cellular membrane is transformed into a terminal chemical liability.

3. The Sub-Cellular Consequence
The Physical Degradation Of The Biological Reserve.
We have established the accumulation of free radicals.
We have identified the integration of combustible lipid fuel.
We must now summarize the terminal biophysical damage to the female gamete. This is the exact mechanical sequence of structural failure within the ovary.
A. The Lipid Peroxidation Cascade:
Reactive oxygen species directly attack the vulnerable cellular membranes. They strip an electron from the unstable lipid chains.
This initiates a violent, self – propagating chain reaction. This is the lipid peroxidation cascade. It physically shreds the protective phospholipid bilayer.
B. The Mitochondrial Impairment:
This violent peroxidation penetrates deep into the cell interior. It damages the inner mitochondrial membrane of the oocyte. The electron transport chain loses physical integrity.
This severely limits total adenosine triphosphate output. The cell rapidly loses its primary energy source.
C. The Granulosa Cell Apoptosis:
The oocyte does not die alone. The surrounding granulosa cells provide critical metabolic support. They are equally vulnerable to this severe oxidative structural damage.
The intense lipid peroxidation triggers programmed cell death.
The granulosa cells undergo widespread, irreversible apoptosis.
D. The Decline In Quality:
The final physiological outcome is absolute.
The devastating combination of adenosine triphosphate depletion and the total loss of cellular support directly compromises maturation.
The gamete cannot properly segregate chromosomes. This is the objective, biophysical reality of ovarian aging.

The Demand For Objective Biophysical Support
Establishing The Absolute Necessity Of The Astaxanthin Vanguard As The Prerequisite For Executing Targeted Lipidomic Reconfiguration In The Ovary.
The biophysical pathology of ovarian decline is fundamentally clear.
The oocyte and its critical supporting granulosa cells are under a continuous, unrelenting oxidative siege.
This cellular siege is aggressively exacerbated by profound systemic lipid imbalances across the entire organism.
To effectively mitigate this precise biological decline, we cannot rely on standard, generic nutritional support.
Standard water – soluble vitamins cannot physically penetrate the deep lipid structures of the cellular mitochondrial engine.
The Keyora protocol demands a highly targeted, intensely lipophilic biophysical intervention.
We must actively deploy the sixteen milligram Astaxanthin vanguard. The objective is to establish an absolute, impenetrable thermodynamic shield around the vulnerable gamete.
This biophysical shield is not the final cure for reproductive aging. It is the fundamental, non – negotiable physical prerequisite required to safely deploy the 1+1+1+1+1+1+1 > 7 matrix.
We must definitively extinguish the chemical fire before we attempt to rebuild the fragile biological architecture.

1. The Failure Of Standard Antioxidants
The Pharmacokinetic Limitations Of Generic Interventions.
Mainstream clinical interventions routinely fail due to basic, fundamental pharmacokinetic incompatibilities.
We must forensically analyze exactly why standard over – the – counter vitamins fail to operate within the specific, heavily guarded ovarian microenvironment.
Firstly, The Hydrophilic Barrier:
Standard biological antioxidants like Vitamin C are strictly water – soluble molecules. They circulate freely and effectively within the aqueous environment of the blood plasma.
However, they are violently, physically repelled by the highly dense lipid membranes of the ovarian cells. They simply cannot penetrate the thick phospholipid bilayer. They bounce off the cellular exterior.
Secondly, The Mitochondrial Isolation:
Because they cannot penetrate the cellular wall, they cannot cross the critical outer and inner mitochondrial membranes. This represents a catastrophic failure in targeted payload delivery.
They cannot reach the exact intracellular site of continuous superoxide generation. The critical mitochondrial matrix is left completely isolated and entirely unprotected from internal oxidative exhaust.
Thirdly, The Pro – Oxidant Risk:
Conventional molecules like Vitamin E operate by donating a single electron to neutralize circulating free radicals. Under states of high, chronic oxidative stress, they rapidly become exhausted.
Without immediate enzymatic regeneration, they themselves mutate into highly unstable free radicals. They trigger severe pro – oxidant phase transitions that accelerate cellular apoptosis.
Fourthly, The Need For A Lipophilic Anchor:
Effective ovarian protection requires absolute, unwavering structural stability.
It demands a specialized molecule that can physically dissolve deep into the dense cellular lipid bilayer.
It must possess the molecular capacity to safely dissipate massive thermal energy without undergoing structural degradation itself.
It must act as a permanent anchor.

2. The Thermodynamic Shield Prerequisite
Deploying The 16mg Astaxanthin Vanguard.
We must introduce the definitive molecular solution to this pharmacokinetic failure. We deploy the sixteen milligram Astaxanthin vanguard. It provides the exact, necessary biophysical specifications required for complete cellular protection and radical neutralization.
Firstly, The Structural Penetration:
Astaxanthin is an extremely unique, highly lipophilic molecule.
It possesses a precise, trans – membrane geometric structure.
It easily and rapidly penetrates the viscous, enclosed fluid of the follicular microenvironment.
It anchors directly and permanently into the cellular membranes of the oocyte and the surrounding granulosa cells.
Secondly, The Electron Resonance:
Its structural framework features a massive, continuous thirty – Angstrom conjugated double – bond system. This rigid structure forms a dense, protective electron cloud across the entire cell wall.
It actively intercepts highly volatile hydroxyl radicals.
It absorbs their kinetic impact and dissipates their destructive energy harmlessly into the surrounding fluid as thermal heat.
Thirdly, The Halting Of Peroxidation:
This continuous, unbroken quenching process is mathematically precise.
It physically halts the catastrophic lipid peroxidation cascade exactly in its tracks.
It effectively shields and protects the highly vulnerable mitochondrial cardiolipin matrix from immediate structural destruction. The biological energy engine is secured.
Fourthly, The Safe Zone Established:
The primary tactical objective of the intervention is fully achieved.
The sixteen milligram vanguard successfully extinguishes the localized, chronic inflammatory fire.
It creates an absolute biochemical safe zone within the highly vulnerable, previously hostile ovarian microenvironment.
The groundwork is now officially laid.

3. The 1+1+1+1+1+1+1 > 7 Synergistic Mandate
The Requirement For Structural Lipidomic Reconstruction.
With the cellular perimeter secured, structural lipidomic reconstruction is now biologically viable.
We must clearly explain why the complex matrix strictly requires the preliminary deployment of the thermodynamic shield.
Firstly, The Fragility Of Structural Lipids:
To physically reverse the massive dietary fifteen to one inflammatory damage, we must deliver highly fluid structural lipids to the cell. These vital molecules are heavily polyunsaturated.
This specific chemical structure means they are inherently extremely fragile. They are highly prone to rapid, catastrophic cellular oxidation.
Secondly, The Futility Of Unshielded Delivery:
Administering these vital, fragile lipids directly into an unquenched, high – ROS ovarian environment is a clinical and mathematical failure.
It guarantees their immediate physical destruction before they can be utilized. They will simply act as fresh combustible fuel for the existing internal oxidative fire. The cellular damage will simply accelerate.
Thirdly, The Astaxanthin Escort:
The synergistic logic of the protocol is absolute and completely irreversible.
Because Astaxanthin has definitively established the thermodynamic safe zone, the restorative lipids can survive the biological transit.
The complex 1+1+1+1+1+1+1 > 7 matrix can now safely enter the follicular microenvironment without being instantly oxidized upon arrival.
Fourthly, The Path To Restoration:
The thermodynamic shield is the absolute physical prerequisite for the successful deployment of the nutritional matrix. They cannot exist or function properly in biological isolation.
Together, they form the unshakable foundation of the Ovarian Time Machine. In Chapter 1, we will forensically dissect the precise anatomy of this vulnerable microenvironment.

Chapter 1: The Root Of Ovarian Decline:
The Microenvironmental Collapse
How reactive oxygen species and the 15:1 dietary variable objectively disrupt the follicular fluid matrix
To objectively modulate ovarian aging, we must first forensically deconstruct the physical environment in which it occurs. The ovary is not a homogenous organ.
It is a highly stratified, complex microenvironment.
It houses the finite biological reserve of the female reproductive system.
Unlike spermatogenesis, which is continuous, folliculogenesis relies on a depleting pool of dormant cells. This fundamental anatomical difference dictates a unique set of biophysical vulnerabilities.
We must map the precise architecture of the ovarian cortex and the follicular unit before we can deploy the Astaxanthin vanguard and the 1+1+1+1+1+1+1 > 7 matrix. The physical tissue parameters govern the absolute biological outcome.
We cannot intervene without a clear map of the cellular terrain.
We must understand the exact lipidomic composition of the organ.
We must observe the distribution of mitochondria within the stroma.
The physical barriers dictate the specific rate of oxidative infiltration.
The structural boundaries determine the success or failure of our targeted nutritional payload.
This anatomical baseline is completely non – negotiable. It is the absolute foundation for the protocols that follow. Every intervention must be mapped directly to a specific anatomical coordinate.

1. The Ovarian Cortex
The Biological Vault Of The Primordial Reserve.
We begin our forensic analysis at the outermost functional layer.
The architecture of the ovary is defined by its extreme spatial organization.
The most critical cellular assets are isolated within specific structural zones.
This is not a random distribution of biological material. It is a highly engineered vault.
We must examine the biophysical properties of this protective outer shell.
It is the first line of defense against systemic oxidative stress.
It is also the primary site of cumulative cellular degradation.
I. The Outer Stratum:
The ovarian cortex is the dense outer layer of the ovary.
It serves as the physical vault for the female genetic payload. This layer is composed of highly specialized connective tissue.
It forms a rigid, protective extracellular matrix. The structural integrity of this matrix is absolutely paramount.
It must maintain strict mechanical stability.
It must resist external inflammatory pressure. The cortex acts as a biological shock absorber.
It buffers the internal cellular reserve against systemic metabolic fluctuations. A compromised cortex immediately exposes the vulnerable inner structures.
We observe clear correlations between cortical tissue stiffness and local oxidative damage. The physical deterioration of this layer is the primary indicator of regional biological decline.
II. The Primordial Pool:
This region houses hundreds of thousands of primordial follicles. These highly specialized structures were completely formed during early fetal development. They represent the absolute limit of the reproductive lifespan.
The biological inventory is fixed before birth. There is absolutely no ongoing generative capacity within this pool.
Each follicle is an isolated, dormant biological unit. It contains a precise package of genetic and bioenergetic material. The spatial distribution of this pool is dense. This high cellular density creates localized metabolic micro – zones. These zones require precise, uninterrupted oxygen diffusion.
Any disruption in microvascular supply triggers immediate, localized cellular apoptosis. The preservation of this finite pool is the ultimate goal of our thermodynamic intervention.
III. The Arrested State:
The oocytes within these primordial follicles remain arrested in prophase I of meiosis. They can remain in this profound dormant state for decades. This is an extreme state of suspended biological animation.
The cellular metabolism operates at a microscopic baseline.
The mitochondrial electron transport chain is heavily suppressed. This deliberate suppression minimizes the production of endogenous free radicals.
However, this stasis requires constant, active biological suppression mechanisms.
The cells must continuously maintain exact osmotic gradients. They must actively repair spontaneous DNA damage. This baseline maintenance requires a constant, albeit low, supply of adenosine triphosphate.
The arrested state is not truly passive. It is an active, heavily regulated biophysical holding pattern.
IV. The Cumulative Exposure:
This prolonged chronological arrest means the cortex is continuously exposed to cumulative environmental and oxidative stressors over the entire lifespan. The protective vault is under constant chemical siege.
Decades of systemic inflammation leave microscopic scars. Circulating reactive oxygen species continuously bombard the cortical boundary.
Every surge in cortisol or insulin generates a localized inflammatory echo. The lipid membranes of the dormant cells slowly accumulate oxidized phospholipids. The cumulative burden of this exposure is massive. It creates a highly precarious biological baseline.
The cells are forced to expend their extremely limited energy reserves on endless defensive repairs. This slow, relentless thermodynamic friction is the true driver of age – related structural depletion.

2. The Follicular Unit
The Fundamental Building Block Of Reproduction.
We must now zoom in to the microscopic level.
We move from the macroscopic tissue down to the individual functional module. The follicular unit is the core machinery of the female reproductive axis. It is a highly specialized, multicellular organism within an organ.
We must deconstruct its precise cellular anatomy.
We must identify the exact mechanisms of internal communication. This unit operates as a strict, interdependent bioenergetic cooperative. The failure of one cellular component guarantees the collapse of the entire unit.
I. The Oocyte Core:
At the heart of every functional follicle lies a single oocyte. This is the highly complex, mitochondria – dense female gamete.
It is the absolute center of the biological reproductive universe.
It is simultaneously the most important and the most biophysically fragile cell in the human body.
The oocyte contains a massive, dense network of mitochondria. These organelles are arranged in strict geometric patterns around the nucleus. They represent an enormous reservoir of potential energy.
However, this density also represents a catastrophic oxidative liability. The internal lipid membranes of the oocyte are heavily composed of polyunsaturated fatty acids. This specific lipidomic profile makes the cell extraordinarily vulnerable to violent lipid peroxidation.
II. The Somatic Cell Envelope:
The oocyte is physically encapsulated by specialized layers of somatic cells. These are primarily the granulosa and thecal cells.
The oocyte cannot survive in biological isolation. It requires constant, direct physical contact with these supporting structures.
The granulosa cells form a tight, concentric sphere around the gamete.
They act as the primary biological filtration system.
They process raw metabolic substrates directly from the bloodstream.
They synthesize essential amino acids and critical signaling molecules.
They physically shuttle these processed nutrients directly into the oocyte via specialized gap junctions.
The structural integrity of this granulosa envelope is completely non – negotiable. It is the absolute life support system for the dormant genetic core.
III. The Basal Lamina:
A specialized extracellular matrix called the basal lamina physically separates the internal granulosa cells from the external vascularized thecal layer. This is a critical anatomical boundary.
It is not a simple physical wall.
It is a highly selective, semi – permeable biological filter.
It dictates exactly which molecules can enter the internal follicular microenvironment.
It actively repels large immune cells and massive inflammatory cytokines.
It allows the passive diffusion of strictly regulated nutrients and hormones.
The precise thickness and chemical composition of this lamina are essential for maintaining internal homeostasis. If oxidative stress degrades this matrix, the selective barrier fails.
Toxic systemic factors immediately flood the internal chamber.
IV. The Avascular Compartment:
The internal architecture presents a stark structural reality.
The inner follicular compartment is strictly avascular. There are absolutely no blood vessels penetrating the granulosa layer.
The oocyte relies entirely and exclusively on the surrounding somatic cells for all nutrient diffusion and metabolic support. It has zero direct access to systemic circulation. It cannot directly absorb oxygen or glucose from the blood. This creates an extreme biophysical dependency.
If the granulosa cells undergo oxidative apoptosis, the oocyte is instantly starved. The gamete suffocates within its own biological vault.
This absolute avascular dependency is the primary reason we must deploy our 1+1+1+1+1+1+1 > 7 matrix to protect the critical somatic support cells.

3. The Demand For Homeostasis
The Strict Biophysical Requirements For Cellular Maturation.
The static architecture of the dormant follicle is only the baseline.
The true biophysical challenge occurs during the activation phase.
The transition from dormancy to active maturation is a violent bioenergetic event.
It requires absolute structural stability.
It demands a perfect, uncorrupted microenvironment.
We must map the thermodynamic parameters of this critical phase transition.
I. The Maturation Trigger:
Periodically, specific cohorts of primordial follicles are recruited into the active growing pool. This strictly controlled biological process forces the resumption of their heavy metabolic activity.
The exact trigger mechanism involves complex local and systemic hormonal signaling.
The dormant cells rapidly awake from decades of stasis. They must immediately resume massive protein synthesis.
The gap junctions between the granulosa cells and the oocyte widen.
The flow of metabolites increases exponentially.
This sudden influx of molecular traffic puts immense stress on the cellular architecture.
The cells must rapidly expand their physical volume while maintaining absolute structural integrity.
II. The Bioenergetic Surge:
This sudden resumption of meiosis requires an extraordinary, sustained surge in mitochondrial adenosine triphosphate synthesis within the oocyte.
The internal energy demand skyrockets.
The suppressed electron transport chains are thrown into maximum overdrive.
The mitochondria must consume massive volumes of oxygen. This hyper – active metabolic state generates an unavoidable spike in endogenous superoxide production.
The cell is deliberately pushing its own thermodynamic limits. This controlled burn is absolutely necessary for chromosomal segregation.
However, it creates a massive, immediate risk of internal oxidative structural collapse. The endogenous antioxidant defenses are pushed to their absolute maximum physiological capacity.
III. The Microenvironmental Sensitivity:
This intricate, high – speed process of maturation is exquisitely sensitive to the surrounding biochemical and physical environment. The fluid filling the growing follicle must remain pristine.
Any slight shift in local pH disrupts critical enzymatic reactions. A minor increase in external reactive oxygen species immediately degrades the delicate spindle fibers required for genetic division.
The presence of oxidized lipid byproducts in the follicular fluid acts as a direct toxic inhibitor.
The maturing gamete operates with zero margin for biochemical error.
The slightest microenvironmental disturbance results in catastrophic meiotic failure and immediate cellular apoptosis.
IV. The Thermodynamic Imperative:
To successfully transition from a dormant state to a fully mature ovum, the follicular unit demands absolute thermodynamic stability and lipidomic integrity.
We cannot alter the fundamental metabolic requirements of this process.
We can only reinforce the biological machinery.
We must pre – load the local environment with our specialized antioxidant vanguard.
We must ensure the physical cell walls are constructed with highly stable, functional lipids. This is the ultimate objective.
We deploy the 1+1+1+1+1+1+1 > 7 matrix to engineer a microenvironment capable of withstanding the violent bioenergetic surge of maturation.
This proactive structural reinforcement is the only viable path to mitigating reproductive decline.

1.1 The Oocyte – Granulosa Cell Crosstalk
Forensically Examining The Absolute Biophysical Dependency And The Physical Communication Networks Within The Developing Follicle.
The oocyte does not mature in biological isolation. It cannot survive without a highly engineered physical support structure.
Because the inner compartment of the follicle entirely lacks direct vascularization, the oocyte is completely dependent on its surrounding microenvironment. This biological network is primarily composed of thousands of highly specialized granulosa cells. The relationship between the central oocyte and the massive granulosa cell envelope is one of the most intimate symbiotic partnerships in human biology.
They physically connect at a microscopic, structural level.
They continuously share critical metabolic substrates.
They exchange constant regulatory chemical signals to maintain strict homeostasis.
To understand exactly how systemic oxidative stress disrupts female fertility, we must first forensically deconstruct the precise physical architecture of this cellular crosstalk.
We must observe exactly how molecular fuel is actively transported across these biological membranes.
We must map the exact physical bridges that sustain gamete life. The ovarian vault is not simply a dormant biological storage container. It is a highly active, metabolically demanding biophysical reactor. The granulosa cells serve as the primary thermodynamic engine for this entire reactor.
They filter all systemic vascular inputs. They strictly regulate the local fluid dynamics.
They dictate the exact bioenergetic status of the resting gamete.
Any microscopic disruption to this local granulosa network immediately threatens the absolute viability of the entire reproductive cycle. The physical architecture demands absolute stability.

1. The Symbiotic Relationship
The Mutual Dependency Of The Follicular Unit.
The interaction between the oocyte and its surrounding granulosa envelope is not a passive fluid exchange. It is a highly regulated, active biochemical and physical dependency.
Both cellular entities strictly require the continuous presence and functional competence of the other. The biological unit operates strictly as a single thermodynamic system.
A. The Paracrine Signaling:
The oocyte and granulosa cells communicate continuously via complex, bidirectional paracrine signaling factors. These highly specific molecules direct precise cellular differentiation and structural proliferation. They govern the exact spatial physical organization of the maturing follicle.
This localized signaling chemical gradient must remain completely uninterrupted. It requires absolute follicular fluid purity to function properly. Oxidative noise instantly corrupts these delicate signaling pathways.
The communication network breaks down completely.
The cells lose their synchronized biological rhythm.
B. The Oocyte Control:
The oocyte is not a passive passenger within this system. It acts as the central biological command and control center.
The oocyte actively secretes highly specialized factors like growth differentiation factor nine. It continuously secretes bone morphogenetic protein fifteen.
These specific physical proteins dictate the exact metabolic rate of the surrounding granulosa cells. They instruct the somatic cells on precisely what nutrients to process.
The gamete actively controls and micromanages its own life support system.
C. The Granulosa Response:
In direct, localized response to these gamete – derived physical signals, the granulosa cells initiate massive metabolic shifts.
They rapidly synthesize and deliver the exact, specific biochemical substrates required for the oocyte to maintain meiotic progression.
They actively alter their own internal lipidomic profiles to accommodate the massive bioenergetic requirements of the growing oocyte.
They act as the highly efficient biological filtration and processing plant for the inner avascular chamber.
They dedicate their entire biological existence to supporting the gamete.
D. The Coupled Fate:
The functional health of the central oocyte is a direct, objective reflection of the health of the granulosa cell network.
Their biological fates are permanently and physically coupled. If the granulosa cells experience even minor bioenergetic failure, the oocyte instantly suffers the exact identical thermodynamic collapse.
We cannot evaluate or protect the gamete without first securing the somatic envelope. The clinical objective must always focus on the supporting architecture first. The gamete simply reflects the integrity of its physical environment.

2. The Gap Junctions (Connexins)
The Physical Bridges Of Cellular Communication.
Chemical signaling through the follicular fluid is insufficient for maintaining true bioenergetic homeostasis.
The intense metabolic demand of the oocyte requires direct, physical pipelines for rapid substrate transfer.
The biological solution is a highly complex network of microscopic physical bridges.
We must examine the precise anatomy of this connection.
A. The Transzonal Projections:
Granulosa cells do not merely sit adjacent to the oocyte.
They extend long, microscopic physical filaments deep into the central cavity. These complex physical structures are called transzonal projections.
They pierce directly through the thick, highly protective zona pellucida extracellular matrix.
They reach out to physically contact the plasma membrane of the central oocyte.
They form a dense, physical web of structural support. This web physically anchors the gamete in three – dimensional space.
B. The Connexin Proteins:
At the precise terminal tips of these microscopic projections, highly specialized transmembrane proteins are deployed. These are the critical connexin proteins.
They represent the absolute pinnacle of cellular structural engineering.
They rapidly assemble into distinct hexameric molecular ring structures.
They insert directly into the opposing lipid bilayers of both cell membranes.
They act as microscopic biological docking stations.
They physically prepare the delicate membranes for direct localized fusion.
C. The Gap Junction Channels:
These localized pores perfectly align between the two adjacent, apposed cell membranes.
They precisely dock together to create fully functional gap junctions. These are direct, physical tubular channels linking the cytoplasm of the granulosa cell entirely to the cytoplasm of the oocyte.
They create a continuous, uninterrupted intracellular bioenergetic highway.
Molecules can pass freely between the two distinct cellular entities. The structural separation between the cells effectively vanishes.
D. The Structural Prerequisite:
The successful initial formation and sustained structural stability of these gap junctions require absolute plasma membrane integrity.
The lipidomic fluidity of the surrounding phospholipid membrane must be absolutely flawless. If the complex lipid bilayer is oxidized or physically rigid, the delicate connexin proteins cannot properly dock.
The physical bridge instantly collapses.
The vital communication channel is permanently severed.
The oocyte is physically isolated from its critical life support network.

3. The Metabolic Support
The Delivery Of Bioenergetic And Antioxidant Substrates.
The physical construction of the gap junction network serves one primary, non – negotiable biological purpose. It facilitates the massive, unidirectional transfer of bioenergetic fuel and critical protective molecules.
The oocyte is metabolically deficient by evolutionary design. It requires constant, massive external caloric and antioxidant subsidies.
A. The Glycolytic Limitation:
The central oocyte operates with a severe, deliberate, and highly specialized metabolic restriction.
It has an extremely poor innate capacity for standard cellular glycolysis.
It completely lacks the sufficient enzymatic machinery to efficiently process raw systemic glucose into usable biological energy on its own.
It is functionally blind to direct glucose metabolism.
It strictly requires an external cellular intermediary to pre – process all incoming caloric fuel.
B. The Pyruvate Transfer:
The surrounding granulosa cells act as this critical, highly active biochemical intermediary.
They rapidly absorb systemic circulating glucose from the external vascularized environment.
They efficiently metabolize this raw glucose directly into specific, highly usable pyruvate and lactate molecules.
They physically push these processed bioenergetic substrates directly through the physical gap junction channels.
The molecular fuel is injected directly into the waiting oocyte cytoplasm.
The internal supply line is direct and massive.
C. The ATP Generation:
The internal bioenergetic engine of the gamete relies entirely and completely on this continuous, uninterrupted supply chain.
The oocyte possesses a massive network of internal mitochondria which rapidly utilizes this delivered pyruvate.
They heavily process it through the complex electron transport chain.
They synthesize massive, highly concentrated quantities of intracellular adenosine triphosphate. This raw physical ATP energy is the absolute prerequisite for driving successful chromosomal maturation.
D. The Antioxidant Delivery:
This direct physical dependency extends far beyond simple caloric fuel delivery. Granulosa cells continuously and actively synthesize critical endogenous structural antioxidants.
They internally generate massive quantities of complex intracellular glutathione.
They rapidly and continuously transfer this essential glutathione directly to the oocyte via the microscopic gap junction channels.
This constant, pressurized delivery provides the highly vulnerable gamete with its primary, non – negotiable structural defense against baseline mitochondrial oxidation and localized free radical leakage.

4. The Shared Vulnerability
How Localized Stress Collapses The Symbiotic Network.
This highly specialized, interdependent architecture is a bioenergetic marvel.
However, it represents an extreme localized vulnerability. The system operates with absolute zero structural redundancy.
A targeted biophysical attack on the supporting granulosa cell membrane guarantees the complete systemic collapse of the entire reproductive module.
We must track the exact mechanism of this failure.
A. The Membrane Disruption:
The physical structural threat immediately initiates at the vulnerable localized lipid bilayer. If the highly sensitive plasma membranes of the granulosa cells become chemically rigid or structurally oxidized by systemic reactive oxygen species, the entire architecture fails.
The delicate connexin hexamer rings are violently displaced from their specific structural lipid anchors.
The gap junctions physically detach and rapidly disintegrate into the surrounding follicular fluid.
B. The Metabolic Starvation:
The immediate localized thermodynamic consequence is absolute, catastrophic, and completely irreversible.
When the physical transzonal projections retract, the localized biological delivery of metabolic pyruvate and critical glutathione is instantly and permanently severed.
The internal biochemical pipeline violently shuts down.
The central oocyte is immediately starved of vital bioenergetic cellular fuel.
The massive internal mitochondrial network violently stalls. Cellular ATP production rapidly drops to absolute zero.
C. The Oxidative Exposure:
The secondary biological and chemical impact is equally devastating to the reproductive unit.
Stripped completely of its constant, required supply of granulosa – derived glutathione and antioxidants, the central oocyte is left completely physically unprotected. It becomes entirely defenseless against rapidly accumulating intracellular reactive oxygen species.
The unquenched localized oxidative fire immediately and violently shreds the internal delicate chromosomal spindle fibers.
The genetic payload is objectively destroyed.
D. The Clinical Target:
The objective structural preservation of this incredibly fragile oocyte – granulosa crosstalk is our absolute primary clinical target. It heavily dictates the strict, completely non – negotiable necessity of deploying the targeted Astaxanthin vanguard.
We must deploy this powerful structural thermodynamic shield to permanently physically protect the delicate lipid gap junctions from catastrophic oxidative collapse.
This critical stabilization strictly prepares the physical microenvironment for the safe deployment of the exact 1+1+1+1+1+1+1 > 7 matrix to rebuild the architecture.

1.2 The ROS Infiltration In Follicular Fluid
Quantifying The Accumulation Of Reactive Oxygen Species Within The Follicular Microenvironment And Its Objective Impact On Oocyte Viability.
The physical architecture of the oocyte – granulosa network is exceptionally delicate.
It does not exist in a dry vacuum.
It is suspended within a highly specialized, dynamic biological medium.
This medium is the follicular fluid.
This fluid acts as the immediate biochemical environment for the developing gamete.
Under optimal homeostatic conditions, it provides essential nourishment. It delivers strict regulatory signals. It maintains exact osmotic pressure.
However, overwhelming clinical consensus confirms a severe biophysical vulnerability. This fluid is highly susceptible to the rapid accumulation of reactive oxygen species.
We must analyze this infiltration.
When the delicate redox balance tips, the follicular fluid completely transforms. It shifts from a nurturing biological medium into a highly oxidative, toxic chemical bath. The gamete is essentially drowning in metabolic exhaust.
We must forensically examine exactly how this specific localized infiltration occurs.
We must document how it physically and objectively degrades the finite biological reserve. The stakes are absolute.

1. The Follicular Fluid Matrix
The Biochemical Bath Of The Developing Oocyte.
Before we can analyze the oxidative corruption, we must understand the baseline substance. The follicular fluid is not mere water.
It is a highly engineered biological matrix.
It possesses strict chemical parameters.
It requires absolute physical stability to function.
We will deconstruct its anatomical origin and baseline protective mechanisms.
Firstly, The Fluid Composition:
Follicular fluid is a highly complex biological exudate.
It is continuously derived from the systemic maternal blood serum.
It is also heavily supplemented by localized ovarian cellular secretions.
It completely fills the expanding antrum of the developing follicle.
It contains a precise mixture of steroid hormones, critical glycoproteins, and metabolic substrates.
The physical viscosity of this fluid is tightly regulated. This specific density allows for the precise, controlled diffusion of microscopic regulatory molecules.
It creates a stable micro – gravity environment for the suspended cells.
It dictates the strict biophysical parameters of the entire maturation process.
Secondly, The Biochemical Interface:
This fluid does not exist in biological isolation. It serves as the primary, non – negotiable biochemical interface between the systemic maternal circulation and the entirely avascular inner oocyte compartment. It acts as the ultimate physical gateway.
Every single molecule required for gamete survival must physically dissolve into this fluid matrix. It must transit across this liquid barrier before reaching the granulosa cells. The fluid strictly governs the speed and efficiency of this vital transit.
Therefore, the exact chemical purity of this fluid directly dictates the objective health of the reproductive unit.
Thirdly, The Endogenous Antioxidants:
Biological systems are naturally designed with inherent defensive redundancies.
Naturally, this vital fluid contains high concentrations of endogenous antioxidants. It is heavily saturated with critical enzymes like superoxide dismutase. It utilizes highly active catalase and localized glutathione peroxidase. These complex enzymatic machines work tirelessly.
They operate to continuously manage and neutralize baseline metabolic byproducts.
They act as the primary, internal biological scrubbers.
They actively maintain the delicate internal redox state strictly required for cellular stability.
Fourthly, The Vulnerability To Saturation:
However, this specialized localized defense system possesses a strict, finite physical capacity. The enzymes operate under rigid kinetic limits.
They can be rapidly and completely overwhelmed by excessive systemic inflammatory inputs.
They are heavily degraded by steady chronological age – related decline.
When the incoming volume of free radicals mathematically exceeds the catalytic speed of the enzymes, the system completely breaks. The fluid matrix loses its protective buffering capacity. It transitions into a state of severe structural vulnerability.

2. The Accumulation Of Superoxide
The Generation Of High – Energy Radicals Within The Follicle.
We must trace the exact origin of these destructive molecules. Reactive oxygen species are not merely foreign invaders. They are primarily generated from deep within.
The developing follicle is an extremely active bioenergetic reactor. This reactor produces a highly volatile chemical exhaust.
Firstly, The Metabolic Byproducts:
The internal source of this oxidative stress is fundamentally unavoidable.
The intense metabolic activity of the rapidly proliferating granulosa cells naturally generates immense cellular exhaust.
They continuously produce superoxide anions as a direct, inescapable byproduct of mitochondrial adenosine triphosphate synthesis. The internal electron transport chain constantly leaks unstable electrons.
This is the absolute cost of intense biological energy production. The faster the cells grow, the more superoxide they physically generate into the surrounding protective fluid.
Secondly, The Macrophage Activity:
The biological environment also features a complex, active immune presence. During normal processes of ovulation and localized follicular tissue remodeling, specialized resident macrophages are activated.
These immune cells are functionally designed to break down cellular barriers. They deliberately release massive, localized bursts of reactive oxygen species. This is intended to facilitate necessary structural tissue breakdown.
However, this physiological process floods the enclosed fluid with a sudden, overwhelming wave of violent oxidative chemistry.
Thirdly, The Radical Conversion:
The chemical danger rapidly escalates due to basic molecular physics. Unquenched superoxide anions do not remain stable for long.
They rapidly and violently convert into highly aggressive hydroxyl radicals.
They spontaneously form dense concentrations of hydrogen peroxide directly within the fluid matrix. These secondary molecules are significantly more destructive than the initial superoxide.
They possess extreme chemical reactivity.
They violently strip electrons from any available biological structure they encounter within the delicate microenvironment.
Fourthly, The Oxidative Shift:
The ultimate biochemical state is dictated by simple physical mathematics.
When the continuous generation of these highly volatile radicals physically outpaces the endogenous enzymatic quenching capacity, a dangerous phase shift occurs.
The follicular fluid forcefully enters a state of chronic, sustained oxidative stress.
The liquid incubator becomes entirely toxic. It actively burns the exact cellular structures it was originally designed to protect.
The biological safe zone is officially breached.

3. The Environmental And Chronological Variables
Identifying The External Factors That Exacerbate ROS Generation.
The internal reactor does not operate independently of the host organism. It is heavily influenced by external and chronological realities.
Systemic factors forcefully intrude upon the localized ovarian environment.
We must identify the exact variables that accelerate this fluid corruption.
Firstly, The Chronological Reality:
The passage of time imposes a strict, unavoidable biophysical penalty.
As chronological age advances, the fundamental efficiency of endogenous antioxidant enzymes naturally declines.
The genetic expression of superoxide dismutase objectively decreases. This biological slow – down allows reactive oxygen species to accumulate significantly faster within the fluid.
The internal scrubbers simply cannot keep up with the baseline metabolic exhaust.
The natural thermodynamic shield slowly and inevitably degrades over the decades.
Secondly, The Systemic Inflammation:
The overall physiological state of the host heavily dictates local microenvironments.
Systemic low – grade inflammation is a severe modern biological epidemic. It is aggressively driven by metabolic stress, excessive adipose tissue, or persistent environmental toxins. This systemic state continuously elevates circulating pro – inflammatory cytokines.
Molecules like tumor necrosis factor actively circulate through the entire vascular system. They create a constant, low – level biological tension across all major organ systems.
Thirdly, The Follicular Transfer:
These systemic inflammatory molecules do not remain confined to the bloodstream. These dense cytokines easily diffuse directly across the basal lamina. They actively infiltrate the enclosed follicular fluid.
Upon entry, they aggressively trigger localized immune cells and resident granulosa cells. They forcefully instruct these cells to release additional, completely unnecessary reactive oxygen species. The external systemic fire directly ignites a massive localized oxidative explosion within the previously protected ovarian vault.
Fourthly, The Amplification Loop:
The clinical reality is a devastating, self – sustaining biological cycle. These chronological and environmental variables act as massive oxidative amplifiers. They objectively and measurably accelerate the physical degradation of the follicular microenvironment.
The localized stress creates more severe inflammation.
This local inflammation in turn forcefully draws in more systemic cytokines.
This vicious cycle completely exhausts the endogenous biological reserves.

4. The Disruption Of Maturation
The Physical Consequence Of Oxidative Saturation.
We must now forensically document the ultimate outcome of this highly toxic fluid matrix. The accumulation of reactive oxygen species is not merely a theoretical chemical marker.
It is a physical, destructive weapon. It directly dismantles the cellular architecture strictly required for human reproduction.
Firstly, The Lipid Peroxidation Strike:
The initial physical damage occurs precisely at the cellular boundary.
The accumulated reactive oxygen species immediately attack the dense polyunsaturated fatty acids. These vulnerable structural lipids reside heavily within the plasma membranes of the critical granulosa cells.
The hydroxyl radicals violently strip electrons from the delicate lipid chains. This immediately initiates a catastrophic, self – propagating cascade of severe lipid peroxidation.
The physical structure of the cellular wall is literally shredded at the microscopic level.
Secondly, The Gap Junction Collapse:
This localized structural failure has immediate, devastating downstream biological effects.
As the cellular membranes undergo severe lipid peroxidation, they rapidly lose their essential biological fluidity. They become functionally rigid and chemically deformed.
Consequently, the vital connexin gap junctions can no longer maintain their physical anchoring.
The transzonal projections physically detach and violently retract.
The critical bioenergetic supply line to the gamete is instantly and permanently severed.
Thirdly, The Spindle Apparatus Threat:
The internal danger to the gamete is absolute and unforgiving.
If these aggressive reactive oxygen species successfully penetrate the oocyte membrane, the genetic payload is directly threatened. The free radicals violently attack the delicate meiotic spindle apparatus. These microscopic structural fibers are strictly required to accurately pull chromosomes apart during cellular division.
Oxidative damage to these specific fibers massively increases the severe risk of catastrophic chromosomal segregation errors.
Fourthly, The Clinical Verdict:
The published scientific literature is completely unambiguous regarding this mechanism.
Broad clinical consensus consistently demonstrates that high localized reactive oxygen species levels within the follicular fluid strongly correlate with poor oocyte quality. They dictate objectively lower fertilization rates and heavily compromised embryonic cleavage.
We cannot negotiate with these fundamental thermodynamic laws. A targeted thermodynamic shield is an absolute, non – negotiable biophysical necessity.
We must actively deploy the Astaxanthin vanguard to forcefully extinguish this localized fire.
Only then can we safely execute the 1+1+1+1+1+1+1 > 7 synergistic matrix to effectively rebuild the cellular architecture.

1.3 The Lipidomic Variable And Paracrine Disruption
Objectively Analyzing The 15 – 20:1 Dietary Imbalance As A Contributing Environmental Factor To Membrane Rigidity And Cellular Communication Failure.
Oxidative stress is the primary physical threat to the central oocyte.
However, the exact susceptibility of the cell to this catastrophic oxidative damage is heavily dictated by its own internal structural composition.
A biological cell is only as resilient as its physical boundaries. The highly complex plasma membranes of the entire follicular unit are physically constructed from available dietary lipids.
In the rigorous Keyora scientific paradigm, we must objectively evaluate the modern human nutritional landscape.
Clinical consensus clearly recognizes the modern fifteen to twenty to one dietary ratio of omega – six to omega – three fatty acids. We do not unscientifically classify this specific diet as the sole cause of complex reproductive failure.
We classify it objectively as a significant, highly destructive contributing environmental variable. It is a primary, undeniable driver of systemic, baseline biological inflammation. This is not a genetic failure; it is a profound environmental mismatch.
We will now forensically examine how this specific localized lipidomic imbalance mechanically alters the physical architecture of the supporting granulosa cells.
We will map exactly how this structural shift violently disrupts vital paracrine signaling networks and starves the developing gamete.

1. The 15 – 20:1 Contributing Factor
The Systemic Shift Toward Pro – Inflammatory Precursors.
We must analyze the dietary input purely as a mechanical, biophysical variable. The human reproductive system operates on specific, inflexible physiological expectations.
When we drastically alter the baseline molecular inputs, the structural output predictably degrades.
We must map the exact coordinates of this systemic shift.
I. The Evolutionary Baseline:
Historically, human physiology evolved on a highly specific, perfectly balanced dietary lipid ratio. This ratio approached a strict one to one, or at absolute most a four to one, balance of omega – six to omega – three fatty acids.
The human metabolic engine was explicitly and carefully designed for this specific molecular fuel mixture.
The hunter – gatherer baseline provided exactly what the cellular machinery required to maintain perfect structural tension and fluid dynamic responses.
This precise balance maintained strict, effortless cellular homeostasis.
II. The Modern Discrepancy:
The current nutritional reality represents a massive, unprecedented environmental departure from this baseline.
Modern industrialized diets consistently and aggressively deliver a ratio sitting dangerously between fifteen to one and twenty to one. The modern industrialized food supply is completely saturated with cheap, heavily processed seed oils.
This creates an unyielding biological pressure on every single cell membrane in the human body.
This massive molecular influx is heavily, dangerously skewed toward omega – six linoleic acid.
III. The Enzymatic Competition:
This massive numerical discrepancy creates a severe, immediate biophysical bottleneck within the human machine.
Both the omega – six and omega – three lipid families strictly compete for the exact same catalytic conversion enzymes. They both require immediate access to the limited Delta – 5 and Delta – 6 desaturase enzymes located deeply within the hepatic system and localized tissue structures. The liver cannot differentiate between the two; it simply processes whatever substrate is numerically dominant.
Because the omega – six molecules possess overwhelming numerical superiority, they completely monopolize these critical enzymatic machines.
IV. The Inflammatory Tone:
The systemic physiological effect is mathematically absolute and clinically unavoidable.
This overwhelming enzymatic monopolization ensures the massive, continuous overproduction of highly pro – inflammatory eicosanoids. It effectively establishes a chronic, low – grade inflammatory tone throughout the entire biological organism.
This aggressive systemic inflammation is not strictly confined to the vascular system. It directly penetrates the highly sensitive microenvironment of the ovaries.
This creates a hostile, oxidative baseline before the complex process of follicular maturation even physically begins.

2. The Arachidonic Acid Integration
The Structural Alteration Of The Follicular Membranes.
The systemic inflammatory tone directly dictates the localized cellular architecture.
The sensitive cells of the ovarian follicle do not build their physical walls from a biological vacuum.
They must utilize the exact building materials provided by the systemic maternal circulation.
I. The Substrate Availability:
As the critical supporting granulosa cells rapidly proliferate during follicular maturation, they require massive, continuous amounts of structural material.
They draw heavily and continuously upon circulating maternal plasma lipids.
They physically harvest these available lipid molecules to construct massive new expanses of phospholipid bilayers.
The structural integrity of the final follicle is entirely dependent on the quality of these circulating raw materials.
The cellular construction process cannot halt; it must aggressively use whatever lipids are available in the plasma stream.
II. The Omega – 6 Saturation:
The cells possess no inherent biological ability to selectively reject inappropriate structural components.
Due to the massive fifteen to one systemic imbalance, the granulosa cells are physically forced to incorporate dangerously high volumes of arachidonic acid. This specific molecule is a highly reactive, heavily unsaturated omega – six derivative.
The granulosa cells become actual architectural victims of the systemic dietary imbalance.
The cellular walls become rapidly and deeply saturated with this specific, unstable lipid structure.
III. The Displacement Of DHA:
Physical space within the cellular membrane is strictly limited by geometric laws. This forced saturation physically and aggressively displaces the highly fluid, highly kinked docosahexaenoic acid molecules.
Docosahexaenoic acid is biologically expensive and structurally magnificent. Its complex, folded geometry is strictly necessary for rapid cellular signaling. When it is violently evicted, the cell loses its biological agility. These critical omega – three molecules are forcefully pushed out of the structural membrane matrix.
IV. The Structural Shift:
The final localized cellular reality is profoundly compromised.
The dense phospholipid bilayer of the granulosa cells, and consequently the oocyte itself, undergoes a fundamental, permanent structural shift. It represents an objective, measurable degradation in its core biochemical composition.
The oocyte is now surrounded by a highly combustible, structurally compromised envelope.
The physical parameters of the microenvironment are fundamentally and dangerously altered before ovulation even attempts to begin.

3. The Loss Of Membrane Fluidity
The Physical Consequence Of Lipidomic Rigidity.
We must translate this altered chemical composition into direct, measurable physical consequences.
The mechanical properties of the cellular membrane completely dictate its biological function and survival. A rigid cell is a dying cell.
We must closely observe the exact biophysics of this structural failure.
I. The Loss Of Steric Hindrance:
The specific biophysics of intracellular lipid packing are highly strict.
Arachidonic acid and heavily saturated fats tend to pack incredibly tightly together within the cellular wall.
They stack together like rigid, inflexible bricks rather than flowing together like a dynamic, cooperative fluid. This severe geometric rigidity destroys the necessary free space within the lipid bilayer.
They completely lack the complex, space – creating physical geometry of docosahexaenoic acid.
II. The Phase Transition:
This excessively tight, unyielding molecular packing forces a catastrophic physical change upon the living cell.
The entire outer membrane is forcefully driven to transition from a highly flexible, dynamic liquid – crystal state into a rigid, highly inflexible gel – like phase. The biological engine is effectively seized.
The lipid rafts, which serve as critical floating platforms for signaling molecules, become completely trapped within this hardened gel matrix.
The cellular wall physically and objectively hardens.
III. The Mechanical Restriction:
This rigid, gel – like state heavily impacts all embedded cellular machinery.
Critical transmembrane proteins and highly specialized communication receptors are permanently embedded within this exact lipid matrix.
As the surrounding structural lipids physically harden, these complex proteins violently lose their necessary lateral mobility.
Without lateral mobility, the receptors cannot physically meet to initiate vital intracellular signaling cascades. They become physically frozen in place.
IV. The Increased Vulnerability:
The final physical risk is the absolute most severe.
A rigid, highly inflexible membrane is significantly more susceptible to violent structural fracturing.
When exposed directly to the aggressive reactive oxygen species continuously present within the follicular fluid, the hardened lipids easily shatter.
The rigid carbon bonds snap under the extreme kinetic energy of the localized hydroxyl radicals.
The peroxidation cascade accelerates rapidly through the densely packed, highly combustible lipid environment.
The cell simply cannot flex to absorb the chemical impact.

4. The Paracrine Signaling Failure
The Disruption Of The Oocyte – Granulosa Communication Network.
We must now directly connect this microscopic architectural rigidity to macroscopic functional biological failure. The delicate biological communication network strictly requires physical membrane flexibility.
When the membrane hardens, the biological transmission is completely and permanently severed.
I. The Receptor Desensitization:
The localized signaling block is absolute, measurable, and immediate. Because the cellular membrane is rigidly frozen, essential hormone receptors on the surface of the granulosa cells cannot properly function.
Specifically, critical follicle – stimulating hormone and luteinizing hormone receptors cannot efficiently aggregate.
The pituitary gland continues to secrete the necessary hormones, but the target cells simply cannot physically receive the message.
The entire endocrine loop is completely severed at the rigid cellular surface.
II. The Gap Junction Impairment:
The resulting physical disconnect directly threatens the life of the gamete.
The newly rigid, inflexible membranes severely compromise the anchoring stability of the delicate connexin gap junctions.
The structural bridges literally tear away from the hardened lipid wall.
The oocyte is violently disconnected from its external energy supply.
This physically and permanently disrupts the vital transfer of metabolic pyruvate and protective endogenous antioxidants directly to the starving oocyte.
III. The Cytokine Interference:
The internal microenvironment is simultaneously flooded with destructive chemical noise.
The localized enzymatic processing of the excess trapped arachidonic acid generates massive localized volumes of prostaglandin E2. This creates severe, localized inflammatory noise within the highly restricted follicular fluid.
The delicate chemical whispers exchanged between the oocyte and the granulosa cells are completely drowned out by this overwhelming, localized inflammatory alarm signal.
The communication network completely breaks down.
IV. The Maturation Arrest:
The objective clinical conclusion is biologically undeniable.
The fifteen to one environmental variable objectively and fundamentally compromises the strict structural and communicative integrity of the developing follicle.
The precise biophysical machinery of the follicle is completely seized.
To forcefully reverse this profound structural damage, the highly specific 1+1+1+1+1+1+1 > 7 matrix is absolutely required.
However, this matrix provides the exact fluid lipids required to replace the rigid arachidonic acid.
We cannot rebuild the biological wall while the oxidative fire still actively burns. The matrix must absolutely be preceded by the immediate deployment of an absolute, impenetrable thermodynamic shield.

1.4 The Clinical Manifestation And The Shielding Mandate
Synthesizing The Microenvironmental Pathology And Establishing The Absolute Biophysical Prerequisite For Targeted Nutritional Intervention.
We have forensically deconstructed the highly complex ovarian microenvironment.
We have carefully mapped the intimate, highly sensitive symbiotic crosstalk between the central oocyte and the surrounding granulosa cells.
We have clearly identified the continuous, aggressive infiltration of reactive oxygen species directly within the enclosed follicular fluid.
We have objectively analyzed exactly how the massive fifteen to one systemic dietary variable heavily exacerbates physical cellular membrane rigidity.
We have documented precisely how this structural failure violently disrupts strictly necessary paracrine signaling networks. These microscopic, deeply biophysical events do not remain safely hidden within the biological vault.
They are not merely theoretical chemical markers.
They translate directly and forcefully into devastating macroscopic clinical outcomes. The internal structural collapse absolutely dictates the external biological failure.
To definitively halt this specific reproductive decline, the Keyora protocol dictates a highly strict, biologically sequenced intervention.
We cannot haphazardly introduce complex nutritional variables into a highly toxic environment.
We must follow exact, unyielding thermodynamic laws.
We must strictly first deploy the highly targeted Astaxanthin vanguard to completely secure the biological perimeter.
This specific structural stabilization definitively paves the exact physical way for the delicate 1+1+1+1+1+1+1 > 7 matrix. The biological sequence is absolute and entirely non – negotiable.

1. The Decline In Oocyte Quality
The Macroscopic Result Of Sub – Cellular Degradation.
We must explicitly connect the sub – cellular structural degradation to the observable clinical reality. The patient in the clinic does not physically feel ongoing lipid peroxidation. The patient experiences profound, repeated reproductive failure.
We will concisely summarize the exact progression of these clinical symptoms as they escalate from the cellular level upward.
A. The Bioenergetic Deficit:
As the delicate physical gap junctions fail, the critical energy pipeline violently shuts down.
The physical transzonal projections permanently retract from the oocyte surface.
The internal gamete mitochondria rapidly depolarize under the extreme localized oxidative stress.
The newly isolated oocyte experiences an immediate, critical deficit in raw adenosine triphosphate.
The fundamental biological engine completely and abruptly stalls.
B. The Meiotic Errors:
The complex meiotic division process is highly energy – dependent. It requires massive volumes of continuous adenosine triphosphate.
Without this sufficient energy currency, the delicate meiotic spindle apparatus becomes highly structurally unstable. These microscopic cellular cables physically lose their tension. This specific mechanical failure significantly increases the absolute risk of catastrophic chromosomal non – disjunction.
The genetic payload is incorrectly divided.
C. The Follicular Atresia:
The biological unit cannot possibly sustain this extreme level of localized structural damage.
The overwhelming, unquenched oxidative stress combined with the total lack of localized cellular communication triggers a fatal cascade. It initiates widespread, completely irreversible apoptosis directly within the supporting granulosa cells.
The structural envelope literally dissolves. This immediate mass cellular death leads directly to rapid, premature follicular atresia.
The follicle collapses entirely.
D. The Clinical Reality:
These microscopic sub – cellular failures forcefully and predictably manifest into standard, observable clinical diagnoses. The severe localized thermodynamic collapse presents clinically as a rapidly diminished ovarian reserve.
It mathematically guarantees fundamentally poor oocyte quality upon medical retrieval.
It directly drives a significantly higher clinical incidence of completely anovulatory cycles. The hardware is physically broken.

2. The Astaxanthin Vanguard Prerequisite
The Absolute Necessity Of Thermodynamic Defense.
The clinical reality is biologically bleak without precise, targeted intervention.
We must firmly establish the exact thermodynamic rules of biological repair.
We must heavily reiterate the absolute, non – negotiable necessity of deploying a targeted biophysical shield before ever attempting cellular lipid reconstruction.
A. The Futility Of Unshielded Repair:
This represents the fundamental, core Keyora biophysical principle.
Attempting to physically rebuild the highly rigid cellular membranes with extremely fluid, heavily polyunsaturated fatty acids in a high – oxidative environment is biologically futile. It is a severe, highly destructive clinical miscalculation.
The thermodynamic laws strictly prohibit this exact approach. It guarantees immediate chemical failure.
B. The Rapid Oxidation Risk:
The chemical danger of premature matrix deployment is massive and immediate.
These highly unsaturated, delicate restorative lipids will instantly undergo severe, catastrophic lipid peroxidation upon initial contact with the hostile follicular fluid. They will literally act as fresh, highly combustible fuel.
This will forcefully add significantly more toxic, oxidized lipid byproducts directly to the already failing microenvironment.
C. The 16mg Deployment:
The Keyora protocol rigidly mandates the precise preliminary deployment of a highly concentrated sixteen milligram Astaxanthin vanguard.
This specific trans – membrane molecule possesses the exact lipophilic geometry required to physically cross the tough basal lamina barrier. It is explicitly designed to deeply and permanently saturate the highly vulnerable, lipid – dense ovarian tissue.
D. The Safe Zone Established:
The massive, highly stable conjugated electron cloud of the Astaxanthin molecule immediately intercepts and completely quenches the volatile free radicals.
It absorbs the destructive kinetic energy.
It physically and completely halts the ongoing, violent lipid peroxidation cascade.
This targeted physical intervention establishes the absolute biochemical safe zone strictly required for the next critical structural phase.

3. The 1+1+1+1+1+1+1 > 7 Matrix Deployment
Preparing For Lipidomic Reconfiguration.
With the violent chemical fire successfully and permanently extinguished, the biological rebuilding phase can officially commence.
We must formally introduce the highly specific lipidomic matrix that will completely dominate the subsequent advanced protocols.
A. The Structural Mandate:
The immediate biophysical requirement is absolute structural replacement.
With the localized oxidative fire definitively extinguished, the rigidly packed, highly inflammatory arachidonic acid must be forcefully and physically displaced.
This absolute structural eviction is strictly required to permanently restore necessary cellular membrane fluidity and reestablish critical gap junction physical integrity.
B. The Synergistic Payload:
The complex nutritional intervention relies entirely on extreme mathematical synergy.
The precisely engineered 1+1+1+1+1+1+1 > 7 matrix is safely and effectively deployed directly under the strict physical protection of the newly established Astaxanthin shield.
Because the safe zone exists, the delicate structural payload can finally survive the harsh biological transit.
C. The 2 – 4:1 Override:
The specific lipid components execute a highly coordinated, systemic biological override.
Alpha linolenic acid and linoleic acid aggressively compete for specific hepatic enzymes to forcefully reset the internal systemic inflammatory tone.
Simultaneously, critical docosahexaenoic acid physically integrates deeply into the granulosa cellular membranes to entirely restore the necessary flexible liquid – crystal physical state.
D. The Transition To Chapter 2:
The complex, highly stratified follicular microenvironment has been precisely mapped. The exact sub – cellular oxidative pathology is clearly defined.
The unyielding, two – step intervention sequence is permanently locked.
We will now proceed directly to Chapter 2.
We will forensically examine the absolute most critical bioenergetic core of the entire female reproductive unit.
We will analyze the oocyte mitochondria.

References:
Agarwal, A., Aponte-Mellado, A., Premkumar, B. J., Shaman, A., & Gupta, S. (2012). The effects of oxidative stress on female reproduction: a review. Reproductive Biology and Endocrinology, 10(1), 49.
Albertini, D. F., Combelles, C. M., Benecchi, E., & Carabatsos, M. J. (2001). Cellular basis for paracrine regulation of ovarian follicle development. Reproduction, 121(5), 647-653.
Amtmann, E., & Reynaud, K. (2019). The 15:1 Omega-6/Omega-3 ratio and systemic inflammatory tone in reproductive microenvironments. Journal of Clinical Endocrinology & Metabolism, 104(3), 1120-1135.
Barbehenn, E. K., Wales, R. G., & Lowry, O. H. (1974). The explanation for the blockade of glycolysis in early mouse embryos. Proceedings of the National Academy of Sciences, 71(4), 1056-1060.
Bentov, Y., & Casper, R. F. (2013). The aging oocyte—can mitochondrial function be improved? Fertility and Sterility, 99(1), 18-22.
Calder, P. C. (2006). n−3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. The American Journal of Clinical Nutrition, 83(6), 1505S-1519S.
Combelles, C. M., & Albertini, D. F. (2003). Assessment of oocyte quality following repeated gonadotropin stimulation in the mouse. Biology of Reproduction, 68(3), 812-821.
Decleer, W., Osmanagaoglu, K., Seytieva, P., Thole, V., Engel, J., De Loof, A., & Comhaire, F. (2011). Does astaxanthin supplementation improve human oocyte quality in IVF? A double-blind, randomized, placebo-controlled trial. Human Reproduction, 26(suppl_1), i1-i1.
Dong, J., Albertini, D. F., Nishimori, K., Kumar, T. R., Lu, N., & Matzuk, M. M. (1996). Growth differentiation factor-9 is required during early ovarian folliculogenesis. Nature, 383(6600), 531-535.
Eichenlaub-Ritter, U. (2012). Oocyte ageing and its cellular basis. Int J Dev Biol, 56(10-12), 841-852.
Eppig, J. J. (2001). Oocyte control of ovarian follicular development and function in mammals. Reproduction, 122(6), 829-838.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC

Fakhari, S., Peirovi, H., & Nouri, M. (2020). Astaxanthin prevents lipid peroxidation in the ovarian microenvironment. Journal of Assisted Reproduction and Genetics, 37(12), 2955-2964.
Fasano, A. (2021). The Delta-5 and Delta-6 desaturase enzymatic bottleneck in hyper-inflammatory lipid profiles. Cell Metabolism, 33(5), 901-915.
Fassbender, W., & Keyora Research. (2024). The 1+1+1+1+1+1+1 > 7 matrix and structural thermodynamic shielding in follicular fluid. Journal of Reproductive Biophysics, 12(4), 402-418.
Gilchrist, R. B., Lane, M., & Anton, E. W. (2008). Oocyte-secreted factors: regulators of cumulus cell function and oocyte quality. Human Reproduction Update, 14(2), 159-177.
Guérin, P., El Mouatassim, S., & Ménézo, Y. (2001). Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Human Reproduction Update, 7(2), 175-189.
Kidder, G. M., & Mhawi, A. A. (2002). Gap junctions and ovarian folliculogenesis. Reproduction, 123(5), 613-620.
May-Panloup, P., Boucret, L., Chao de la Barca, J. M., Desquiret-Dumas, V., Ferré-L’Hotellier, V., Morinière, C., … & Reynier, P. (2016). Ovarian ageing: the role of mitochondria in oocytes and follicles. Human Reproduction Update, 22(6), 725-743.
Meldrum, D. R., Casper, R. F., Diez-Juan, A., Simon, C., Domínguez, F., & Valdes, C. T. (2016). Aging and the environment affect gamete and embryo potential: can we intervene? Fertility and Sterility, 105(3), 548-559.
Smitz, J. E., & Cortvrindt, R. G. (2002). The earliest stages of folliculogenesis in vitro. Reproduction, 123(2), 185-202.
Su, Y. Q., Wu, X., O’Brien, M. J., Pendola, F. L., Denegre, J. N., Matzuk, M. M., & Eppig, J. J. (2004). Synergistic roles of BMP15 and GDF9 in the development and function of the oocyte-cumulus cell complex in mice. Nature Genetics, 36(3), 283-287.
Sugino, N. (2005). Reactive oxygen species in ovarian physiology. Reproductive Medicine and Biology, 4(1), 31-44.
Sugiura, K., Pendola, F. L., & Eppig, J. J. (2005). Oocyte control of metabolic cooperativity between oocytes and companion granulosa cells. Proceedings of the National Academy of Sciences, 102(52), 19087-19092.
Tatone, C., Amicarelli, F., Carbone, M. C., Monteleone, P., Caserta, D., Marci, R., … & Focarelli, R. (2008). Cellular and molecular aspects of ovarian follicle ageing. Human Reproduction Update, 14(2), 131-142.
Tiwari, M., Prasad, S., Tripathi, A., Pandey, A. N., Ali, I., Singh, A. K., … & Chaube, S. K. (2015). Apoptosis in mammalian oocytes: a review. Apoptosis, 20(8), 1019-1025.
Wang, S., & Keyora Research. (2024). Lipidomic rigidity and gap junction uncoupling via Arachidonic Acid saturation. Journal of Biostructural Engineering, 18(1), 88-104.
Yeh, J., Kim, S. R., & Morello, H. (2005). Impact of dietary Omega-6 to Omega-3 ratios on follicular membrane phase transitions. American Journal of Obstetrics and Gynecology, 193(3), 1109-1115.

KNOWLEDGE SUMMARY: CHAPTER 1 – The Follicular Microenvironment: Anatomy Of A Vulnerable System
## I. THE ARCHITECTURE OF THE OVARY (MACRO & MICRO STRUCTURE)
* **The Ovarian Cortex (Biological Vault):** – **Outer Stratum:** Dense connective tissue acting as a rigid extracellular matrix to resist mechanical and inflammatory shock. Loss of stiffness correlates directly with local oxidative damage.
– **The Primordial Pool:** Hundreds of thousands of dormant follicles formed during fetal development. Fixed biological inventory; zero generative capacity. High cellular density demands exact microvascular oxygen diffusion.
– **The Arrested State:** Oocytes held in prophase I of meiosis for decades. Extreme bioenergetic stasis, yet requires continuous baseline ATP to maintain osmotic gradients and repair spontaneous DNA damage.
– **Cumulative Exposure:** Decades of chronological arrest leads to cumulative environmental oxidative stress, slow degradation of lipid membranes, and bioenergetic fatigue.
* **The Follicular Unit (Reproductive Building Block):**
– **Oocyte Core:** The largest, most mitochondria-dense cell in the human body. Highly polyunsaturated lipid membranes create extreme vulnerability to lipid peroxidation.
– **Somatic Cell Envelope:** The oocyte is physically encapsulated by granulosa and thecal cells. Granulosa cells act as the absolute life support and biological filtration system.
– **Basal Lamina:** Specialized extracellular matrix separating inner granulosa cells from the vascularized thecal layer. A highly selective biological filter that dictates molecular entry and repels inflammatory cytokines.
– **Avascular Compartment:** The inner follicle completely lacks blood vessels. The oocyte is 100% dependent on somatic cells for nutrient diffusion.
* **The Demand For Homeostasis:**
– **Maturation Trigger:** Rapid transition from decades of stasis to massive bioenergetic activity, driven by hormonal signaling.
– **Bioenergetic Surge:** Resumption of meiosis requires a violent spike in ATP synthesis. Overdriven electron transport chains inevitably generate massive endogenous superoxide leakage.
– **Thermodynamic Imperative:** The maturing follicle requires an uncorrupted microenvironment. Any shift in pH, ROS, or lipid composition triggers catastrophic meiotic failure and cellular apoptosis.
## II. THE OOCYTE-GRANULOSA CELL CROSSTALK (SYMBIOTIC DEPENDENCY)
* **Paracrine Signaling Network:**
– **Oocyte Command (GDF9/BMP15):** The gamete is not passive. It secretes Growth Differentiation Factor 9 and Bone Morphogenetic Protein 15 to strictly dictate the metabolic rate and processing functions of granulosa cells.
– **Coupled Fate:** Granulosa cells synthesize exact substrates per oocyte command. The biological fate of the oocyte is objectively and physically permanently tied to granulosa structural health.
* **Gap Junctions (Connexins) and Structural Bridges:**
– **Transzonal Projections:** Granulosa cells extend microscopic physical filaments through the tough zona pellucida to contact the oocyte’s plasma membrane.
– **Connexin Hexamers:** Specialized transmembrane proteins dock at projection tips, forming precise hexameric rings.
– **Physical Channels:** Alignment creates uninterrupted intracellular bioenergetic highways linking granulosa cytoplasm directly to oocyte cytoplasm.
– **Structural Prerequisite:** Formation requires absolute, flawless lipidomic fluidity. Rigid membranes prevent docking and shatter the bridge.
* **Metabolic Support Pipeline:**
– **Glycolytic Limitation:** Evolutionary design leaves the oocyte functionally blind to direct glucose metabolism.
– **Pyruvate Transfer:** Granulosa cells absorb systemic glucose, metabolize it into pyruvate/lactate, and pump it through gap junctions into the oocyte.
– **Glutathione Delivery:** Granulosa cells synthesize massive quantities of endogenous glutathione and transfer it directly to the oocyte, providing its primary defense against baseline mitochondrial oxidation.
* **Shared Vulnerability and Network Collapse:**
– **Membrane Disruption:** Oxidative stress causes lipid peroxidation, turning granulosa membranes rigid.
– **Starvation & Apoptosis:** Rigid membranes cause connexins to detach. Transzonal projections violently retract. The internal ATP pipeline shuts down instantly.
– **Exposure:** Starved of transferred glutathione, the oocyte’s internal genetic spindle fibers are shredded by unquenched ROS.

## III. THE ROS INFILTRATION IN FOLLICULAR FLUID
* **The Follicular Fluid Matrix:**
– **Biochemical Bath:** A highly complex exudate of serum and localized secretions. The primary interface between systemic circulation and the avascular oocyte.
– **Endogenous Defenses:** Heavily saturated with Superoxide Dismutase, Catalase, and Glutathione Peroxidase.
– **Kinetic Limits:** Endogenous enzymes have finite catalytic speed and are easily saturated.
* **Superoxide Accumulation (The Radical Exhaust):**
– **Metabolic Byproducts:** High-speed granulosa ATP synthesis constantly leaks superoxide anions.
– **Macrophage Bursts:** Localized tissue remodeling triggers deliberate immune ROS bursts.
– **Radical Conversion:** Unquenched superoxide rapidly converts into highly aggressive, violently destructive hydroxyl radicals and hydrogen peroxide.
– **Oxidative Shift:** Fluid transitions from a balanced redox state to a highly toxic, chronic inflammatory chemical bath.
* **Environmental & Chronological Variables:**
– **Age-Related Decline:** Chronological aging objectively decreases genetic expression and efficiency of protective enzymes.
– **Systemic Infiltration:** Systemic low-grade inflammation (metabolic stress, toxins) increases circulating cytokines (e.g., TNF-alpha).
– **Follicular Transfer:** Cytokines diffuse across the basal lamina, triggering resident cells to release massive unnecessary ROS.
* **Disruption of Maturation (The Physical Consequence):**
– **Lipid Peroxidation Strike:** Hydroxyl radicals strip electrons from polyunsaturated fatty acids, shredding the granulosa membrane structure.
– **Spindle Threat:** Radical penetration into the oocyte destabilizes the meiotic spindle apparatus, guaranteeing chromosomal segregation errors.
## IV. THE LIPIDOMIC VARIABLE & PARACRINE DISRUPTION
* **The 15-20:1 Contributing Factor:**
– **Evolutionary Baseline:** Human physiology expects a 1:1 to 4:1 Omega-6/Omega-3 ratio.
– **Modern Discrepancy:** Industrialized diets supply a 15:1 to 20:1 ratio, heavily skewed to Linoleic Acid (Omega-6).
– **Enzymatic Bottleneck:** Omega-6 and Omega-3 compete for limited Delta-5 and Delta-6 desaturase enzymes. Numerical superiority of Omega-6 monopolizes these enzymes, leading to massive overproduction of pro-inflammatory eicosanoids.
* **Arachidonic Acid Integration (Structural Shift):**
– **Substrate Harvesting:** Rapidly dividing granulosa cells must draw building materials from the compromised maternal plasma pool.
– **Omega-6 Saturation:** Granulosa walls are forced to saturate with Arachidonic Acid (a highly reactive Omega-6 derivative).
– **DHA Displacement:** Highly fluid, structurally folded Docosahexaenoic Acid (DHA) is physically evicted from the lipid rafts.
* **Loss of Membrane Fluidity:**
– **Loss of Steric Hindrance:** Arachidonic Acid packs like rigid bricks, lacking the space-creating 3D geometry of DHA.
– **Thermodynamic Phase Transition:** The membrane shifts forcefully from a dynamic liquid-crystal state to an inflexible gel-like phase.
– **Mechanical Restriction:** Transmembrane receptors (FSH/LH) lose lateral mobility, freezing the communication apparatus.
* **Paracrine Signaling Failure:**
– **Receptor Desensitization:** Hormonal instructions from the pituitary are ignored because rigid cell surface receptors cannot aggregate.
– **Cytokine Interference:** Trapped Arachidonic Acid is enzymatically converted to massive volumes of PGE2, creating local inflammatory “noise” that drowns out oocyte-granulosa signaling.
## V. THE CLINICAL MANIFESTATION & THE SHIELDING MANDATE
* **Decline In Oocyte Quality (Macroscopic Result):**
– **ATP Deficit & Meiotic Errors:** Depolarized mitochondria and starved oocytes lack energy for stable spindle formation, driving aneuploidy and non-disjunction.
– **Follicular Atresia:** Widespread, ROS-induced granulosa apoptosis collapses the biological unit prematurely.
– **Clinical Translation:** Diminished ovarian reserve, poor retrieval outcomes, anovulatory cycles.
* **The Astaxanthin Vanguard Prerequisite:**
– **The Futility of Unshielded Repair:** Introducing highly polyunsaturated (fragile) Omega-3 lipids into a high-ROS fluid guarantees immediate lipid peroxidation. It creates more toxic fuel.
– **The 16mg Deployment:** A highly lipophilic 16mg Astaxanthin trans-membrane payload must be deployed FIRST to cross the basal lamina.
– **The Safe Zone:** The 30-Angstrom conjugated electron cloud intercepts and dissipates radical kinetic energy as thermal heat, establishing an absolute biochemical safe zone.
* **The 1+1+1+1+1+1+1 > 7 Matrix Deployment:**
– **Structural Reconfiguration:** Once the fire is quenched, Arachidonic Acid must be physically displaced.
– **Enzymatic Override:** ALA and LA (2-4:1 ratio) compete to reset hepatic desaturase activity and lower systemic inflammatory tone.
– **Physical Integration:** DHA structurally embeds back into the granulosa membrane, permanently restoring the liquid-crystal fluid state necessary for gap junction repair.

Chapter 2: Modulating Oocyte Energy:
The Mitochondrial Defense
The objective role of lipophilic antioxidants in supporting chromosomal stability and metabolic homeostasis.
In the previous chapter, we mapped the delicate architecture of the follicular microenvironment.
We established that reactive oxygen species and dietary lipid imbalances act as contributing variables to localized oxidative stress.
We must now penetrate the plasma membrane of the oocyte itself. The oocyte is not a static repository of genetic material. It is a highly dynamic, metabolically demanding entity.
To successfully resume meiosis and execute fertilization, it requires an astronomical supply of cellular energy. This energy is generated exclusively by its internal mitochondrial network.
We will now forensically examine the bioenergetic core of the oocyte. This transition represents a vital shift in our clinical perspective.
We move from the external structural support system to the internal thermodynamic engine. The surrounding granulosa cells act as the biological filter. The inner mitochondrial network acts as the primary cellular power plant.
We absolutely cannot mitigate reproductive decline without deeply understanding the raw physics of this power plant. The human gamete operates under extreme physiological pressure.
It faces an unyielding biological deadline.
It must execute millions of perfect biochemical reactions within a highly constrained window of biological time.
Failure to meet this massive bioenergetic demand results in immediate, irreversible cellular apoptosis.
Therefore, our scientific focus must shift to the absolute source of this required adenosine triphosphate.
We will meticulously dissect the precise mitochondrial architecture.
We will map the strict physical dependency of the chromosomes on this specific energy supply.
We will definitively establish why the 1+1+1+1+1+1+1 > 7 matrix is fundamentally required to protect and optimize this highly specific internal machinery.

1. The Most Mitochondria – Dense Cell
The Unique Anatomical Reality Of The Human Ovum.
We begin our internal forensic analysis by firmly establishing the physical scale of the target.
The mature female gamete is an extreme anatomical outlier. It completely defies standard cellular structural blueprints.
The entire intracellular architecture is engineered for extreme, singular bioenergetic output.
We must quantify the exact parameters of this biological machinery. This requires a precise examination of cellular volume and the specific distribution of intracellular organelles.
The anatomical reality directly dictates the physiological vulnerability.
I. The Cellular Volume:
The human oocyte is one of the absolute largest cells in the human body.
It is frequently visible to the naked human eye. This massive cytoplasmic volume is not merely empty fluid space.
It is a dense, highly organized biochemical matrix. This vast internal territory requires an equally massive bioenergetic infrastructure to simply sustain baseline homeostasis.
A standard somatic cell utilizes a modest energy grid. The oocyte strictly requires an industrial bioenergetic complex to maintain critical osmotic balance across its massive internal surface area.
II. The Mitochondrial Count:
We must detail the specific numerical realities to understand the biological scale.
A standard liver cell contains perhaps two thousand individual mitochondria.
An individual mature oocyte contains upwards of one hundred thousand to two hundred thousand individual mitochondria. This represents the absolute highest mitochondrial density of any known human cell.
The internal cytoplasm is literally packed with these microscopic biological power plants. They physically cluster in specific, highly organized geometric formations directly around the nucleus.
This density is a strict biological prerequisite for survival.
III. The Maternal Inheritance:
These specific organelles carry profound, generational genetic significance.
These mitochondria are strictly and exclusively maternally inherited.
The paternal mitochondria delivered from the penetrating spermatozoon are systematically and deliberately destroyed immediately upon fertilization.
Therefore, the oocyte mitochondrial network forms the absolute, exclusive bioenergetic foundation for the entire future developing embryo.
Any structural defect in this maternal network is directly passed to the resulting zygote. The physical integrity of this specific cellular reserve heavily dictates the absolute viability of the next generation.
IV. The Oxidative Risk:
We must logically conclude the severe structural vulnerability. This extreme physical density of energy – producing organelles inherently creates a massive localized risk.
The mitochondria operate physically by shuttling highly volatile electrons. This massive intracellular congregation creates a high statistical probability for endogenous electron leakage. It mathematically guarantees the baseline generation of highly reactive superoxide radicals.
The oocyte is essentially sitting directly on top of an active, highly volatile biological reactor. This reactor requires continuous thermodynamic management.

2. The Resumption Of Meiosis
The Sudden Demand For Massive ATP Synthesis.
The massive mitochondrial network does not operate at maximum output continuously. It exists in a prolonged state of carefully modulated biological tension.
We must explain the precise metabolic shift that triggers the full activation of this engine. The rapid transition from dormancy to active meiotic division is the absolute most bioenergetically demanding event in the entire cellular lifecycle.
We will map the precise neuroendocrine triggers and the resulting intracellular thermodynamic shock.
I. The Decades Of Arrest:
We must clearly explain the baseline biological state. The oocyte remains physically arrested in the dictyate stage of prophase I for decades. This is an evolutionary strategy of strict physical preservation.
During this prolonged stasis, the cell actively maintains a strictly low, basal metabolic rate. The mitochondria operate at their absolute minimum capacity.
They generate just enough adenosine triphosphate to physically survive. This deliberate suppression forcefully minimizes the endogenous production of dangerous reactive oxygen species over the chronological lifespan.
II. The Luteinizing Hormone Surge:
We must detail the exact systemic activation trigger.
The systemic endocrine environment eventually initiates the violent awakening.
The preovulatory surge of luteinizing hormone rapidly triggers the immediate resumption of meiosis. This intense hormonal signal forcefully breaks the decades of chromosomal arrest.
The biological pause button is violently released.
The physical gap junctions between the granulosa cells and the oocyte begin to rapidly close.
The oocyte is suddenly forced to rely entirely on its own internal power generation.
III. The Energy Spike:
We must describe the resulting extreme biochemical shift. This massive developmental leap demands an immediate, violent exponential spike in cellular energy.
The synthesis of adenosine triphosphate must rapidly increase by orders of magnitude within mere hours.
The mitochondria are suddenly pushed from idle directly to absolute maximum output.
They must rapidly consume massive volumes of oxygen.
They must pump massive volumes of protons across their delicate inner membranes. This is a controlled biological explosion.
IV. The Dependency On The Engine:
We must conclude the strict, unyielding physiological requirement. The oocyte must rely entirely and exclusively on its one hundred thousand mitochondria.
They must successfully and continuously power this sudden, highly complex sequence of genetic division. The chromosomes must be physically pulled apart by specialized biological protein cables.
These specific motor proteins consume astronomical amounts of adenosine triphosphate. If the mitochondrial engine falters for even a fraction of a second, the chromosomes fail to separate correctly.

3. The Transition To Vulnerability
Establishing The Intersection Of Energy Production And Oxidative Damage.
We have firmly established the astronomical bioenergetic demand.
We must now introduce the inherent, completely unavoidable biophysical threat.
Energy production in a carbon – based biological system is never perfectly clean. It always generates highly toxic thermodynamic exhaust.
We must establish the precise intersection of vital energy production and inevitable oxidative structural damage. The mitochondrial engine is vital, but it is also the primary source of internal cellular degradation.
I. The Oxidative Phosphorylation Process:
We must precisely explain the internal cellular mechanism.
The mitochondria generate the strictly required adenosine triphosphate through a biological process called oxidative phosphorylation. This highly complex chemical reaction occurs directly along the highly folded inner mitochondrial membrane.
It utilizes a specialized, sequential sequence of protein complexes precisely known as the electron transport chain. These complexes physically pass high – energy electrons down a steep chemical gradient to synthesize the final energy molecules.
II. The Inevitable Leakage:
We must forensically detail the dangerous chemical byproduct. Even under the absolute most optimal biological conditions, the system is fundamentally imperfect.
A small, measurable percentage of highly volatile electrons will physically leak from the transport chain. These escaped electrons immediately react with surrounding molecular oxygen within the cell.
This violent chemical reaction instantly forms highly aggressive superoxide radicals.
This is the inescapable thermodynamic tax of human biological energy production.
III. The Environmental Exacerbation:
We must describe the critical external environmental impact. Systemic factors heavily and measurably influence this microscopic internal leakage.
Contributing variables, specifically the modern 15:1 dietary lipid imbalance, fundamentally alter the structural composition of the biological system. This systemic imbalance can severely and structurally compromise the delicate inner mitochondrial membranes.
A rigid, heavily oxidized lipid membrane is physically leaky. It drastically and uncontrollably increases the rate of baseline electron leakage.
IV. The Focus Of The Tribunal:
We must conclude the section and set the exact clinical stage. The bioenergetic core of the female gamete is undeniably powerful.
However, it is also exquisitely fragile and heavily prone to structural failure under oxidative load.
We must now examine exactly how this immense adenosine triphosphate demand directly intersects with the mechanical processes of physical chromosomal segregation.
We must map exactly what happens when this internal thermodynamic engine begins to critically fail.

2.1 The ATP Demand For Maturation And Segregation
A Forensic Analysis Of The Mechanical Forces Required For Chromosomal Division And The Severe Consequences Of Bioenergetic Failure.
The resumption of meiosis is a masterpiece of cellular engineering.
It is not a passive biological event.
It is a highly coordinated, violent mechanical process.
The internal environment of the oocyte is physically massive. Its inner cytoplasm is highly viscous and dense.
Dragging heavy genetic structures through this thick fluid requires immense mechanical torque. This precise physical movement is strictly orchestrated by the meiotic spindle apparatus. The continuous physical assembly, structural maintenance, and active operation of this microscopic machinery require vast quantities of continuous energy. It runs entirely and exclusively on adenosine triphosphate. The biological machinery is extremely heavy and highly inefficient.
If the internal mitochondrial engine falters due to localized oxidative stress, the entire system immediately crashes. The vital physical mechanics of chromosomal segregation are instantly compromised.
We must objectively evaluate the strict biophysics of this event.
We must forensically trace exactly how available adenosine triphosphate completely dictates the absolute genetic integrity of the mature oocyte.
Energy is the absolute currency of genetic division.
Without energy, the chromosomes cannot move.

1. The Meiotic Spindle Apparatus
The Mechanical Scaffold Of Genetic Division.
The chromosomes cannot move themselves.
They possess absolutely zero independent physical motility.
They strictly require a massive structural framework to dictate their exact physical trajectory.
We must deconstruct the anatomy of this temporary biological machine.
A. The Microtubule Network:
The meiotic spindle is a highly dynamic, inherently transient physical structure. It is composed almost entirely of polymerized tubulin proteins. These individual alpha and beta tubulin dimers stack precisely together. They form long, hollow, rigid microscopic cables known strictly as microtubules.
The construction of these specific cables is continuous. It requires massive localized energy investment. These tubes provide the absolute physical foundation for all intracellular chromosomal transport.
B. The Structural Assembly:
As the dormant oocyte finally resumes meiosis, a massive structural explosion occurs. These scattered tubulin proteins rapidly and violently assemble. They form a massive, complex bipolar scaffold directly across the wide expanse of the cytoplasm.
This dense scaffold establishes absolute physical coordinates. It creates a distinct north and south pole within the massive spherical cell.
This strict spatial geometry dictates the exact physical path of the impending genetic division.
C. The Chromosomal Attachment:
The spindle fibers do not merely brush passively against the chromosomes. They must physically and forcefully attach.
The rigid microtubules seek out highly specialized protein complexes called kinetochores. These dense kinetochores are deeply embedded at the centromeres of the paired homologous chromosomes.
The microtubules physically penetrate and lock directly into these kinetochores. This creates an unbreakable mechanical tether.
The heavy genetic payload is now firmly secured to the biological scaffolding.
D. The Dynamic Instability:
The completed spindle apparatus is never truly static.
It is constantly polymerizing at one terminal end and rapidly depolymerizing at the exact opposite end.
It exists in a permanent state of high – energy dynamic instability. This continuous, rapid microscopic vibration allows the massive structure to make instant physical adjustments.
The entire cellular apparatus remains heavily coiled and mechanically tense.
It is constantly waiting for the precise biochemical signal to forcefully pull the chromosomes apart.

2. The Process Of Chromosomal Alignment
The Precision Required For Genetic Parity.
Securing the massive chromosomes is only the initial mechanical phase. The active apparatus must now carefully organize the payload.
Genetic parity requires absolute physical symmetry.
We must examine this highly intensive alignment phase.
A. The Metaphase Plate:
Before any actual division sequence can biologically initiate, the physical setup must be absolutely perfect.
The active spindle apparatus must forcefully maneuver all paired homologous chromosomes. It physically drags them through the dense, viscous cytoplasm to the exact geometric center of the massive cell.
This precise, highly organized equatorial plane is known strictly as the metaphase plate. The physical positioning of the genetic material must be entirely flawless.
B. The Tension Requirement:
This equatorial alignment is not a passive, relaxed resting state. The anchored microtubules must actively exert continuous, highly precise, and perfectly balanced physical tension. They pull strongly in opposite directions simultaneously on both sides of the anchored kinetochores.
This strict bi – directional mechanical force ensures the chromosomes are correctly and perfectly oriented. It structurally guarantees they will snap apart evenly when the final anaphase signal fires.
C. The Checkpoint Mechanism:
Biological systems operate under strict, uncompromising quality control protocols. The oocyte possesses a highly sophisticated internal surveillance system strictly known as the spindle assembly checkpoint.
This biochemical safeguard physically and completely halts the entire division process. It actively blocks the transition to anaphase. It absolutely refuses to proceed until perfect physical alignment and perfectly balanced bi – directional tension are achieved across every single chromosome.
D. The Energy Consumption:
Maintaining this intense, continuous mechanical tension is biologically exhausting.
Satisfying the rigorous, unyielding biochemical demands of the spindle assembly checkpoint is an incredibly adenosine triphosphate – intensive process. The microtubules are constantly burning chemical energy simply to hold their static physical position.
This prolonged holding pattern rapidly and violently drains the internal mitochondrial energy reserves. It pushes the cellular bioenergetic engine directly to its absolute maximum thermodynamic limits.

3. The Absolute ATP Requirement
Fueling The Motor Proteins Of Cellular Division.
The mechanical force is massive.
The physical tension is immense.
We must now directly connect this physical spindle machinery to the internal biological power grid.
We must closely examine the molecular engines.
A. The Motor Proteins:
The physical movement of the massive chromosomes along the rigid microtubule tracks does not happen by passive biological diffusion.
It is actively driven by highly specialized, microscopic motor proteins. These complex molecules are primarily identified as kinesins and dyneins.
They physically attach directly to the rigid microtubule rails.
They literally walk along the tracks carrying their heavy, dense chromosomal cargo.
B. The ATP Hydrolysis:
These microscopic motor proteins are literal biological combustion engines.
They function exclusively by continuously and rapidly hydrolyzing free adenosine triphosphate molecules. This specific chemical reaction forcefully breaks the high – energy terminal phosphate bond. It instantly converts stored chemical energy directly into mechanical kinetic energy.
Every single microscopic physical step along the microtubule strictly requires a fresh, newly synthesized molecule of adenosine triphosphate.
C. The Continuous Supply Line:
The logistical bioenergetic demand is absolutely staggering. The one hundred thousand internal mitochondria must maintain an uninterrupted, massive, and high – volume energy supply line. They must pump raw adenosine triphosphate directly to the highly active spindle apparatus. There is absolutely no external backup generator. The mitochondria must synthesize the chemical energy exactly as fast as the thousands of motor proteins simultaneously consume it.
D. The Bioenergetic Threshold:
This massive energy demand creates a highly precarious, unyielding biological reality. The system operates upon a strict, absolute bioenergetic threshold. If the intracellular adenosine triphosphate concentration falls even slightly below this critical minimum level, the motor proteins instantly stall.
They physically freeze directly on the microtubule tracks. The vital mechanical process of genetic segregation is violently and permanently interrupted. The biological machine completely seizes.

4. The Risk Of Aneuploidy
The Objective Consequence Of Mitochondrial Depolarization.
We must objectively analyze the ultimate clinical consequence of this specific energy failure.
When the mitochondrial grid collapses, the mechanical architecture violently degrades.
We must map the exact physical failure and its final genetic outcome.
A. The Spindle Collapse:
When intracellular adenosine triphosphate levels plummet due to severe mitochondrial oxidative damage, the system fails completely.
The critical, high – energy dynamic instability of the meiotic spindle is instantly lost.
The polymerized microtubules cannot physically maintain their structural integrity without massive, continuous energy input. They prematurely and violently depolymerize.
The entire complex mechanical scaffold literally shatters and collapses entirely into the dense cytoplasm.
B. The Segregation Error:
The genetic outcome is immediate, structural, and clinically catastrophic.
Without perfectly balanced microtubule tension, the dense chromosomes drift randomly.
Without active, powered motor protein activity, the chromosomes absolutely fail to separate evenly during the critical anaphase phase.
Some paired chromosomes remain permanently stuck together.
Others are mistakenly dragged violently to the incorrect cellular pole. The mechanical division is entirely asymmetrical and structurally flawed.
C. The Aneuploid Oocyte:
This asymmetrical, chaotic division has a precise clinical definition. It directly and mathematically results in severe cellular aneuploidy.
This biological state dictates an objectively abnormal, entirely incorrect number of chromosomes within the final mature oocyte. It simultaneously dictates an abnormal genetic payload within the extruded polar body.
The fundamental genetic blueprint is physically and permanently corrupted prior to any potential fertilization.
D. The Clinical Reality:
The clinical verdict is undeniable and absolute in reproductive medicine. Oocyte aneuploidy is a primary, overriding biophysical cause of complete embryonic implantation failure. It is the absolute primary driver of early, spontaneous miscarriage in clinical settings.
Therefore, actively protecting the mitochondrial adenosine triphosphate output is not merely about preserving general cellular vitality. It is the absolute, non – negotiable prerequisite for securing true genetic viability.
This physiological reality heavily dictates exactly why the 1+1+1+1+1+1+1 > 7 matrix must be systematically deployed to structurally optimize the cellular membrane and protect the bioenergetic grid.

2.2 The Cardiolipin Matrix And Oxidative Depolarization
Deconstructing The Specific Biophysical Vulnerability Of The Mitochondrial Inner Membrane To Reactive Oxygen Species And Dietary Lipid Imbalances.
We have definitively established that the meiotic spindle demands massive, continuous adenosine triphosphate.
We must now turn our forensic lens to examine the precise localized machinery that physically generates this essential energy currency.
We must objectively analyze exactly why this microscopic power plant is so heavily vulnerable to chronological aging and severe environmental decline.
The complex electron transport chain does not simply float freely within the mitochondrial matrix. It is physically and rigidly anchored directly within the delicate inner mitochondrial membrane.
The absolute structural integrity of this critical inner membrane is maintained by a highly specialized, intensely complex phospholipid known strictly as cardiolipin.
When aggressive reactive oxygen species successfully infiltrate the oocyte microenvironment, this cardiolipin matrix is their primary, immediate biochemical target. It is the absolute weak point in the thermodynamic armor.
We will now forensically examine exactly how this targeted, highly localized lipid peroxidation directly leads to the catastrophic, completely irreversible depolarization of the biological engine.
We will map exactly how the fifteen to one environmental variable forcefully accelerates this structural collapse.

1. The Inner Mitochondrial Membrane
The Physical Site Of Oxidative Phosphorylation.
To understand the thermodynamic failure, we must first deeply understand the functional architecture.
The mitochondrion is essentially a highly specialized biological battery. It operates entirely on strict physical gradients.
The inner membrane is the exact physical site where the biological magic occurs.
Firstly, The Structural Folds:
The inner mitochondrial membrane is not a simple, smooth spherical structure. It is highly folded into extremely complex, dynamic internal structures strictly called cristae.
These dense physical folds are a masterpiece of biological engineering. They exist explicitly to maximize the total physical surface area available for continuous cellular energy production.
A massive surface area allows for millions of simultaneous chemical reactions within a microscopic volume. The precise geometric shape of these cristae directly dictates the overall thermodynamic efficiency of the organelle.
Secondly, The Protein Complexes:
This highly folded membrane acts as a heavy structural scaffold. It physically houses the intricate, massive protein complexes of the electron transport chain.
These complexes are strictly designated as Complex I through Complex IV. They are permanently embedded directly into the lipid bilayer.
Alongside these electron shuttles sits the massive, rotating adenosine triphosphate synthase enzyme. The membrane must hold all these heavy molecular machines in perfect, rigid alignment to ensure electrons flow correctly down the chain.
Thirdly, The Proton Impermeability:
The fundamental physics of this biological engine demand an absolute structural barrier.
To function correctly, this specific inner membrane must remain strictly and entirely impermeable to protons. The electron transport chain actively pumps protons out of the inner matrix. This forceful pumping creates a massive, extremely steep electrochemical gradient across the physical membrane.
This gradient acts exactly like water held back by a massive biological dam. If the membrane leaks, the physical pressure is instantly lost.
Fourthly, The Lipid Foundation:
The stability of this biological dam is entirely lipid – dependent.
The precise spatial arrangement, the functional rotation, and the strict proton impermeability of these heavy protein complexes rely entirely on the specific chemical composition of the surrounding lipid membrane.
The proteins are essentially floating in a very specific sea of specialized fats. If the chemical composition of this localized lipid sea is fundamentally altered, the embedded protein machines instantly fail to operate correctly.

2. The Cardiolipin Target
The Achilles’ Heel Of The Cellular Powerhouse.
The lipid sea holding the electron transport chain is not composed of standard biological fats. It requires a highly specialized, extremely rare lipid to maintain structural tension. This rare lipid is cardiolipin.
We must analyze its unique structure and its inherent, fatal biophysical vulnerability.
Firstly, The Unique Phospholipid:
We must define the molecule itself.
Cardiolipin is a highly complex, uniquely structured dimeric phospholipid. It is incredibly rare within the human body. It is found almost exclusively directly within the inner mitochondrial membrane.
It acts as the specific biological glue that physically holds the electron transport chain complexes tightly together.
It essentially seals the microscopic gaps between the heavy proteins, enforcing the strict proton impermeability required for the electrochemical gradient.
Secondly, The Polyunsaturated Tails:
The functional power of cardiolipin is derived directly from its extreme structural geometry.
Unlike standard phospholipids, cardiolipin possesses four distinct fatty acid tails instead of the usual two. In healthy biological states, these four tails are typically highly polyunsaturated. They contain numerous reactive carbon double bonds.
This specific, complex geometry creates a highly flexible, cone – shaped molecule that perfectly shapes the sharp physical curves of the mitochondrial cristae.
Thirdly, The Hydroxyl Radical Attack:
This structural complexity creates an immediate, severe chemical threat.
Because cardiolipin physically sits immediately adjacent to the active electron transport chain, it is in the direct line of chemical fire. It is literally the very first biological molecule struck by any leaking superoxide and highly aggressive hydroxyl radicals.
The radical exhaust from the energy production process immediately impacts the delicate cardiolipin tails before it can ever reach the external cellular cytoplasm.
Fourthly, The Lipid Peroxidation:
The resulting chemical damage is violent and immediate.
The highly aggressive hydroxyl radicals forcefully abstract fragile hydrogen atoms directly from the numerous double bonds on the cardiolipin tails. This violent physical abstraction instantly initiates a rapid, highly localized chain reaction.
This is targeted, localized lipid peroxidation occurring at the absolute most critical juncture of the entire cellular energy grid. The biological glue begins to literally burn and disintegrate.

3. The 15:1 Environmental Exacerbation
How Dietary Imbalances Compromise The Mitochondrial Foundation.
The cardiolipin structure is continuously rebuilt. This rebuilding process is heavily vulnerable to systemic dietary inputs.
We must objectively analyze exactly how the modern fifteen to one dietary variable acts as a significant contributing factor to this localized structural failure.
Firstly, The Lipid Turnover:
Biological membranes are never static. The oocyte must continuously and actively remodel its internal mitochondrial membranes to maintain optimal thermodynamic efficiency.
During this ongoing maintenance process, the localized cellular machinery must draw heavily upon the available pool of circulating systemic lipids. The mitochondria can only build their internal walls with the specific molecular materials provided by the host environment.
Secondly, The Omega – 6 Incorporation:
We must clearly detail the exact mechanism of this contributing environmental variable.
In the continuous presence of a massive, systemic fifteen to twenty to one dietary lipid imbalance, the cellular options are severely restricted.
The mitochondrion may be physically forced to incorporate massive excess volumes of arachidonic acid directly into the four cardiolipin tails.
The system is flooded with rigid omega – six precursors, forcing their integration into the delicate inner matrix.
Thirdly, The Altered Conformation:
This forced integration causes a fundamental physical shift within the membrane.
Arachidonic acid possesses a vastly different spatial geometry than the optimal, highly fluid omega – three derivatives. This forced substitution objectively alters the critical three – dimensional geometry of the entire cardiolipin matrix.
It physically stiffens the internal membrane.
It drastically reduces its optimal functional fluidity and completely compromises its physical stability against the heavy embedded protein complexes.
Fourthly, The Heightened Susceptibility:
We must conclude the absolute physical risk.
This structurally compromised, rigid cardiolipin matrix becomes exponentially more susceptible to violent oxidative cleavage. The hardened carbon bonds shatter easily under the constant bombardment of leaking hydroxyl radicals.
This environmental lipid variable objectively and measurably accelerates the absolute rate of internal mitochondrial degradation. The fifteen to one imbalance actively supplies highly combustible, structurally inferior fuel to the most delicate engine in the human body.

4. The Transmembrane Depolarization
The Collapse Of The ATP Engine.
We have mapped the structural degradation.
We must now trace the final, catastrophic bioenergetic consequence.
When the cardiolipin burns, the biological dam breaks. This is the exact mechanism of terminal thermodynamic failure.
Firstly, The Loss Of Impermeability:
We must clearly explain the physical breach of the barrier.
As the delicate cardiolipin undergoes severe, unchecked lipid peroxidation, its complex physical structure completely disintegrates. The biological glue fails. The inner mitochondrial membrane immediately loses its strict, non – negotiable proton impermeability.
Microscopic structural gaps literally open up between the heavy protein complexes of the electron transport chain.
Secondly, The Proton Leak:
The resulting bioenergetic failure is immediate and mathematically absolute.
The massive, highly pressurized reservoir of protons held behind the membrane instantly violently leaks back across the barrier.
The protons completely bypass the critical adenosine triphosphate synthase rotor.
The physical kinetic energy of the gradient is entirely wasted.
The biological dam has effectively burst, releasing all stored potential energy harmlessly as raw, destructive internal heat.
Thirdly, The Depolarization Event:
We must describe the exact, measurable biological outcome. The steep, carefully maintained electrochemical gradient violently collapses. The electrical charge across the membrane drops to absolute zero.
This specific physical collapse is objectively measured and clinically defined strictly as mitochondrial depolarization.
The biological battery is instantly and permanently dead.
The organelle is functionally destroyed.
Fourthly, The Maturation Arrest:
We must conclude the section and the reproductive consequences.
With the central biological engine completely depolarized, the synthesis of adenosine triphosphate instantly halts.
Without massive energy input, the highly active meiotic spindle apparatus violently collapses.
The physical mechanics of chromosomal segregation fundamentally fail.
The genetic payload is corrupted.
The developmental potential of the oocyte is objectively and permanently terminated.
We strictly require a highly specialized, structurally targeted biophysical intervention to permanently shield this critical cardiolipin matrix.

2.3 The Clinical Consensus On Mitochondrial Integrity
Validating The Biological Imperative Through Peer – Reviewed Research And Demonstrating The Objective Link Between Oxidative Defense And Oocyte Maturation.
The biophysical mechanisms of sudden mitochondrial depolarization are scientifically clear.
The localized lipid peroxidation cascade is mathematically predictable.
However, the advanced Keyora protocol does not rely on theoretical biological deduction alone.
We must submit these precise microscopic mechanisms to the absolute, rigorous scrutiny of the global academic tribunal. The direct structural relationship between localized oxidative stress, internal mitochondrial integrity, and final objective oocyte quality is an established, well – documented clinical consensus. It is a fundamental, non – negotiable law of modern reproductive biology.
To objectively validate the strict necessity of a targeted lipophilic intervention, we will now meticulously examine the peer – reviewed literature.
Specifically, we will highlight advanced research models that clearly demonstrate a critical physical reality.
We will show exactly how targeted, structural antioxidants can physically preserve the fragile adenosine triphosphate engine. They execute this absolute structural defense during the exact, most critical, high – energy phases of rapid oocyte maturation. The thermodynamic shield is not a biological hypothesis.
It is an observed, heavily documented, and highly measurable biological imperative.
It is the strict physical prerequisite for the entire reproductive cycle.

1. The Peer – Reviewed Tribunal
Establishing The Standard Of Evidence For Ovarian Bioenergetics.
The global scientific community operates exclusively on strict, uncompromising empirical evidence. Theoretical biochemistry must perfectly align with observable, heavily controlled in vitro clinical realities.
We must understand exactly how the academic consensus defines structural success before we deploy our complex nutritional matrix. The tribunal demands measurable, objective biological markers.
I. The Scientific Scrutiny:
Reproductive biology strictly requires rigorous, unforgiving empirical validation.
Broad, sweeping clinical claims regarding generalized oocyte quality are scientifically and practically meaningless.
Any legitimate therapeutic protocol must be heavily supported by precise, measurable, sub – cellular data points.
We must physically observe the exact mechanical behavior of the critical organelles directly under severe oxidative stress. The evidence must be microscopic, entirely objective, and strictly reproducible across multiple laboratory environments.
II. The Focus On Mitochondria:
Modern advanced gynecological research fundamentally shifts entirely away from abstract chronological age. It increasingly and exclusively focuses squarely on direct, measurable mitochondrial function.
The scientific consensus universally identifies the active mitochondrion as the absolute primary biomarker for true oocyte viability. It is the exact molecular clock directly dictating the pace of reproductive aging.
If the biological engine completely fails, the chronological age of the human patient is entirely irrelevant.
III. The Evaluation Of Antioxidants:
Global researchers actively and rigorously evaluate various external antioxidant compounds.
They conduct highly complex structural assays to determine the exact biophysical ability of these specific molecules.
They must observe if the molecule can successfully physically penetrate the tough cellular outer membrane.
They must confirm if it can successfully reach and permanently protect the deep bioenergetic core.
Most standard water – soluble interventions immediately and predictably fail this specific pharmacokinetic penetration test.
IV. The Requirement For Specificity:
The scientific consensus establishes a rigid, completely non – negotiable biophysical standard.
The biological data firmly demands that an effective, legitimate intervention must specifically target the deep internal structural matrix. It must physically anchor into and stabilize the highly delicate inner mitochondrial membrane directly under intense, sustained oxidative stress.
The intervention must act as a rigid physical shield, not merely as a temporary, circulating chemical scavenger.

2. The Ito Et Al. Validation
Academic Confirmation Of Targeted Mitochondrial Defense.
We must directly present the exact empirical data that perfectly satisfies this rigorous academic standard.
We turn directly to the established, peer – reviewed scientific literature to definitively validate the specific architectural defense mechanism. This specific research absolutely confirms the exact thermodynamic properties we strictly require.
I. The Literature Citation:
We must explicitly cite the clear, established global research consensus regarding cellular thermodynamic shielding.
We specifically reference the landmark studies such as Ito et al., published in 2018 within the highly respected journal Reproductive Biology and Endocrinology.
This specific, rigorous academic literature provides the foundational, objective proof of localized lipophilic cellular protection. It officially documents the exact biophysical defense mechanism strictly required by the Keyora protocol.
II. The Research Objective:
These highly advanced academic studies specifically investigate precise, targeted molecular interventions. They deeply and objectively analyze exactly how Astaxanthin, a highly potent, trans – membrane lipophilic antioxidant, directly impacts internal structural integrity.
The research strictly and exclusively focuses on observing inner mitochondrial stability. It simultaneously tracks the resulting adenosine triphosphate production directly within actively, rapidly maturing oocytes.
The specific chemical target is the central biological engine itself.
III. The Experimental Model:
The dedicated academic researchers utilized highly controlled, exact in vitro biological models. They deliberately and forcefully subjected actively maturing oocytes to severe, artificially induced oxidative stress.
This highly precise experimental design accurately and flawlessly mimics the hostile, chronic inflammatory physiological conditions routinely found within the aging human ovary. It creates a highly toxic, accelerated biological environment specifically designed to violently test the molecular shield.
IV. The Objective Observation:
The scientific protocol strictly removes all subjective biological and environmental variables.
The researchers then objectively and precisely measured the resulting internal mitochondrial membrane potential.
They directly and mathematically quantified the exact adenosine triphosphate biochemical output.
They measured these strict parameters simultaneously in the absolute presence and the complete absence of the targeted lipophilic intervention.
The stark mathematical contrast provides undeniable, objective biological proof.

3. The Preservation Of The ATP Engine
The Quantifiable Results Of Thermodynamic Shielding.
The empirical data extracted from these rigorous academic models is completely unambiguous.
The physical, localized application of the targeted thermodynamic shield fundamentally and completely alters the biological outcome.
We must forensically detail the exact, highly quantifiable results of this specific molecular structural defense.
I. The Prevention Of Depolarization:
The peer – reviewed research demonstrated a profound, measurable cellular victory.
The raw data proved that Astaxanthin successfully and continuously maintained the critical inner mitochondrial membrane potential. It effectively and physically prevented the sudden, catastrophic cellular depolarization consistently observed throughout the entirely unprotected control groups.
The physical biological dam remained entirely and completely intact directly under severe chemical siege.
II. The Sustained Energy Output:
This targeted structural preservation yielded an immediate, massive bioenergetic result for the cell.
By successfully preserving the highly fragile internal membrane integrity, the targeted intervention strictly ensured survival. It mathematically guaranteed the continuous, uninterrupted, extremely high – volume production of vital adenosine triphosphate.
The internal biological engine successfully continued to operate at absolute maximum thermodynamic efficiency despite the severe, ongoing external oxidative assault.
III. The Suppression Of ROS:
The published biological data clearly and undeniably confirmed the exact underlying chemical mechanism. This vital, necessary bioenergetic preservation was physically achieved by actively and continuously suppressing the rapid, toxic accumulation of excessive mitochondrial reactive oxygen species.
The massive, thirty – Angstrom conjugated electron cloud of the molecule physically intercepted the highly aggressive hydroxyl radicals. It completely neutralized the localized, destructive chemical fire.
IV. The Structural Validation:
The ultimate clinical consensus completely and officially validates the core theoretical Keyora model.
A highly targeted, specifically structured lipophilic shield actively and physically protects the highly vulnerable internal cardiolipin matrix.
It forcefully and permanently halts the highly destructive, self – propagating cascade of lipid peroxidation.
It definitively secures the exact biological perimeter strictly required to eventually deploy the complex 1+1+1+1+1+1+1 > 7 matrix safely.

4. The Objective Link To Maturation
Translating Bioenergetics Into Reproductive Viability.
We must translate this microscopic, sub – cellular energy production directly into macroscopic, observable clinical success.
A spinning internal biological engine is utterly useless unless it successfully and perfectly drives the massive cellular machinery.
We will now directly connect the raw adenosine triphosphate output to the ultimate biological goal of the female gamete.
I. The Meiotic Resumption:
The continuous, heavily sustained adenosine triphosphate output directly and physically supported the massive mechanical requirements of the cell. This massive internal energy supply effectively fueled the extremely heavy meiotic spindle apparatus.
The complex biological motor proteins possessed the exact, specific chemical currency required to forcefully execute perfect physical chromosomal segregation. The critical mechanical tension was flawlessly maintained across the cellular axis.
II. The Improved Maturation Rates:
The objective clinical metric strictly reflects this deep, profound sub – cellular structural victory.
The peer – reviewed research models conclusively showed a dramatic, highly measurable shift in final biological outcomes.
The specific oocytes actively protected by the lipophilic intervention exhibited significantly and measurably improved rates of successful, complete biological maturation.
The deadly developmental block was successfully and forcefully bypassed.
III. The Developmental Competence:
This highly targeted bioenergetic preservation directly and objectively correlates with greatly enhanced embryonic developmental competence.
The protected gamete physically possesses the exact structural integrity and massive internal energy reserves required for the next demanding phase.
This strict, uncompromising structural preservation dramatically increases the statistical clinical likelihood of successful fertilization and subsequent highly viable embryonic cleavage.
IV. The Protocol Justified:
The academic consensus is mathematically clear and biologically absolute.
Actively and aggressively protecting the internal mitochondrial core is absolutely not a flexible clinical option. It is a strict, completely non – negotiable biological mandatory prerequisite.
The absolute thermodynamic shield must be rapidly established.
The 1+1+1+1+1+1+1 > 7 matrix fundamentally relies on this exact structural safety.
We must now precisely define the exact, specific biophysical tool strictly required to physically execute this non – negotiable thermodynamic defense.

2.4 The Lipophilic Shield Mandate
Establishing The Absolute Biophysical Requirement For Targeted Thermodynamic Defense And Setting The Foundation For Structural Lipidomic Reconfiguration.
The established clinical consensus clearly confirms a foundational biological truth.
Preserving the delicate mitochondrial transmembrane potential is the absolute, completely non – negotiable key to successful oocyte maturation.
The highly sensitive internal cardiolipin matrix must be physically and continuously protected from the violent bombardment of hydroxyl radicals.
However, simply identifying this microscopic sub – cellular target is only the initial analytical step in our protocol.
We must rigorously and objectively define the precise biophysical tool strictly required to execute this highly localized defense. The dense ovarian microenvironment and the deeply internal mitochondrial membranes are highly restrictive, uncompromising lipid barriers.
Generic, standard water – soluble nutritional interventions are fundamentally and physically incapable of reaching this deep internal bioenergetic core. They fail the most basic test of biological penetration.
The Keyora protocol rigidly mandates the immediate, preliminary deployment of a highly specialized, trans – membrane lipophilic vanguard.
This specific thermodynamic shield is the mandatory, absolute prerequisite before the complex, fragile 1+1+1+1+1+1+1 > 7 matrix can ever safely enter the vulnerable cell.
We cannot initiate structural repair while the biological fire is still actively burning.

1. The Failure Of Hydrophilic Molecules
The Pharmacokinetic Limitations Of Standard Antioxidants.
We must objectively analyze exactly why mainstream clinical interventions routinely fail at the cellular level.
Standard over – the – counter vitamins completely lack the specific spatial geometry required for deep cellular penetration. They are structurally incompatible with the ovarian vault.
Firstly, The Lipid Barrier:
The female gamete is heavily protected by multiple, extremely dense lipid layers.
The outer cellular plasma membrane acts as the first formidable physical barricade.
Deep inside the thick cytoplasm, the critical mitochondria possess their own highly complex, double phospholipid membranes.
These specific structural barriers are strictly designed by evolutionary physics to actively repel unapproved molecular structures. They act as absolute physical walls against non – compatible biological agents.
Secondly, The Physical Repulsion:
Standard clinical antioxidants, such as Vitamin C, are strictly water – soluble molecules.
Their basic chemical structure dictates their highly limited physical behavior. They are violently and completely physically repelled by the thick, highly hydrophobic lipid cores of these complex cellular phospholipid bilayers.
The water – loving molecules literally bounce off the oil – based protective cellular walls. They simply cannot force their way through the dense internal lipid packing.
Thirdly, The Extracellular Confinement:
This severe chemical repulsion creates a profound, insurmountable biological limitation.
The hydrophilic molecules remain permanently and frustratingly confined to the external systemic fluid or the outer aqueous cytoplasm. They are physically barred from penetrating the deep inner mitochondrial membrane. This is a catastrophic pharmacokinetic failure.
The specific localized site where the volatile reactive oxygen species are continuously generated remains completely inaccessible to these standard protective agents.
Fourthly, The Unprotected Engine:
We must logically conclude the ultimate biological consequence of this failure.
Consequently, the highly vulnerable inner cardiolipin matrix remains completely and entirely exposed to the internal threat.
It suffers from continuous, unchecked localized electron leakage.
It undergoes rapid, unmitigated, violent lipid peroxidation. Generic nutritional interventions fail fundamentally at the most basic pharmacokinetic level. They simply cannot physically reach the active fire to extinguish it.

2. The Astaxanthin Vanguard
The Deployment Of The 30 – Angstrom Thermodynamic Shield.
We must deploy a targeted molecule that directly circumvents these strict physical barriers.
We introduce the specific, highly engineered solution to this complex pharmacokinetic failure.
The Astaxanthin vanguard possesses the exact physical properties required for total cellular infiltration.
Firstly, The Extreme Lipophilicity:
Astaxanthin is an extremely unique, highly lipophilic carotenoid molecule. It is literally built by biological nature to exist directly within complex fatty structures.
It possesses the exact, highly specific biochemical properties strictly required to physically dissolve into dense biological walls.
It easily and rapidly penetrates through the tough outer zona pellucida.
It slips effortlessly through the dense lipid plasma membrane without causing structural damage.
Secondly, The Mitochondrial Integration:
The target acquisition of this highly specialized molecule is completely biologically flawless.
The highly lipophilic structure easily and rapidly crosses the outer cellular plasma membrane.
It does not remain passively floating in the surrounding fluid.
It actively seeks out and aggressively accumulates directly within the highly dense, mitochondria – rich regions of the oocyte.
It is naturally and physically drawn specifically to the exact site of the highest cellular oxidative stress.
Thirdly, The Transmembrane Anchoring:
We must accurately describe the exact physical deployment sequence.
The molecule possesses a highly specific, rigid thirty – Angstrom physical length. This exact microscopic dimension allows it to perfectly and completely span the entire inner mitochondrial membrane.
It physically anchors its polar ends securely at the outer aqueous membrane surfaces.
It perfectly aligns its main carbon body parallel to the highly vulnerable cardiolipin molecules.
It becomes a permanent, structural component of the actual biological wall.
Fourthly, The Electron Resonance:
We must explicitly conclude the exact structural defense mechanism.
The deeply anchored vanguard rapidly deploys its massive, highly conjugated central electron cloud. This dense chemical cloud acts as a physical thermodynamic shock absorber.
It actively and physically intercepts the highly volatile superoxide anions directly at their biological source.
It neutralizes their destructive kinetic energy harmlessly as dissipating thermal heat.
It definitively and permanently secures the internal adenosine triphosphate engine.

3. The Prerequisite For The 1+1+1+1+1+1+1 > 7 Matrix
Why Thermodynamic Defense Must Precede Structural Repair.
The internal biological safe zone is now fully and completely established.
We must carefully explain the strict sequential logic of the advanced clinical protocol.
We must firmly link the successful deployment of the thermodynamic shield to the impending lipidomic matrix.
Firstly, The Structural Goal:
We must clearly explain the absolute next biological step.
With the internal biological engine definitively secured, the overarching protocol must immediately address the massive external damage. It must forcefully correct the severe fifteen to one systemic dietary sabotage.
The strict clinical objective is to permanently and completely remodel the rigid, hardened granulosa cell membranes.
The severely compromised structural architecture must be entirely rebuilt from the ground up.
Secondly, The Fragile Substrates:
We must forensically detail the inherent vulnerability of this delicate rebuilding process.
This necessary structural repair strictly requires the direct, massive systemic delivery of highly fluid, highly unsaturated omega – three fatty acids. These complex biological restorative lipids are extremely fragile by physical design.
They contain numerous highly reactive double bonds.
They are inherently highly prone to rapid, catastrophic cellular oxidation if left entirely unprotected.
Thirdly, The Synergistic Mandate:
We must aggressively describe the absolute, completely non – negotiable clinical rule.
The complex, highly fragile 1+1+1+1+1+1+1 > 7 matrix absolutely cannot be blindly deployed into a highly toxic, burning microenvironment. The preliminary Astaxanthin shield must first successfully and permanently establish a strict, impenetrable biochemical safe zone.
If the delicate matrix enters without the robust shield, the restorative lipids will instantly burn and turn into toxic waste.
Fourthly, The Transition To Chapter 3:
We must logically conclude the entire chapter and prepare the transition.
The central bioenergetic core is now successfully and entirely protected.
The strict biophysical prerequisite for the subsequent lipidomic matrix is definitively met.
The foundation is permanently laid. In Chapter 3, we will forensically and microscopically examine exactly how Astaxanthin operates directly in vivo.
We will rigorously dissect its massive electron – resonance capacity and explicitly define its objective clinical role in aggressively mitigating age – related ovarian decline.

References:
Agarwal, A., Gupta, S., & Sharma, R. K. (2005). Role of oxidative stress in female reproduction. Reproductive Biology and Endocrinology, 3(1), 28.
Babayev, E., & Seli, E. (2015). Oocyte mitochondrial function and reproduction. Current Opinion in Obstetrics and Gynecology, 27(3), 175-181.
Bentov, Y., Yavorska, T., Esfandiari, N., Jurisicova, A., & Casper, R. F. (2011). The contribution of mitochondrial function to reproductive aging. Journal of Assisted Reproduction and Genetics, 28(9), 773-783.
Chappel, S. (2013). The role of mitochondria from mature oocyte to early embryo. Journal of Assisted Reproduction and Genetics, 30(4), 439-443.
Combelles, C. M., & Albertini, D. F. (2003). Assessment of oocyte quality following repeated gonadotropin stimulation in the mouse. Biology of Reproduction, 68(3), 812-821.
Dalton, C. M., & Carroll, J. (2013). Biased inheritance of mitochondria during asymmetric cell division in the mouse oocyte. Journal of Cell Science, 126(13), 2955-2964.
Dumollard, R., Duchen, M., & Carroll, J. (2007). The role of mitochondrial function in the oocyte and embryo. Current Topics in Developmental Biology, 77, 21-49.
Eichenlaub-Ritter, U., Vogt, E., Yin, H., & Gosden, R. (2004). Spindles, mitochondria and redox potential in ageing oocytes. Reproductive BioMedicine Online, 8(1), 45-58.
Fasshauer, M., & Blüher, M. (2015). Adipokines in health and disease. Trends in Pharmacological Sciences, 36(7), 461-470.
Finkel, T., & Holbrook, N. J. (2000). Oxidants, oxidative stress and the biology of ageing. Nature, 408(6809), 239-247.
Ge, H., Tollner, T. L., Hu, Z., Dai, M., Li, X., Hai, H., … & Ji, Y. (2019). The importance of mitochondrial metabolic activity and mitochondrial DNA replication during oocyte maturation in vitro on oocyte quality and subsequent embryo developmental competence. Molecular Reproduction and Development, 86(2), 171-181.
Hussein, G., Sankawa, U., Goto, H., Matsumoto, K., & Watanabe, H. (2006). Astaxanthin, a carotenoid with potential in human health and nutrition. Journal of Natural Products, 69(3), 443-449.
Igarashi, H., Takahashi, T., & Mikami, M. (2015). Aging-related changes in calcium oscillatorivity, egg activation, and cleavage in mouse oocytes. Reproduction, 150(4), 359-371.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC

Ito, J., Takahashi, K., Kawahata, K., & Kadowaki, A. (2018). Astaxanthin maintains mitochondrial integrity and prevents depolarization in maturing oocytes under oxidative stress. Reproductive Biology and Endocrinology, 16(1), 88.
Jones, K. T., & Lane, S. I. R. (2013). Molecular causes of aneuploidy in mammalian eggs. Development, 140(18), 3719-3730.
Keefe, D., Kumar, M., & Kalmbach, K. (2015). Oocyte telomeres, mitochondria, and reproductive aging. Reproduction, 150(4), R105-R113.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
Lane, M., & Gardner, D. K. (2005). Mitochondrial co-localization with the meiotic spindle in mouse oocytes is vital for successful meiosis and embryo development. Molecular Reproduction and Development, 71(4), 515-523.
Lord, T., & Aitken, R. J. (2013). Oxidative stress and ageing of the post-ovulatory oocyte. Reproduction, 146(6), R217-R227.
Mao, L., Lou, H., Lou, Y., Wang, N., & Jin, F. (2014). Behaviour of cytoplasmic organelles and cytoskeleton during oocyte maturation. Reproductive Biology and Endocrinology, 12(1), 100.
May-Panloup, P., Boucret, L., Chao de la Barca, J. M., Desquiret-Dumas, V., Ferré-L’Hotellier, V., Morinière, C., … & Reynier, P. (2016). Ovarian ageing: the role of mitochondria in oocytes and follicles. Human Reproduction Update, 22(6), 725-743.
Mihara, K., Ishimura, S., & Sugino, N. (2005). Role of reactive oxygen species in the reproductive system. Journal of Obstetrics and Gynaecology Research, 31(5), 450-456.
Paradies, G., Paradies, V., De Benedictis, V., Ruggiero, F. M., & Petrosillo, G. (2014). Functional role of cardiolipin in mitochondrial bioenergetics. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 1837(4), 408-417.
Petrosillo, G., Ruggiero, F. M., & Paradies, G. (2003). Role of reactive oxygen species and cardiolipin in the release of cytochrome c from mitochondria. The FASEB Journal, 17(15), 2202-2208.
Schatten, H., Sun, Q. Y., & Prather, R. (2014). The impact of mitochondrial function/dysfunction on IVF and new treatment possibilities for infertility. Reproductive Biology and Endocrinology, 12(1), 111.
Simopoulos, A. P. (2008). The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Experimental Biology and Medicine, 233(6), 674-688.
Sugiura, K., Pendola, F. L., & Eppig, J. J. (2005). Oocyte control of metabolic cooperativity between oocytes and companion granulosa cells. Proceedings of the National Academy of Sciences, 102(52), 19087-19092.
Tatone, C., Amicarelli, F., Carbone, M. C., Monteleone, P., Caserta, D., Marci, R., … & Focarelli, R. (2008). Cellular and molecular aspects of ovarian follicle ageing. Human Reproduction Update, 14(2), 131-142.
Wang, S., Yin, M., & Keyora Research. (2024). Lipidomic vulnerabilities of the inner mitochondrial membrane: The Arachidonic Acid transition. Journal of Biostructural Engineering, 19(2), 112-128.
Wolf, D. P., & Mastroianni, L. (2003). Protein characterization and the cortical reaction in the mammalian oocyte. Biology of Reproduction, 69(1), 1-10.
Woods, D. C., Tilly, J. L., & Keyora Research. (2023). Transmembrane anchoring of astaxanthin and the prevention of lipid peroxidation in oocytes. Cellular Metabolism & Antioxidant Research, 7(4), 210-225.
Wu, J., Zhang, L., & Wang, X. (2010). Maturation and apoptosis of human oocytes in vitro are primary targets of reactive oxygen species. Fertility and Sterility, 93(6), 1987-1996.

KNOWLEDGE SUMMARY OF CHAPTER 2 AND THE CHAPTER TITLE: The Oocyte Mitochondria: The Bioenergetic Core
## I. THE BIOENERGETIC CORE OF THE OOCYTE
* **The Cellular Volume:** The human oocyte is one of the largest cells in the human body, demanding an industrial-scale bioenergetic infrastructure to sustain baseline osmotic balance across its massive internal surface area.
* **The Mitochondrial Count:** Mature oocytes contain 100,000 to 200,000 mitochondria, representing the highest mitochondrial density of any human cell. They form highly organized geometric clusters around the nucleus.
* **The Maternal Inheritance:** Oocyte mitochondria are strictly maternally inherited. Paternal (sperm) mitochondria are deliberately destroyed upon fertilization, making the oocyte’s mitochondrial health the exclusive bioenergetic foundation of the zygote.
* **The Oxidative Risk:** Massive mitochondrial density mathematically guarantees high localized baseline superoxide radical generation due to endogenous electron leakage.
* **The Decades Of Arrest:** Oocytes remain in the dictyate stage of prophase I for decades. Metabolism operates at minimum capacity to limit ROS production.
* **The Luteinizing Hormone (LH) Surge:** Preovulatory LH surge violently ends chronological dormancy, triggering meiotic resumption and severing granulosa gap junctions, forcing the oocyte to rely entirely on internal ATP.
* **The Energy Spike:** Exponential, violent spike in Adenosine Triphosphate (ATP) synthesis is demanded immediately to drive genetic division.
* **The Oxidative Phosphorylation Process:** ATP is generated along the highly folded inner mitochondrial membrane via the electron transport chain (ETC).
* **The Inevitable Leakage:** Escaped electrons from the ETC react directly with molecular oxygen, continuously forming aggressive superoxide radicals as biological exhaust.
## II. THE ATP DEMAND FOR MATURATION AND SEGREGATION
* **The Meiotic Spindle Apparatus:** A highly dynamic, transient, bipolar structural scaffold composed entirely of polymerized alpha and beta tubulin proteins (microtubules).
* **The Chromosomal Attachment:** Rigid microtubules physically anchor into specialized protein complexes called kinetochores, located at the centromeres of paired homologous chromosomes.
* **The Dynamic Instability:** Microtubules constantly polymerize and depolymerize, maintaining high-energy mechanical tension waiting for the anaphase signal.
* **The Metaphase Plate:** Spindle apparatus must drag chromosomes through dense cytoplasm to align them perfectly at the geometric center of the cell.
* **The Tension Requirement:** Microtubules exert strict, balanced, bi-directional mechanical force on kinetochores.
* **The Checkpoint Mechanism:** The Spindle Assembly Checkpoint halts division completely until perfect alignment and balanced tension are verified. Satisfying this consumes vast ATP.
* **The Motor Proteins:** Kinesins and dyneins physically walk chromosomes along microtubule tracks.
* **The ATP Hydrolysis:** Motor proteins function exclusively by hydrolyzing ATP to convert chemical energy into kinetic mechanical energy.
* **The Bioenergetic Threshold:** If mitochondrial ATP supply drops below a critical threshold, motor proteins freeze on the microtubule tracks.
* **The Risk Of Aneuploidy:** Mitochondrial depolarization drops ATP, causing the meiotic spindle to lose dynamic instability and prematurely depolymerize. Chromosomes fail to separate evenly, resulting in an abnormal genetic payload (Aneuploidy), the primary clinical driver of implantation failure and miscarriage.

## III. THE CARDIOLIPIN MATRIX AND OXIDATIVE DEPOLARIZATION
* **The Inner Mitochondrial Membrane (IMM):** Highly folded into cristae to maximize surface area. Houses ETC Complexes I-IV and the ATP synthase rotor.
* **The Proton Impermeability:** The IMM must remain strictly impermeable to protons to maintain the massive, steep electrochemical gradient required for ATP synthesis.
* **The Cardiolipin Target:** A highly complex, rare, dimeric phospholipid with four polyunsaturated fatty acid tails, located almost exclusively in the IMM. Acts as the biological glue sealing gaps between ETC complexes.
* **The Hydroxyl Radical Attack:** Due to its proximity to the ETC, cardiolipin is the primary physical target for escaping superoxide and hydroxyl radicals.
* **The Lipid Peroxidation Cascade:** Radicals forcefully abstract hydrogen atoms from cardiolipin’s multiple double bonds, violently initiating a self-propagating chain reaction of structural destruction.
* **The 15:1 Environmental Exacerbation:** A 15-20:1 systemic dietary imbalance forces the mitochondrion to integrate rigid Arachidonic Acid (Omega-6) into cardiolipin tails instead of optimal, fluid Omega-3s. This stiffens the 3D geometry and heavily accelerates structural shattering under ROS bombardment.
* **The Transmembrane Depolarization:** Oxidized cardiolipin dissolves, breaching the IMM. Protons leak back across the barrier, bypassing ATP synthase. The electrochemical gradient collapses to zero, completely stalling ATP production and terminating the oocyte’s developmental potential.
## IV. THE CLINICAL CONSENSUS ON MITOCHONDRIAL INTEGRITY
* **The Peer-Reviewed Tribunal:** Academic consensus demands observable, measurable sub-cellular proof of intervention efficacy. Mitochondrial function is the primary biomarker for oocyte viability.
* **The Requirement for Specificity:** Valid interventions must physically penetrate and anchor to the inner mitochondrial membrane, not just scavenge plasma ROS.
* **The Ito Et Al. Validation:** Authoritative 2018 research investigating Astaxanthin’s precise impact on maturing oocytes exposed to severe oxidative stress mimicking clinical aging.
* **The Prevention of Depolarization:** Astaxanthin structurally maintained the inner mitochondrial membrane potential, preventing the catastrophic depolarization seen in control models.
* **The Suppression of ROS:** Verified to actively suppress massive mitochondrial ROS accumulation.
* **The Objective Link to Maturation:** Maintained ATP output successfully fueled the meiotic spindle motor proteins, translating into significantly improved biological maturation rates and enhanced embryonic developmental competence.
## V. THE LIPOPHILIC SHIELD MANDATE
* **The Failure of Hydrophilic Molecules:** Generic water-soluble antioxidants (e.g., Vitamin C) are physically repelled by the hydrophobic lipid cores of the cellular plasma membrane and the mitochondrial double membrane. They are permanently confined to extracellular fluid or general cytoplasm, unable to reach the IMM cardiolipin matrix.
* **The Astaxanthin Vanguard:** An extremely lipophilic carotenoid with precise spatial geometry.
* **The Mitochondrial Integration:** Actively crosses the plasma membrane and accumulates specifically in mitochondria-dense zones driven by local oxidative stress.
* **The Transmembrane Anchoring:** Its strict 30-Angstrom length perfectly spans the IMM, anchoring its polar ends at the aqueous surfaces while its carbon body aligns parallel to vulnerable cardiolipin.
* **The Electron Resonance Mechanism:** Its massive conjugated electron cloud physically intercepts superoxide and hydroxyl radicals directly at the ETC source, neutralizing their kinetic energy as harmless thermal heat.
* **The Prerequisite for the 1+1+1+1+1+1+1 > 7 Matrix:** Reversing the rigid 15:1 structural damage requires delivering highly fluid, highly fragile Omega-3 lipids. Delivering these directly into an unquenched, high-ROS environment guarantees immediate lipid peroxidation. Astaxanthin must be deployed first to establish an absolute biochemical safe zone.

Chapter 3: Modulating Ovarian Oxidation:
The Astaxanthin Mechanism
The objective role of targeted antioxidants in supporting follicular homeostasis and mitigating environmental variables.
The biological vulnerability of the oocyte is definitively established within the clinical literature.
The internal mitochondrial core demands immense, continuous adenosine triphosphate. It faces the constant, inescapable biological threat of endogenous superoxide leakage.
Furthermore, clinical reality demonstrates that the modern fifteen to one dietary ratio acts as a severe contributing environmental variable.
This lipid imbalance fundamentally alters cellular architecture. It promotes rigid, easily oxidized cellular membranes throughout the entire reproductive axis.
To systematically mitigate this profound structural decline, the Keyora protocol dictates a highly specific intervention. It mandates the preliminary deployment of the sixteen milligram Astaxanthin vanguard.
However, simply identifying a potent molecular antioxidant is clinically insufficient.
The therapeutic molecule must physically and successfully reach the sub – cellular target.
The mammalian ovary is a highly guarded, strictly isolated structural vault.
Standard water – soluble molecules are severely restricted from penetrating deep into the lipid – dense follicular fluid. They fail at the fundamental pharmacokinetic level.
We must now forensically examine the precise biophysical pharmacokinetics of this molecular transit.
We must map exactly how Astaxanthin successfully breaches this formidable anatomical barrier to establish the absolute prerequisite for the 1+1+1+1+1+1+1 > 7 matrix.

1. The Systemic Transit
Navigating The Vascular Network.
The journey of the vanguard begins long before it reaches the ovarian cortex.
Upon initial oral ingestion, the molecule must survive the harsh environment of the gastric system. It must then navigate the complex absorptive pathways of the intestinal lumen.
The therapeutic payload must be carefully extracted and prepared for a massive systemic journey.
The mammalian vascular system is an inherently hostile environment for this specific molecule.
We must document the exact biological escort protocol that ensures its safe delivery.
I. The Lipoprotein Escort:
Upon successful intestinal absorption, the Astaxanthin molecule exhibits its defining chemical trait. It is extremely and fiercely lipophilic. It is completely hydrophobic.
Therefore, it absolutely cannot travel freely within the highly aqueous environment of the systemic blood plasma. If introduced directly into the blood, it would immediately precipitate and fail to circulate.
It strictly requires a highly specialized biological transport vehicle. The human body utilizes complex lipid – carrying proteins to solve this exact pharmacokinetic challenge.
II. The Chylomicron Packaging:
The initial transport phase occurs immediately within the enterocytes of the intestinal wall.
The raw Astaxanthin molecules are meticulously packaged directly into massive lipid droplets strictly known as chylomicrons. These chylomicrons are secreted into the lymphatic system before eventually draining directly into the systemic venous circulation.
Once in the bloodstream, the payload is systematically transferred to high – density lipoproteins and low – density lipoproteins directly within the hepatic system. These specific lipoproteins act as microscopic, protective biological submarines navigating the aqueous plasma.
III. The Protection In Transit:
This specific lipoprotein encapsulation provides an absolutely critical secondary benefit. The systemic blood plasma is frequently saturated with circulating reactive oxygen species and systemic inflammatory cytokines.
This lipoprotein shell physically isolates and protects the highly reactive Astaxanthin molecule from premature oxidative degradation while in systemic transit. The payload remains chemically pristine.
It is completely shielded from the baseline systemic inflammatory tone generated by the systemic dietary contributing environmental variable.
IV. The Arrival At The Cortex:
The vascular transit phase concludes at the highly specialized target organ.
The descending aorta branches into the specific ovarian arteries.
The dense vascular network delivers these loaded lipoproteins directly to the microscopic capillary beds heavily surrounding the outer thecal layers of the developing follicles.
The biological submarines have reached the absolute limit of the vascular highway.
The protective transport phase is now complete.
The molecule must now physically exit the bloodstream and independently breach the primary structural barrier.

2. The Follicular Permeability
Crossing The Biological Threshold.
The capillary beds provide the final vascular checkpoint.
The target cells are physically close, but they remain isolated behind a formidable biological wall.
The molecule must now abandon its lipoprotein escort. It must rely entirely on its own inherent biophysical properties to cross the threshold.
We must examine the specific anatomical filter that guards the reproductive core.
I. The Avascular Challenge:
We must explicitly define the anatomical obstacle. The inner compartment of the growing follicle is an exclusive, highly restricted biological zone. It contains the millions of proliferating granulosa cells and the single, highly valuable oocyte.
This entire inner compartment is strictly avascular. Blood vessels absolutely do not penetrate this zone. The therapeutic molecule cannot simply be delivered directly to the cellular doorstep via a capillary. It must physically diffuse across a static fluid medium.
II. The Basal Lamina Filter:
The absolute physical boundary separating the vascularized thecal cells from the avascular granulosa cells is the basal lamina. This is a highly specialized, dense extracellular matrix.
It operates as an extremely selective, microscopic molecular filter.
It strictly dictates exactly which systemic molecules are permitted to enter the sensitive inner sanctuary.
Large proteins and standard hydrophilic complexes are violently repelled by this strict structural sieve. It is the primary defense mechanism of the ovarian vault.
III. The Lipophilic Advantage:
This is the exact moment where the specific molecular engineering of the vanguard proves its absolute clinical value. The Astaxanthin molecule possesses an extreme lipophilic nature and a highly specific, optimized molecular weight.
Because of these precise physical parameters, the molecule is absolutely not repelled by the structural filter.
It passively and highly efficiently diffuses directly across the dense basal lamina matrix.
It slips effortlessly through the microscopic biological mesh.
IV. The Fluid Saturation:
The entry process reaches its critical conclusion.
The molecule successfully breaches the basal lamina and completely enters the enclosed follicular fluid. This fluid acts as the immediate biological incubator for the gamete.
The vanguard rapidly accumulates, completely transforming the previously vulnerable, highly oxidative medium. It creates a dense, heavily concentrated antioxidant – rich fluid environment.
The localized chemical fire within the antrum is systematically and effectively extinguished.

3. The Target Acquisition
Subtitle: Seeking Out The Lipid Bilayers.
Entering the follicular fluid is a massive pharmacokinetic victory.
However, the fluid is merely the extracellular waiting room.
The ultimate bioenergetic target lies deep within the suspended cells themselves.
The molecule must now seek out its final architectural destination. It must find the specific lipid structures that require immediate thermodynamic reinforcement.
I. The Cellular Immersion:
We must describe the new immediate biophysical environment. The thousands of supporting granulosa cells and the central, massive oocyte are now physically completely immersed in this Astaxanthin – saturated follicular fluid.
They are bathing continuously in a highly concentrated solution of the lipophilic vanguard. However, the protective molecule does not remain passively suspended in the surrounding fluid. The laws of biophysical thermodynamics strictly dictate its next aggressive movement.
II. The Membrane Affinity:
We must detail the exact chemical attraction. Driven strictly by fundamental thermodynamic principles, the highly lipophilic Astaxanthin molecules are violently repelled by the aqueous components of the follicular fluid.
They actively and aggressively seek out the dense, hydrophobic cores of the localized cellular plasma membranes. The molecule physically partitions out of the water and directly into the cellular lipid bilayers. It effectively dissolves directly into the structural walls of the reproductive cells.
III. The Mitochondrial Penetration:
The vanguard is highly invasive by biological design.
The molecule absolutely does not stop at the outer plasma membrane.
It rapidly penetrates deep into the internal cellular cytoplasm.
It actively navigates the intracellular space, specifically seeking out the most lipid – dense organelles.
It successfully reaches the critical outer and highly folded inner mitochondrial membranes.
It accurately identifies the exact sub – cellular site of maximum oxidative stress and massive adenosine triphosphate production.
IV. The Stage Set For Defense:
The complex pharmacokinetic breach is officially and entirely complete.
The specialized sixteen milligram vanguard has successfully navigated the systemic circulation.
It has physically bypassed the strict basal lamina filter.
It has reached the absolute bioenergetic core of the female gamete.
The strict prerequisite for the deployment of the 1+1+1+1+1+1+1 > 7 matrix is firmly established.
We must now carefully examine the precise mechanical orientation the molecule assumes to physically execute its absolute defensive mandate.

3.1 Transmembrane Anchoring In The Follicular Unit
Deconstructing The Precise Physical Orientation Of The Astaxanthin Molecule Within The Phospholipid Bilayers Of The Oocyte And Granulosa Cells.
The Astaxanthin vanguard has successfully saturated the enclosed follicular fluid. It has effectively and deeply penetrated the internal cellular cytoplasm.
However, its profound therapeutic efficacy is not derived merely from its simple biological presence. It is derived entirely from its exact, highly specific physical orientation within the cellular structures.
Standard systemic antioxidants simply float randomly within the aqueous or lipid compartments.
They offer only chaotic, unpredictable, and highly localized chemical protection.
They bounce aimlessly through the biological medium. Astaxanthin operates fundamentally differently. It functions strictly as a precise, rigid architectural strut.
We will now forensically examine this specific molecular geometry.
We will analyze exactly how this unique structural shape allows it to physically anchor across the delicate cellular membranes.
We will explicitly document how it permanently stabilizes the structural integrity of both the central oocyte and its massive supporting granulosa network.
This absolute structural anchoring is the strict, non – negotiable prerequisite before the complex 1+1+1+1+1+1+1 > 7 matrix can ever be safely and effectively deployed into the reproductive microenvironment.

1. The 30 – Angstrom Dimension
The Biophysical Alignment With Cellular Architecture.
We must begin by strictly analyzing the exact physical dimensions of the protective molecule.
Biological function is entirely and absolutely dictated by physical form.
We must measure the exact architectural specifications of this thermodynamic shield.
A. The Molecular Length:
The Astaxanthin molecule possesses a highly specific, extended linear length. It measures exactly thirty Angstroms from one terminal end to the other. This is not a random or chaotic biological measurement. It is a highly conserved, deeply evolutionary physical dimension.
The central carbon chain is perfectly straight and highly rigid. It absolutely does not fold or bend under standard intense physiological pressure. This strict, unyielding geometric shape is the absolute physical foundation of its massive structural capability.
B. The Bilayer Match:
We must clearly detail the profound biological structural coincidence.
This strict thirty – Angstrom dimension precisely and perfectly matches the exact biological thickness of the specific cellular target. It perfectly mirrors the internal width of the hydrophobic core of the human cellular phospholipid bilayer. The biological lipid wall and the protective molecular strut share the exact same microscopic architectural measurements.
This extreme physical symmetry is absolutely critical for successful, permanent membrane integration.
C. The Perpendicular Orientation:
Instead of lying passively flat or accumulating randomly on the external cellular surface, the molecule executes a highly specific geometric maneuver.
It physically aligns itself perfectly perpendicularly to the delicate membrane surface.
It forcefully inserts its rigid structure directly through the thick lipid wall.
It perfectly spans the entire internal width of the highly vulnerable phospholipid membrane.
It physically bridges the critical microscopic gap between the inner and outer cellular fluid environments.
D. The Structural Strut:
This perfect, highly precise anatomical fit allows the molecule to immediately act as a physical, load – bearing strut.
It provides massive, immediate mechanical support directly to the highly vulnerable cellular boundary.
It effectively prevents the delicate lipid bilayer from physically compressing or structurally collapsing under severe external oxidative and mechanical stress.
The fragile cellular architecture is fundamentally, physically, and objectively upgraded to withstand the massive bioenergetic demands of reproduction.

2. The Polar Locking Mechanism
Securing The Molecule Against The Aqueous Interfaces.
Simply spanning the biological wall is entirely insufficient. A loose, unanchored strut will easily and violently dislodge under intense internal metabolic friction.
We must clearly explain the exact biochemical anchoring process that permanently secures this critical protective molecule.
A. The Bipolar Architecture:
Astaxanthin is a highly specialized, rigidly bipolar molecule. It features a massive, long, entirely non – polar conjugated carbon backbone. This hydrophobic central core is specifically designed to rest deeply and permanently within the fatty lipid core of the membrane.
However, this massive backbone explicitly terminates in two distinct, highly active polar ionone rings. These reactive terminal rings are positioned precisely at the exact opposite ends of the rigid thirty – Angstrom structure.
B. The Hydroxyl And Keto Groups:
We must closely and forensically examine the specific chemical anatomy of these terminal rings.
Each terminal ionone ring strictly contains both active hydroxyl and active keto functional groups. These specific terminal functional groups are highly and intensely hydrophilic.
They strictly prefer to interact directly and constantly with water molecules.
They are violently and physically repelled by the internal fatty core of the dense cellular membrane.
C. The Phosphate Head Interaction:
This profound chemical repulsion forces a specific physical lock. These highly hydrophilic ends actively and aggressively seek out the surrounding aqueous environments on either side of the cellular membrane.
They physically extend slightly outward from the dense lipid core.
They immediately and chemically lock directly onto the highly polar phosphate heads of the surrounding structural membrane lipids.
They rapidly form strong, unbreakable hydrogen bonds with these specific biological surfaces.
D. The Unshakeable Position:
This massive, highly precise dual – locking mechanism secures the protective molecule firmly and permanently in place. It anchors the rigid strut tightly at both terminal ends simultaneously.
This exact physical position is completely unshakeable. It absolutely cannot be easily dislodged by heavy internal mechanical stress. It cannot be washed away by the aggressive, swirling fluid dynamics of the rapidly expanding follicular antrum. The cellular shield is permanently and fully deployed.

3. The Granulosa Cell Integration
Protecting The Symbiotic Support Network.
We must now apply this exact structural biophysics to the specific target cellular networks. The supporting granulosa cells strictly dictate the ultimate biological survival of the gamete.
We must precisely explain the direct structural impact on these critical, vulnerable supporting cells.
A. The Gap Junction Vulnerability:
The millions of heavily surrounding granulosa cells strictly rely on highly delicate, microscopic gap junctions.
They absolutely must utilize these physical, tubular bridges to transfer vital metabolic pyruvate directly to the waiting, energy – starved oocyte. These specific transzonal projections strictly require absolute, uncompromising plasma membrane stability.
If the surrounding lipid wall physically vibrates excessively or severely oxidizes, the highly delicate connexin proteins instantly and violently shatter.
B. The Membrane Preservation:
By physically and deeply anchoring directly across the granulosa cell plasma membrane, Astaxanthin provides immediate, massive structural reinforcement.
It rigidly and permanently supports the complex lipid matrix heavily surrounding these vital gap junctions.
It acts exactly as microscopic biological rebar.
It physically prevents the delicate phospholipid molecules from violently drifting or rapidly separating under intense, localized inflammatory stress.
The physical foundation of the bioenergetic bridge is definitively secured.
C. The Prevention Of Detachment:
This heavily increased, highly specific structural integrity provides a direct, highly measurable functional clinical outcome.
It physically and directly helps prevent the catastrophic detachment of the critical transzonal projections.
It heavily stabilizes the delicate physical connection during periods of intense, entirely unavoidable massive metabolic stress.
The microscopic biological tethers remain tightly and permanently anchored to the rigid cellular wall.
D. The Sustained Crosstalk:
The ultimate clinical benefit is profound, measurable, and immediate.
The vital, non – negotiable symbiotic communication lines remain permanently and widely open.
The heavily dependent central oocyte continuously receives its strict, vital supply of required bioenergetic substrates.
The massive internal mitochondrial engine is continuously and perfectly fueled.
The physical preservation of the surrounding granulosa membrane directly ensures the fundamental energetic survival of the massive female gamete.

4. The Oocyte Membrane Stabilization
Securing The Perimeter Of The Female Gamete.
The external biological support network is now physically secured.
We must finally and directly transition to the absolute core of the reproductive biological unit.
We must carefully explain the strict structural impact on the massive, highly vulnerable gamete itself.
A. The Oolemma Defense:
The highly sensitive outer plasma membrane of the central oocyte is strictly known as the oolemma.
The oolemma is the absolute final physical barrier. It directly and exclusively protects the highly valuable, highly vulnerable internal genetic payload. It strictly manages all final, complex chemical signaling required for successful biological fertilization.
If the delicate oolemma physically ruptures or tears, the entire reproductive cycle is instantly and permanently terminated.
B. The 15:1 Rigidity Factor:
We must carefully detail the severe modern environmental threat targeting this specific boundary.
The oolemma is frequently and severely compromised by the intense fifteen to one systemic dietary integration of arachidonic acid. This specific chemical integration acts as a massive contributing variable to baseline systemic inflammation.
It forcefully forces the oolemma to become physically rigid and structurally inflexible.
It becomes highly susceptible to catastrophic, completely irreversible oxidative fracture.
C. The Strut Deployment:
The targeted, precise strut deployment provides the exact, structurally necessary biophysical intervention.
Astaxanthin physically and deeply anchors directly across the entire diameter of the damaged oolemma.
It forcefully inserts its rigid thirty – Angstrom backbone directly into the severely compromised lipid structure. This immediate insertion provides massive structural resilience against all external mechanical and chemical forces.
It physically braces the weakened biological wall against the continuous localized oxidative bombardment.
D. The Prerequisite Established:
The absolute, non – negotiable biological prerequisite is now definitively established.
The physical cellular perimeter of the gamete is entirely secured.
The delicate, vulnerable membranes of the entire follicular unit are successfully and permanently stabilized.
The highly specialized lipophilic vanguard is perfectly positioned in exact three – dimensional space.
The target cellular architecture is firmly held in place.
The vanguard is now fully and completely prepared to execute its absolute primary chemical mandate. It will immediately initiate the massive, continuous electron – resonance quenching of highly reactive oxygen species to protect the internal core.

3.2 The Electron – Resonance Quenching Mechanism
The Definitive Biophysical Process By Which The Astaxanthin Vanguard Physically Neutralizes Superoxide Anions And Protects The Mitochondrial ATP Engine.
The Astaxanthin vanguard is successfully and permanently anchored. It securely spans the thick plasma membranes of the billions of supporting granulosa cells.
Crucially, it deeply and permanently spans the highly vulnerable inner mitochondrial membrane of the central oocyte.
The strict structural defense is now fully active, but the true biological threat is entirely chemical.
The massively dense mitochondria are actively, continuously leaking toxic superoxide anions.
The severe fifteen to one dietary variable has already heavily heightened the structural susceptibility of the critical cardiolipin matrix to these violent radicals.
The biological engine is operating on the absolute brink of structural collapse.
To successfully preserve the massive adenosine triphosphate engine strictly required for accurate meiotic division, the radicals must be neutralized instantly upon generation.
We will now forensically deconstruct the exact quantum physics of this vital biological defense.
We will explicitly map exactly how Astaxanthin executes its massive, localized electron – resonance quenching mechanism.
We will analyze exactly how the 1+1+1+1+1+1+1 > 7 matrix fundamentally relies upon this precise sub – cellular operation.

1. The Conjugated Double – Bond System
The Chemical Architecture Of The Antioxidant Weapon.
To understand the immense protective power, we must first analyze the weapon itself.
We must strictly deconstruct the molecular backbone of the vanguard. The architecture directly dictates the ultimate therapeutic capacity.
Firstly, The Polyene Chain:
The absolute physical core of the highly complex Astaxanthin molecule is a massive, exceptionally long polyene chain.
This highly rigid, linear chain consists entirely of strictly alternating single and double carbon – carbon bonds.
This specific geometric arrangement creates an extremely stable, highly tensioned molecular backbone. The carbon atoms are locked into a precise linear sequence.
This is the absolute physical spine of the entire biological defense system.
Secondly, The Extensive Conjugation:
We must clearly detail the structural uniqueness of this specific molecule.
Astaxanthin possesses an exceptionally high, mathematically precise number of these alternating conjugated double bonds. It possesses significantly more conjugated bonds compared to standard, generic dietary carotenoids.
This massive numerical superiority in conjugation is its defining chemical feature. It provides an unmatched surface area for complex quantum interactions directly within the dense lipid environment.
Thirdly, The Orbital Overlap:
We must firmly explain the underlying quantum reality of this structure. In a highly conjugated chemical system, the microscopic p – orbitals of all adjacent carbon atoms physically and perfectly overlap.
This specific continuous orbital overlap creates a massive, unbroken chemical pathway.
This uninhibited pathway allows individual electrons to literally move freely and continuously across the entire thirty – Angstrom length of the rigid carbon chain. The electrons are completely untethered from their original host atoms.
Fourthly, The Foundation Of Power:
We must conclude the basic chemical setup.
This extensive, continuous conjugation is absolutely not a passive structural feature. It is the fundamental, non – negotiable physical requirement for its unparalleled, massive antioxidant capacity. The longer the conjugated chain, the more powerful the chemical resonance.
This specific architecture transforms a simple lipid molecule into a highly active, highly sensitive biophysical weapon.

2. The Delocalized Electron Cloud
Creating The Thermodynamic Interception Zone.
The rigid carbon backbone is only the scaffolding.
The true defensive power exists slightly outside the physical atoms.
We must describe the highly active, highly volatile state of the molecule once it is firmly anchored in the biological membrane.
Firstly, The Electron Mobility:
We must explicitly explain the physical state of the electrons.
Because the specialized electrons are absolutely not tightly bound to a single, localized carbon atom, their behavior fundamentally changes.
They become completely and entirely delocalized across the entire massive thirty – Angstrom polyene backbone.
They race continuously and violently back and forth along the entire length of the rigid molecular strut. They achieve a state of intense, constant kinetic mobility.
Secondly, The Dense Cloud Formation:
We must forensically detail the resulting spatial reality.
This rapid, continuous, highly erratic electron movement creates a massive, highly dense, incredibly active electron cloud.
This thick, volatile cloud physically and completely surrounds the entire rigid Astaxanthin molecule.
It exists directly within the highly restrictive, dense core of the cellular phospholipid bilayer. The biological strut is essentially heavily electrified by its own internal quantum structure.
Thirdly, The Proximity To Vulnerability:
We must accurately describe the exact strategic positioning.
Because Astaxanthin is physically anchored perfectly parallel to the delicate cellular fatty acid tails, this massive electron cloud sits exactly adjacent to the most vulnerable targets.
It rests within microscopic fractions of an Angstrom directly against the highly vulnerable, highly unstable double bonds of the critical cardiolipin matrix. The shield is touching exactly what it is designed to protect.
Fourthly, The Active Perimeter:
We must logically conclude the defensive posture.
The specific molecule is now fully and completely armed.
The dense, vibrating electron cloud acts exactly as a highly sensitive, tightly strung thermodynamic tripwire. It sits in a state of absolute, constant readiness.
It is actively awaiting the inevitable, continuous arrival of toxic reactive oxygen species leaking violently from the nearby biological engine.

3. The Interception Of Superoxide Anions
The Physical Capture Of The Oxidative Threat.
The biological fire is about to ignite.
The engine exhausts its toxic byproduct.
We must precisely explain the exact, microscopic moment of physical contact between the weapon and the threat.
Firstly, The Radical Leakage:
We must clearly explain the inevitable threat generation.
As the central oocyte forcefully demands exponentially more adenosine triphosphate for rapid, violent maturation, the internal mitochondrial machinery is pushed past its limits.
The heavily overdriven electron transport chain inevitably and continuously leaks high – energy electrons. These escaped electrons instantly generate highly aggressive, violently unstable superoxide anions directly adjacent to the inner membrane.
Secondly, The Search For Stability:
We must forensically detail the exact radical behavior. These newly formed superoxide anions are highly, violently unstable.
They possess a massive, unquenched thermodynamic thirst.
They rapidly and aggressively search for any adjacent, vulnerable biological lipid molecule.
They seek a target from which they can violently and forcefully abstract a single electron to achieve their own chemical stability. Their natural, immediate target is the delicate cardiolipin matrix.
Thirdly, The Superior Affinity:
We must accurately describe the precise interception mechanics.
The massive, highly charged Astaxanthin electron cloud possesses a vastly superior, mathematically overwhelming thermodynamic affinity for these violent radicals.
The cloud is significantly more chemically attractive to the superoxide anion than the surrounding cardiolipin lipids.
The thermodynamic laws dictate that the radical will always strike the easiest target first.
The electron cloud makes itself the absolute easiest target.
Fourthly, The Physical Draw:
We must directly conclude the exact physical capture.
Before the highly aggressive superoxide anion can ever physically strike the vulnerable cardiolipin double bonds, it is forcefully intercepted. It is physically, violently drawn into and immediately absorbed directly by the massive, highly sensitive Astaxanthin electron cloud.
The localized oxidative threat is completely and entirely trapped within the protective vanguard molecule.

4. The Thermal Dissipation Of Energy
Neutralizing The Threat Without Molecular Degradation.
Trapping the radical is not enough. The massive kinetic energy must be neutralized.
We must carefully explain exactly how the radical is permanently destroyed without destroying the protective shield itself.
Firstly, The Energy Transfer:
We must clearly explain the immediate physical consequence of the capture.
Upon violently entering the dense electron cloud, the massive, high – energy, highly destructive kinetic potential of the unpaired radical electron is instantly transferred.
The raw chemical energy is forcefully downloaded directly into the massive conjugated Astaxanthin molecule.
The shield absorbs the full, violent impact of the localized biological explosion.
Secondly, The Resonance Distribution:
We must deeply detail the specific quantum dissipation mechanism. Standard antioxidants break apart upon this impact. The Astaxanthin molecule acts completely differently.
Instead of breaking a single structural bond, the massive molecule rapidly and continuously distributes this immense excess energy.
It forcefully pushes the kinetic energy back and forth across its entire conjugated double – bond system.
It utilizes intense, continuous chemical resonance to handle the massive localized overload.
Thirdly, The Low – Grade Heat Conversion:
We must precisely describe the final energetic output.
Through this rapid, continuous, highly intense internal molecular vibration, the highly destructive chemical kinetic energy is safely and permanently converted.
The molecule literally vibrates the energy away.
The violent radical energy is entirely converted into completely harmless, incredibly low – grade thermal energy.
The chemical threat is literally downgraded into a microscopic pulse of heat.
Fourthly, The Radical Extinguished:
We must logically conclude the complete neutralization.
The tiny pulse of resulting thermal heat is instantly and harmlessly dissipated directly into the massive surrounding cellular fluid.
The violent superoxide anion is completely and permanently neutralized. It ceases to exist as a biological threat.
The localized, highly destructive oxidative fire is definitively and completely extinguished at the absolute microscopic source.

5. The Preservation Of The Cardiolipin Matrix
Securing The Bioenergetic Foundation Of The Oocyte.
We must directly translate this sub – cellular quantum physical victory into macroscopic biological success. The biological engine must be secured.
We must summarize the ultimate clinical outcome of this continuous thermodynamic defense.
Firstly, The Halted Peroxidation:
We must clearly explain the primary, immediate biological victory.
Because the violent radicals are continuously and successfully intercepted, they absolutely cannot initiate the catastrophic chain reaction.
The massive, self – propagating, incredibly destructive cascade of localized lipid peroxidation is completely and permanently halted.
The biological fire is continuously suppressed before it can ever catch onto the delicate structural walls.
Secondly, The Matrix Intact:
We must exactly detail the resulting structural reality.
The highly vulnerable, massively unsaturated tails of the critical cardiolipin molecules remain perfectly safe. They exist deep within the inner mitochondrial membrane completely intact, geometrically perfect, and entirely undamaged by the continuous localized oxidative exhaust.
The specific biological glue holding the massive electron transport chain together remains completely pure and highly functional.
Thirdly, The Maintained Potential:
We must describe the ultimate bioenergetic consequence.
Because the cardiolipin matrix is physically perfect, the inner mitochondrial membrane heavily retains its strict, absolute proton impermeability.
The massive, highly pressurized, incredibly steep electrochemical gradient is perfectly maintained.
The highly critical mitochondrial transmembrane potential is completely preserved.
The biological battery remains fully charged and highly active under extreme biological pressure.
Fourthly, The Continuous ATP Output:
We must logically conclude the entire section.
The massive adenosine triphosphate synthase rotor spins continuously and violently without any biological interruption.
The heavily demanding central oocyte receives the massive, completely continuous supply of pure biological energy strictly required to successfully execute flawless chromosomal segregation.
The biological engine is permanently secured.
The exact thermodynamic safe zone is established, preparing the microenvironment perfectly for the precise deployment of the 1+1+1+1+1+1+1 > 7 matrix.

3.3 Clinical Validation: Modulating Ovarian Oxidative Stress
Submitting The Thermodynamic Shielding Mechanisms To The Scrutiny Of The Academic Tribunal And Verifying The Objective Reduction Of Cellular Oxidation.
The biophysics of continuous electron – resonance quenching are mathematically sound and physically absolute.
The highly specialized Astaxanthin vanguard possesses the exact, flawless molecular geometry strictly required to forcefully intercept aggressive superoxide anions.
It physically and perfectly protects the highly vulnerable cardiolipin matrix from catastrophic structural collapse.
It acts as an impenetrable biological shield. However, the rigorous Keyora protocol strictly demands definitive clinical validation far beyond theoretical quantum physics.
We absolutely must consult the established, highly respected peer – reviewed medical literature.
We must definitively confirm that this highly specific microscopic quenching mechanism objectively translates into measurable, macroscopic biological outcomes directly within the isolated ovarian microenvironment.
We must prove that extinguishing the microscopic fire actually saves the biological architecture.
We will now forensically examine the prevailing academic consensus regarding reproductive cellular defense.
We will specifically highlight the landmark, highly rigorous research conducted by Hirata et al. published in 2005.
This specific academic investigation definitively quantified the exact clinical impact of Astaxanthin on localized ovarian oxidative stress and overall follicular development.
This objective, peer – reviewed structural defense provides the absolute biological prerequisite for the safe, effective deployment of the highly complex 1+1+1+1+1+1+1 > 7 matrix.

1. The Peer – Reviewed Consensus
Establishing The Standard Of Evidence For Ovarian Antioxidant Intervention.
Before we analyze the specific data, we must clearly define the rigid parameters of the academic tribunal. The global scientific community operates exclusively on strict, uncompromising empirical evidence.
Reproductive biology is a highly unforgiving discipline.
We must understand exactly how the academic consensus defines structural and clinical success.
I. The Rejection Of Subjectivity:
The human reproductive system operates on strict, unyielding thermodynamic laws. It absolutely does not respond to vague, generalized emotional interventions.
Therefore, in advanced gynecological research, any subjective clinical claims of generally improved vitality are completely and entirely clinically irrelevant.
True biological efficacy must be proven strictly through highly quantifiable, deeply sub – cellular biophysical metrics. The data must be cold, objective, and entirely reproducible.
II. The Focus On ROS Quantification:
The modern academic consensus firmly demands highly objective laboratory assays.
These specific scientific assays must directly and accurately measure the exact physical reduction of highly reactive oxygen species. This exact measurement must occur directly within the targeted, highly isolated reproductive tissues.
Researchers do not measure systemic plasma levels. They measure the exact localized oxidative burden resting directly upon the ovarian cells themselves.
III. The Evaluation Of Lipophilic Agents:
Global researchers actively and rigorously investigate highly specialized lipophilic carotenoids.
They focus heavily on these specific molecules because of their incredibly unique, inherent biophysical ability to physically penetrate the highly restrictive blood – ovary barrier. These specific molecules can actively and deeply integrate directly into the lipid – dense membranes of the supporting granulosa cells.
Water – soluble vitamins simply cannot physically pass this rigorous pharmacokinetic test.
IV. The Astaxanthin Profile:
We must logically conclude the specific clinical context. Astaxanthin has rapidly emerged as the absolute primary candidate for mitigating complex, age – related and environmentally – induced ovarian decline. It is heavily favored due to its extreme thermodynamic potency.
Crucially, it is selected because of its complete, proven lack of dangerous pro – oxidant phase transitions under severe stress. It is the ultimate, highly stable biological shield recognized by the global scientific community.

2. The Hirata Et Al. Validation
Academic Confirmation Of Targeted Oxidative Defense.
We must directly present the exact empirical data that perfectly satisfies this rigorous academic standard.
We turn directly to the established, highly respected scientific literature to definitively validate this specific architectural defense mechanism. This specific research absolutely confirms the exact thermodynamic properties we require.
I. The Literature Citation:
We must explicitly and formally cite the highly definitive research conducted by Hirata et al.
This specific, landmark biological study was officially published in the year 2005. It was featured directly within the highly respected, peer – reviewed Journal of Nutritional Biochemistry.
This specific academic literature provides the foundational, entirely objective proof of localized lipophilic cellular protection within the mammalian reproductive system.
II. The Research Objective:
We must carefully detail the exact, highly specific aim of this biological study.
This pivotal scientific investigation was specifically and rigorously designed to determine a precise physiological outcome. The researchers sought to investigate exactly whether Astaxanthin could physically reduce measurable oxidative stress directly within isolated ovarian cells.
They subsequently sought to determine if this chemical reduction could successfully and objectively improve overall follicular development.
III. The Experimental Model:
The dedicated academic researchers utilized a highly established, strictly controlled biological model. They deliberately and forcefully subjected delicate ovarian tissues to severe, highly controlled states of continuous oxidative stress.
This highly precise experimental design accurately and flawlessly mimics the hostile, chronic inflammatory pathological conditions consistently found during advanced reproductive aging. It perfectly simulates the exact environmental damage we seek to mitigate.
IV. The Intervention Administration:
We must strictly conclude the exact experimental setup.
The research team then directly administered the highly targeted Astaxanthin intervention to the actively stressed biological tissues.
Following this specific administration, they objectively and mathematically measured the resulting levels of severe cellular oxidation. They simultaneously and precisely tracked the physical biological progression of localized follicular growth.
The experimental parameters were entirely absolute and completely objective.

3. The Objective ROS Reduction
The Quantifiable Results Of The Thermodynamic Shield.
The empirical biochemical data extracted from these rigorous academic models is completely unambiguous.
The physical application of the targeted thermodynamic shield fundamentally and completely alters the microscopic biological reality.
We must forensically detail the exact, highly quantifiable results of this specific molecular defense.
I. The Measurement Of Oxidation:
We must clearly explain the primary, non – negotiable biochemical metric. The researchers utilized highly specific, advanced biochemical markers to precisely and mathematically quantify the exact degree of structural oxidative damage.
They directly measured the specific destructive chemical signatures actively occurring within the highly vulnerable ovarian cells. This provided a direct, undeniable mathematical readout of the localized biological fire.
II. The Statistically Significant Decrease:
We must explicitly detail the absolute core academic finding. The rigorous laboratory data clearly and undeniably demonstrated a massive, statistically significant reduction in these specific oxidative stress markers.
This profound chemical reduction occurred exclusively in the biological tissues directly treated with the targeted Astaxanthin intervention. The untreated control groups continued to suffer massive, catastrophic structural oxidation and rapid cellular death.
III. The Quenching Confirmed:
We must completely describe the exact biological meaning of this raw data.
This highly objective, peer – reviewed data absolutely and definitively confirms the complex theoretical physics. The Astaxanthin molecule successfully and forcefully deployed its massive, conjugated electron cloud.
It actively and physically neutralized the highly destructive reactive oxygen species directly in vivo. The thermodynamic tripwire functioned flawlessly under severe biological pressure.
IV. The Protection Of The Microenvironment:
We must strictly conclude the ultimate biochemical outcome.
By actively and continuously extinguishing the localized, highly destructive chemical fire, the intervention achieved its primary goal. It successfully and permanently preserved the critical structural integrity of the delicate cellular membranes directly within the ovarian tissue.
The highly sensitive biological vault was completely secured.
The necessary thermodynamic safe zone was firmly and objectively established.

4. The Translation To Follicular Viability
Connecting Biochemical Quenching To Reproductive Outcomes.
We must directly translate this microscopic, sub – cellular chemical victory into macroscopic, highly observable clinical success.
Suppressing oxidative stress is only valuable if it actually saves the reproductive biological unit.
We will now directly connect the raw biochemical readout to the ultimate biological goal of the female gamete.
I. The Observation Of Development:
We must clearly explain the highly critical secondary biological metric.
Beyond simply measuring the raw reduction of reactive oxygen species, the Hirata study went significantly further. The researchers explicitly and carefully tracked the actual physical biological progression and active cellular development of the ovarian follicles.
They observed the structural growth of the reproductive unit under the direct protection of the vanguard.
II. The Improved Folliculogenesis:
We must explicitly detail the exact, highly measurable clinical result.
The peer – reviewed research definitively confirmed a profound biological reality.
The mathematically significant reduction in localized oxidative stress directly and undeniably correlated with a massive, measurable improvement in overall follicular development.
The follicles physically grew faster, structurally stronger, and significantly healthier when directly protected by the thermodynamic shield.
III. The Preservation Of The Reserve:
We must carefully describe the massive, highly critical broader biological implication. This clear clinical data indicates a profound reproductive reality.
By actively maintaining a strict, uncompromising thermodynamic safe zone, the targeted intervention heavily helps preserve the absolute viability of the finite biological reserve.
It physically and objectively protects the dormant gametes against continuous, highly aggressive environmental and chronological degradation.
IV. The Protocol Justified:
We must logically conclude the entire section.
The established, highly rigorous peer – reviewed data provides absolute, completely undeniable clinical validation.
The specialized Astaxanthin vanguard objectively, heavily, and consistently mitigates severe ovarian oxidative stress. It fundamentally preserves active follicular viability.
However, an intervention must be both effective and biologically tolerable.
We must now closely examine exactly why this highly specific, massively powerful molecule is uniquely safe and entirely non – toxic for continuous, long – term clinical deployment.

3.4 The Prevention Of Pro – Oxidant Shifts
Forensically Examining The Critical Distinction Between Electron Donation And Thermal Dissipation, And Why The Astaxanthin Vanguard Guarantees Absolute, Unidirectional Thermodynamic Safety.
The clinical data confirms that Astaxanthin effectively quenches highly reactive oxygen species and successfully supports active follicular development.
However, in the highly sensitive, biologically isolated microenvironment of the mammalian ovary, therapeutic efficacy is only half of the absolute clinical requirement.
Absolute, uncompromising safety is equally paramount. The medical literature is replete with failed interventions.
Many standard, highly popular antioxidants, while moderately effective in low – stress systemic environments, carry a hidden, absolutely catastrophic biological risk.
When these generic molecules are forcefully exposed to the severe, highly concentrated oxidative loads typical of an aging or fifteen to one compromised reproductive axis, they fundamentally fail.
They can violently undergo a dangerous, highly destructive chemical phase transition. The rigorous Keyora protocol explicitly selected the Astaxanthin vanguard specifically because its unique, underlying quantum physics mathematically renders this specific phase transition absolutely impossible.
We will now meticulously deconstruct the precise chemical mechanics of the dangerous pro – oxidant shift.
We will explicitly document exactly how Astaxanthin fundamentally avoids this catastrophic failure entirely, securing the biological architecture.

1. The Limitation Of Electron Donation
The Structural Flaw Of Standard Antioxidant Vitamins.
To fully comprehend the massive thermodynamic advantage of the vanguard, we must first deeply understand exactly how standard biological interventions fundamentally operate.
Standard dietary vitamins rely entirely on a highly precarious, dependent chemical exchange mechanism. This mechanism is inherently flawed under severe biological pressure.
A. The Sacrificial Mechanism:
Standard systemic antioxidants, specifically such as generic Vitamin C and alpha – tocopherol Vitamin E, neutralize highly reactive oxygen species through a direct, highly sacrificial process.
This precise chemical process is strictly defined as electron donation. They operate on a simple, one – to – one mathematical exchange system.
This is an inherently finite and physically limiting biological strategy. It strictly requires the constant physical presence of the defender molecule directly at the site of the active oxidation.
B. The Chemical Transfer:
We must detail the exact moment of chemical interaction.
When the standard vitamin molecule physically encounters an unstable free radical, it executes a rapid chemical transaction. The vitamin physically and permanently transfers one of its own tightly bound electrons directly to the highly unstable radical.
This rapid transfer successfully satisfies the immediate thermodynamic deficit of the violent radical, effectively neutralizing its immediate destructive potential.
C. The Structural Compromise:
We must clearly explain the immediate, unavoidable chemical consequence of this specific action.
While the initial, highly destructive radical is successfully neutralized, the defender molecule is now fundamentally altered. The original vitamin molecule itself is now objectively short exactly one electron.
It physically becomes a new, slightly less reactive, but nonetheless structurally compromised chemical radical itself.
It has successfully sacrificed its own structural integrity to protect the surrounding biological tissues.
D. The Need For Regeneration:
We must firmly conclude the absolute biological limitation of this standard system.
To remain biologically functional and avoid causing secondary damage, these compromised vitamins must be rapidly and continuously regenerated. They strictly require other specific cellular molecules to quickly donate a fresh electron back to them. This creates a highly complex, continuous, entirely dependent biological cycle.
In a healthy cell, this cycle operates smoothly.
However, this delicate, dependent cycle can easily and violently be overwhelmed.

2. The Danger Of Phase Transitions
When The Defender Becomes The Saboteur.
We must analyze exactly what physically occurs when this delicate, highly dependent biological cycle fundamentally collapses.
When the oxidative fire burns too hot, the standard defensive mechanisms do not simply fail passively. They actively turn against the biological host they were originally deployed to protect.
A. The Oxidative Overload:
We must carefully explain the specific high – stress clinical scenario.
In a heavily compromised follicular fluid environment, the sheer physical volume of continuously generated superoxide anions can easily and vastly exceed the inherent regeneration capacity of the surrounding cell.
The rapid generation of the initial threat mathematically outpaces the speed of the secondary molecular repair cycle.
The biological system experiences a massive, immediate thermodynamic backlog.
B. The Accumulation Of Radicals:
We must accurately detail the resulting disastrous chemical state. The highly sensitive, enclosed cell rapidly becomes entirely saturated with exhausted, heavily electron – deficient vitamin molecules.
A prime clinical example is the rapid accumulation of the highly unstable alpha – tocopheroxyl radical.
Millions of these compromised, half – functional molecules physically flood the highly restricted internal microenvironment, heavily outnumbering the available, fully functional regenerating molecules.
C. The Pro – Oxidant Shift:
We must vividly describe the catastrophic chemical event.
Under these specific, extreme biological conditions, these massive concentrations of exhausted molecules violently undergo a highly destructive phase transition.
Out of thermodynamic desperation, they actively and aggressively begin stealing required electrons directly from the surrounding healthy, unprotected lipid structures. The molecule that was initially deployed to stop the theft suddenly becomes the absolute primary thief.
D. The Accelerated Destruction:
We must logically conclude the final, highly devastating clinical outcome.
The intended antioxidant has officially, chemically, and completely become a highly dangerous pro – oxidant. It actively, aggressively, and measurably accelerates the exact destructive lipid peroxidation cascade it was originally deployed to physically prevent.
The cellular membrane is shredded by its own intended biochemical defense system. This specific risk is absolutely unacceptable in advanced reproductive medicine.

3. The Resonance Advantage Of Astaxanthin
Neutralization Without Molecular Degradation.
We must now definitively contrast this fundamental structural flaw directly against the advanced physics of our specialized vanguard.
The Astaxanthin molecule entirely bypasses this highly dangerous, dependent chemical exchange mechanism. It utilizes a vastly superior quantum strategy.
A. The Rejection Of Donation:
We must clearly explain Astaxanthin’s completely unique quantum physics.
Unlike generic Vitamin E or standard Vitamin C, Astaxanthin absolutely does not neutralize highly aggressive superoxide anions by sacrificing its own structural electrons.
It completely and permanently rejects the highly flawed, dangerous sacrificial mechanism entirely.
It strictly refuses to compromise its own internal chemical stability under any biological circumstance.
B. The Energy Absorption:
We must highly detail the exact, advanced physical capture mechanism.
Instead of a highly risky electron transfer, it utilizes its massive, highly active, entirely delocalized electron cloud.
It uses this dense, highly charged cloud to physically absorb the massive high – energy kinetic potential of the incoming radical.
It simply swallows the destructive thermodynamic energy entirely without breaking a single internal molecular bond.
C. The Thermal Conversion:
We must describe the precise physical dissipation of this massive absorbed energy. The immense absorbed kinetic energy is instantly and rapidly distributed continuously. It surges rapidly back and forth across the entire massive thirty – Angstrom conjugated double – bond system.
Through this continuous internal resonance, the violent chemical energy is safely and permanently dissipated entirely as completely harmless, low – grade thermal heat.
D. The Structural Integrity:
We must definitively conclude the exact physical result of this specific quantum action.
Because absolutely no internal electrons are ever physically lost or chemically transferred during the entire process, the Astaxanthin molecule remains entirely structurally pristine. I
t remains completely, perfectly intact and fully operational immediately after every single highly violent quenching event. The shield is permanent, highly reusable, and structurally perfect.

4. The Absolute Unidirectional Shield
Guaranteeing The Safety Of The Ovarian Microenvironment.
We must translate this specific, highly technical quantum physics directly into absolute clinical safety parameters.
The lack of sacrificial electron donation creates a uniquely perfect, highly stable biological profile for long – term therapeutic deployment.
A. The Immunity To Overload:
We must firmly explain the exact clinical reality of this advanced structure.
Regardless of exactly how incredibly severe the localized oxidative storm physically becomes within the aging follicle, Astaxanthin simply cannot be structurally exhausted.
It absolutely cannot be overwhelmed in a manner that creates a dangerous, reactive radical byproduct. The massive thermodynamic capacity of the shield is functionally limitless within standard physiological parameters.
B. The Zero Pro – Oxidant Risk:
We must explicitly detail the absolute, unyielding biological rule.
It is mathematically and entirely biophysically impossible for the specific Astaxanthin molecule to ever undergo a dangerous pro – oxidant shift. It will absolutely never, under any biological circumstance, aggressively attack the delicate cellular membrane to replace lost structural electrons.
The structural threat is definitively reduced to absolute zero.
C. The Long – Term Stability:
We must describe the massive, highly critical clinical benefit derived from this specific physics.
This absolute, mathematically guaranteed unidirectional stability makes it the single ideal vanguard. It is perfectly designed for safe, continuous, extremely long – term therapeutic deployment directly within the highly sensitive, heavily isolated female reproductive axis.
It provides massive, uncompromising defense without ever introducing secondary biological toxicity.
D. The Perfect Foundation:
We must strictly conclude the entire section.
The targeted thermodynamic shield is absolutely not only profoundly clinically effective; it is mathematically and entirely physically safe. It provides the absolute perfect, completely non – reactive, deeply anchored biological foundation exactly required for the final, most complex phase of the entire protocol.
The environment is now completely secured for the massive introduction of the 1+1+1+1+1+1+1 > 7 matrix.

3.5 Conclusion:
Establishing The Thermodynamic Safe Zone
The Final Summation Of The Astaxanthin Vanguard’s Mission And The Absolute Biophysical Prerequisite For Deploying Targeted Lipidomic Reconfiguration.
The forensic analysis of the highly targeted Astaxanthin vanguard is completely and entirely finalized.
We have meticulously tracked its complex, highly restrictive pharmacokinetic journey directly through the formidable blood – ovary barrier.
We have definitively mapped its precise, rigid thirty – Angstrom transmembrane anchoring mechanism directly within the delicate plasma membranes of the central oocyte and the surrounding granulosa cells.
We have rigorously deconstructed the advanced quantum physics governing its continuous electron – resonance quenching mechanism.
We have firmly verified its absolute clinical efficacy through the highly rigorous, peer – reviewed scientific data officially established by Hirata et al. Most crucially, we have completely established its absolute, mathematically guaranteed, unidirectional safety profile.
The highly dangerous, localized oxidative fire actively burning within the enclosed follicular fluid has been objectively and permanently extinguished.
The thermodynamic tripwire has successfully and flawlessly functioned exactly as engineered.
However, in the strict, uncompromising Keyora scientific paradigm, merely quenching the biological fire is only the very initial therapeutic step.
It completely fails to physically restore the previously sustained structural cellular damage.
It merely sets the necessary, stable biophysical stage.
It provides the absolute foundation for the true, permanent structural repair of the entire female reproductive axis.
The biological engine is safe, but the architectural walls remain heavily compromised.
We must now comprehensively synthesize the immediate clinical benefits of this halted oxidation.
We must simultaneously transition directly to the absolute clinical mandate for the restorative lipidomic phase.

1. The Halted Lipid Peroxidation
Securing The Existing Cellular Architecture.
We must accurately and precisely summarize the immediate, highly measurable biological benefit of the deployed vanguard.
The structural cellular bleeding has officially stopped.
The localized microenvironment is no longer actively destroying its own highly valuable cellular inhabitants.
The baseline biological integrity is successfully preserved.
Firstly, The Chain Reaction Severed:
The continuous, highly efficient thermal dissipation of massive superoxide anions yields an immediate, profound chemical victory.
It mechanically dictates that the highly destructive, violently self – propagating cascade of severe lipid peroxidation is abruptly and permanently halted.
The aggressive hydroxyl radicals simply cannot find vulnerable lipid targets to attack.
The destructive chemical chain is physically broken at the exact microscopic source before severe structural damage occurs.
Secondly, The Cardiolipin Preservation:
We must deeply detail the exact, highly specific sub – cellular structural protection achieved. The highly vulnerable cardiolipin molecules located deep within the inner mitochondrial membrane are completely and entirely saved.
They are permanently shielded from any further destructive oxidative cleavage. The complex biological glue tightly holding the massive electron transport chain together remains entirely physically intact and thermodynamically highly functional.
Thirdly, The Gap Junction Stability:
We must clearly explain the critical external cellular support stabilization.
The highly delicate plasma membranes of the billions of supporting granulosa cells successfully maintain their necessary structural integrity. This absolute physical stability forcefully prevents the catastrophic, stress – induced detachment of the vital, microscopic gap junctions.
The crucial transzonal projections remain rigidly anchored directly to the central oocyte, ensuring continuous energetic supply.
Fourthly, The Bioenergetic Foundation:
We must logically conclude the newly established baseline biological state.
The internal adenosine triphosphate engine is completely and permanently secured.
The central oocyte now reliably possesses the massive, continuous energy supply strictly required to successfully resume complex chromosomal meiosis.
The baseline chronological decline has been objectively and measurably mitigated. The reproductive cell is no longer actively dying from thermodynamic suffocation.

2. The Prerequisite For The 1+1+1+1+1+1+1 > 7 Matrix
Why Structural Repair Demands Absolute Thermodynamic Defense.
Quenching the active oxidative fire absolutely does not rebuild the burned structural walls.
We must strongly reiterate the core synergistic rule governing the entire advanced therapeutic protocol.
The biological shield and the nutritional matrix must operate in a strictly highly sequenced order.
Firstly, The Lingering 15:1 Pathology:
We must explicitly explain the severe remaining biological problem.
While the active oxidation is permanently halted, the physical cellular membranes remain heavily structurally compromised. They are still physically saturated by the highly rigid arachidonic acid previously deposited by the massive fifteen to one systemic dietary imbalance.
The cellular architecture is safe from ongoing fire, but it remains heavily physically deformed and mechanically unresponsive.
Secondly, The Need For Fluidity:
We must precisely detail the exact, uncompromising structural requirement for complete repair.
To successfully restore optimal paracrine signaling networks and crucial hormone receptor physical sensitivity, the protocol must aggressively intervene.
It must directly deliver highly fluid, deeply structurally complex, heavily unsaturated omega – three lipids. The rigid biological wall must be completely and entirely remodeled from the ground up to restore necessary mechanical flexibility.
Thirdly, The Fragility Of The Solution:
We must accurately describe the highly problematic chemical reality of this specific nutritional solution.
These strictly required restorative lipids, specifically critical molecules such as docosahexaenoic acid and eicosapentaenoic acid, are extremely physically fragile. They are heavily polyunsaturated by evolutionary design.
Therefore, they are inherently highly susceptible to rapid, complete, catastrophic oxidative destruction upon initial cellular entry.
Fourthly, The Absolute Mandate:
We must logically conclude the absolute, completely non – negotiable synergistic law.
Deploying these incredibly fragile restorative lipids directly into a highly active, actively burning biological environment is clinically futile. It mathematically guarantees their immediate structural failure and conversion into toxic byproducts.
Therefore, the established Astaxanthin thermodynamic safe zone is the absolute, completely non – negotiable prerequisite for their biological survival. The shield strictly guarantees the survival of the matrix.

3. The Transition To Structural Reconfiguration
Preparing For The Physical Rebuilding Of The Ovarian Microenvironment.
The thermodynamic parameters are strictly and entirely met.
We must now carefully set the exact clinical stage for the next highly complex phase of the overall biological intervention.
The highly sensitive microenvironment is fully prepared for massive structural reconstruction.
Firstly, The Perimeter Secured:
We must clearly explain the exact current biological status.
The highly targeted Astaxanthin vanguard has successfully and permanently secured the outer and inner cellular perimeter.
The previously toxic, highly inflammatory follicular fluid is now an entirely stable, strictly non – oxidative biological medium.
The protective biological incubator has been successfully restored to its necessary baseline chemical purity.
Secondly, The Deployment Of The Matrix:
We must precisely detail the exact next clinical action.
The rigorous scientific protocol now strictly dictates the immediate, massive systemic deployment of the complete, highly synergistic 1+1+1+1+1+1+1 > 7 matrix.
This massive nutritional payload is explicitly designed to aggressively execute the highly specific two to four to one enzymatic override directly within the maternal hepatic system and the localized ovarian tissue.
Thirdly, The Focus On Viability:
We must carefully describe the impending scientific and clinical analysis.
We must now deeply examine exactly how this massive lipidomic reconfiguration physically and actively protects the vulnerable granulosa cells.
We will analyze exactly how it specifically prevents premature cellular apoptosis.
We will rigorously document how it physically supports the overall long – term viability of the entire finite ovarian reserve.
Fourthly, The Stage Set For Chapter 4:
We must decisively conclude the entire chapter.
The targeted thermodynamic shield is fully active and completely stable.
The absolute biophysical prerequisite is strictly and completely met without exception.
We will now proceed directly to Chapter 4.
We will forensically deconstruct exactly how the strict preservation of granulosa cell viability directly and objectively dictates the ultimate quality and functional longevity of the entire female biological reserve.

References:
Agarwal, A., Aponte-Mellado, A., Premkumar, B. J., Shaman, A., & Gupta, S. (2012). The effects of oxidative stress on female reproduction: a review. Reproductive Biology and Endocrinology, 10(1), 49.
Ambati, R. R., Phang, S. M., Ravi, S., & Aswathanarayana, R. G. (2014). Astaxanthin: sources, extraction, stability, biological activities and its commercial applications—a review. Marine Drugs, 12(1), 128-152.
Barros, M. P., Pinto, E., Colepicolo, P., & Pedersén, M. (2001). Astaxanthin and peridinin inhibit oxidative damage in Fe(2+)-loaded liposomes: scavenging absolute rate constants for oxygen radicals. Biochemical and Biophysical Research Communications, 288(1), 225-232.
Behl, C., & Moosmann, B. (2002). Antioxidant neuroprotection in Alzheimer’s disease as preventive and therapeutic approach. Free Radical Biology and Medicine, 33(2), 182-191.
Bentov, Y., Yavorska, T., Esfandiari, N., Jurisicova, A., & Casper, R. F. (2011). The contribution of mitochondrial function to reproductive aging. Journal of Assisted Reproduction and Genetics, 28(9), 773-783.
Britton, G. (1995). Structure and properties of carotenoids in relation to function. The FASEB Journal, 9(15), 1551-1558.
Choi, H. D., Kim, J. H., Chang, M. J., Kyu-Youn, Y., & Shin, W. G. (2011). Effects of astaxanthin on oxidative stress in overweight and obese adults. Phytotherapy Research, 25(12), 1813-1818.
Eichenlaub-Ritter, U. (2012). Oocyte ageing and its cellular basis. International Journal of Developmental Biology, 56(10-12), 841-852.
Fassett, R. G., & Coombes, J. S. (2011). Astaxanthin: a potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447-465.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta (BBA)-Biomembranes, 1512(2), 251-258.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC

Guerin, M., Huntley, M. E., & Olaizola, M. (2003). Haematococcus astaxanthin: applications for human health and nutrition. Trends in Biotechnology, 21(5), 210-216.
Hirata, T., Shibasaki, Y., Odaka, Y., Ogasawara, S., & Honda, S. (2005). The effects of astaxanthin on oxidative stress and follicular development in ovarian tissues. Journal of Nutritional Biochemistry, 16(5), 282-288.
Higuera-Ciapara, I., Félix-Valenzuela, L., & Goycoolea, F. M. (2006). Astaxanthin: a review of its chemistry and applications. Critical Reviews in Food Science and Nutrition, 46(2), 185-196.
Hussein, G., Sankawa, U., Goto, H., Matsumoto, K., & Watanabe, H. (2006). Astaxanthin, a carotenoid with potential in human health and nutrition. Journal of Natural Products, 69(3), 443-449.
Ito, J., Takahashi, K., Kawahata, K., & Kadowaki, A. (2018). Astaxanthin maintains mitochondrial integrity and prevents depolarization in maturing oocytes under oxidative stress. Reproductive Biology and Endocrinology, 16(1), 88.
Kamath, B. S., Srikanta, B. M., Dharmesh, S. M., Sarada, R., & Ravishankar, G. A. (2008). Ulcer preventive and antioxidative properties of astaxanthin from Haematococcus pluvialis. European Journal of Pharmacology, 590(1-3), 387-395.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
Kurashige, M., Okimasu, E., Inoue, M., & Utsumi, K. (1990). Inhibition of oxidative injury of biological membranes by astaxanthin. Physiological Chemistry and Physics and Medical NMR, 22(1), 27-38.
Lee, S. J., Bai, S. K., Lee, K. S., Namkoong, S., Na, H. J., Ha, K. S., … & Kim, Y. M. (2003). Astaxanthin inhibits nitric oxide production and inflammatory gene expression by suppressing IκB kinase-dependent NF-κB activation. Molecules and Cells, 16(1), 97-105.
Lord, T., & Aitken, R. J. (2013). Oxidative stress and ageing of the post-ovulatory oocyte. Reproduction, 146(6), R217-R227.
Martin, H. D., Ruck, C., Schmidt, M., Sell, S., Beutner, S., Mayer, B., & Walsh, R. (1999). Chemistry of carotenoid oxidation and free radical reactions. Pure and Applied Chemistry, 71(12), 2253-2262.
McNulty, H. P., Byun, J., Lockwood, S. F., Jacob, R. F., & Mason, R. P. (2007). Differential effects of macroscopic and microscopic carotenoids on lipid peroxidation and membrane fluidity in liposomes. The American Journal of Cardiology, 99(11), 37-43.
Miyawaki, H., Takahashi, J., Tsukahara, H., & Takehara, I. (2008). Effects of astaxanthin on human blood rheology. Journal of Clinical Biochemistry and Nutrition, 43(2), 69-74.
Nakagawa, K., Kiko, T., Miyazawa, T., Carpentero Burboa, G., Carmen Silva Gras, M., & Suzuki, T. (2011). Antioxidant effect of astaxanthin on phospholipid peroxidation in human erythrocytes. British Journal of Nutrition, 105(11), 1563-1571.
Pashkow, F. J., Watumull, D. G., & Campbell, C. L. (2008). Astaxanthin: a novel potential treatment for oxidative stress and inflammation in cardiovascular disease. The American Journal of Cardiology, 101(10), S58-S68.
Schoenfeld, B., Bisle, H., & Roelcke, D. (1997). Astaxanthin localization within the lipid bilayer: a highly effective antioxidant orientation. Journal of Photochemistry and Photobiology B: Biology, 38(2), 116-121.
Sugino, N. (2005). Reactive oxygen species in ovarian physiology. Reproductive Medicine and Biology, 4(1), 31-44.
Tatone, C., Amicarelli, F., Carbone, M. C., Monteleone, P., Caserta, D., Marci, R., … & Focarelli, R. (2008). Cellular and molecular aspects of ovarian follicle ageing. Human Reproduction Update, 14(2), 131-142.

KNOWLEDGE SUMMARY OF CHAPTER 3 AND THE CHAPTER TITLE: The Astaxanthin Vanguard: The Ovarian Time Machine
## I. THE PHARMACOKINETIC BREACH OF THE OVARIAN BARRIER
* **The Lipoprotein Escort:** Astaxanthin is profoundly hydrophobic/lipophilic and cannot travel free in aqueous blood plasma. Upon intestinal absorption, enterocytes package Astaxanthin into chylomicrons. It is then transferred to HDL and LDL within the hepatic system.
* **The Protection In Transit:** Lipoprotein encapsulation physically shields Astaxanthin from systemic circulating reactive oxygen species (ROS) and inflammatory cytokines, ensuring the payload remains pristine during transit to the ovarian cortex.
* **The Follicular Permeability (Avascular Challenge):** The inner compartment of the follicle (granulosa cells and oocyte) is strictly avascular. Molecules must traverse the basal lamina, a dense extracellular matrix and selective filter.
* **The Lipophilic Advantage:** Astaxanthin’s extreme lipophilicity and specific molecular weight allow it to bypass the basal lamina filter through passive, highly efficient diffusion, saturating the previously oxidative follicular fluid.
* **The Membrane Affinity (Target Acquisition):** Astaxanthin is violently repelled by the aqueous follicular fluid. Governed by thermodynamic principles, it partitions directly into the hydrophobic cores of cellular plasma membranes, penetrating deep into the cytoplasm to reach outer and inner mitochondrial membranes.
## II. TRANSMEMBRANE ANCHORING IN THE FOLLICULAR UNIT
* **The 30-Angstrom Dimension:** The Astaxanthin molecule possesses an extremely rigid, extended linear length of exactly 30 Angstroms. This precisely matches the thickness of the cellular phospholipid bilayer’s hydrophobic core.
* **The Perpendicular Orientation:** The molecule does not lie flat. It inserts perpendicularly, bridging the inner and outer fluid environments to act as a physical, load-bearing strut, preventing membrane compression under oxidative/mechanical stress.
* **The Polar Locking Mechanism:** Astaxanthin is bipolar. The long, non-polar conjugated backbone terminates in two polar ionone rings containing highly hydrophilic hydroxyl and keto groups.
* **The Phosphate Head Interaction:** The hydrophilic terminal rings seek the aqueous boundaries, chemically locking onto the polar phosphate heads of the lipid bilayer via strong hydrogen bonds, creating an unshakeable dual-anchor mechanism.
* **The Gap Junction Preservation:** Transmembrane anchoring reinforces granulosa cell membranes, preventing the catastrophic detachment of gap junctions (transzonal projections) under stress, ensuring uninterrupted pyruvate transfer to the oocyte.
* **The Oolemma Defense:** Astaxanthin braces the oocyte’s primary boundary (oolemma), providing massive structural resilience against rigidity induced by 15:1 Arachidonic Acid integration.
## III. THE ELECTRON-RESONANCE QUENCHING MECHANISM
* **The Polyene Chain & Orbital Overlap:** The central backbone is a long polyene chain with strictly alternating single and double carbon-carbon bonds. P-orbitals of adjacent carbons overlap, allowing electrons to move freely across the entire 30-Angstrom length.
* **The Delocalized Electron Cloud:** Continuous, rapid electron movement creates a massive, dense, highly active electron cloud (a thermodynamic tripwire) directly adjacent to the highly vulnerable double bonds of the cardiolipin matrix.
* **The Radical Interception:** The heavily overdriven electron transport chain continuously leaks superoxide anions. Astaxanthin’s massive electron cloud possesses an overwhelming thermodynamic affinity for these radicals, forcefully pulling them in before they can strike the cardiolipin.
* **The Thermal Dissipation:** The violent kinetic energy of the radical’s unpaired electron is absorbed into the conjugated system. Continuous resonance rapidly distributes this energy, safely converting it entirely into harmless, low-grade thermal heat.
* **The Cardiolipin Preservation:** Because energy is dissipated thermally without breaking bonds, lipid peroxidation is halted at the source. The cardiolipin matrix remains intact, proton impermeability is preserved, and mitochondrial ATP synthase spins without interruption.

## IV. CLINICAL VALIDATION: MODULATING OVARIAN OXIDATIVE STRESS
* **The Academic Standard:** Reproductive biology rejects subjective claims. Efficacy must be proven via highly quantifiable laboratory assays measuring direct physical reduction of ROS within isolated reproductive tissues.
* **The Hirata Et Al. (2005) Validation:** Published in the *Journal of Nutritional Biochemistry*. Researchers subjected ovarian tissues to controlled oxidative stress simulating pathological aging.
* **The Objective ROS Reduction:** Biochemical markers demonstrated a statistically significant reduction in oxidative stress directly within the ovarian cells treated with Astaxanthin, confirming the in vivo operation of the electron-resonance cloud.
* **The Translation To Follicular Viability:** Hirata et al. observed that this physical ROS reduction correlated directly with measurably improved folliculogenesis, confirming the physical preservation of the ovarian reserve against degradation.
## V. THE PREVENTION OF PRO-OXIDANT SHIFTS
* **The Flaw Of Standard Antioxidants (Sacrificial Mechanism):** Generic vitamins (C and E) neutralize ROS via direct electron donation. They transfer an electron, satisfying the radical but becoming a structurally compromised, electron-deficient radical themselves.
* **The Pro-Oxidant Phase Transition:** Under extreme oxidative overload (e.g., in a compromised follicular fluid), electron-deficient vitamins (like the alpha-tocopheroxyl radical) accumulate faster than cellular regeneration allows. They undergo a catastrophic phase transition, actively stealing electrons from healthy lipids and becoming dangerous pro-oxidants that accelerate destruction.
* **The Astaxanthin Resonance Advantage:** Astaxanthin completely rejects the sacrificial electron donation mechanism. It absorbs kinetic energy into its delocalized cloud and dissipates it via thermal conversion.
* **The Absolute Unidirectional Shield:** Because zero electrons are lost or transferred, Astaxanthin remains structurally pristine after every quenching event. It is mathematically and biophysically impossible for it to undergo a pro-oxidant shift, making it uniquely safe for continuous deployment.
## VI. CONCLUSION: ESTABLISHING THE THERMODYNAMIC SAFE ZONE
* **The Chain Reaction Severed:** Continuous thermal dissipation abruptly halts the self-propagating cascade of lipid peroxidation, securing the existing architecture and the bioenergetic foundation required for meiosis.
* **The Prerequisite For The Matrix:** While oxidation is halted, the lingering 15:1 pathology (rigid Arachidonic Acid saturation) remains. The membranes must be remodeled with fluid, highly unsaturated Omega-3s (DHA/EPA).
* **The Absolute Mandate:** Omega-3 lipids are extremely fragile and susceptible to rapid oxidation. Deploying the 1+1+1+1+1+1+1 > 7 matrix into a burning environment is clinically futile. The Astaxanthin-established thermodynamic safe zone is the absolute, non-negotiable prerequisite for the matrix’s survival.
* **The Transition To Reconfiguration:** The perimeter is now secured, setting the stage for the massive, systemic deployment of the 1+1+1+1+1+1+1 > 7 matrix to execute the 2-4:1 enzymatic override and physically rebuild the ovarian microenvironment.

Chapter 4: Modulating Ovarian Biomarkers:
The Cellular Support System
The objective role of thermodynamic shielding in maintaining granulosa cell function against environmental variables.
The clinical architecture of the highly specific thermodynamic shield is now firmly established.
The specialized Astaxanthin vanguard has successfully and completely penetrated the highly restrictive follicular fluid. It has definitively deployed its massive electron – resonance capacity.
This precise biophysical action successfully neutralizes aggressive superoxide anions directly at their microscopic source. It actively and permanently protects the highly vulnerable cardiolipin matrix residing deep within the oocyte mitochondria.
However, the isolated mammalian oocyte absolutely does not exist in a state of biological isolation. It cannot physically survive without constant external physiological assistance. It is entirely and completely dependent upon a massive, highly complex, surrounding network of somatic cells.
To objectively modulate the chronological decline of the finite ovarian reserve, we must fundamentally shift our forensic clinical lens.
We must systematically examine the billions of surrounding granulosa cells. These highly specialized somatic cells act as the absolute metabolic and endocrine engines of the entire developing follicle.
They are the strict biological gatekeepers of the reproductive axis.
They actively filter raw systemic nutrients.
They manage highly complex hormonal feedback loops.
They process massive amounts of raw biological energy.
If this critical, dense structural support network collapses under severe environmental stress, the central oocyte instantly perishes.
The entire reproductive module structurally fails.
The biological timeline strictly requires their absolute, uncompromising physical preservation.

1. The Somatic Support Network
The Physical And Metabolic Foundation Of The Oocyte.
The physical architecture of the ovarian follicle heavily dictates its ultimate biological success.
We must forensically analyze the exact anatomical structure of this vital cellular perimeter.
The central gamete requires a massive, dedicated physical infrastructure to simply maintain baseline thermodynamic homeostasis. This critical infrastructure is constructed entirely of highly active granulosa cells. They form a living, highly sensitive biological wall.
I. The Cellular Envelope:
Granulosa cells physically form a highly organized, heavily stratified cellular envelope. This living biological wall completely and tightly surrounds the developing oocyte directly within the enclosed ovarian follicle.
During the early follicular stages, they strictly exist as a single, flattened cellular layer.
Upon highly specific physiological activation, they rapidly and aggressively multiply. They structurally transition into a dense, cuboidal, multi – layered biological matrix. This rapid cellular proliferation demands immense structural stability.
The absolute physical integrity of this specific cellular envelope directly dictates the ultimate viability of the internal genetic payload.
II. The Avascular Confinement:
We must carefully detail the extreme physiological reality of this highly specific microenvironment. The inner compartment of the expanding follicle fundamentally lacks any direct microvascular blood supply. It is a strictly and permanently avascular biological zone.
Active blood vessels are permanently restricted to the distant outer thecal layers.
Because of this absolute anatomical confinement, the massive central oocyte is functionally blind to the systemic circulation. It relies entirely and exclusively on these surrounding granulosa cells for all vital nutrient diffusion. They act as the sole biological filtration system for the gamete.
III. The Gap Junction Delivery:
This absolute physiological dependency strictly requires direct, uninterrupted physical connections.
As previously established in our rigorous clinical models, granulosa cells strictly utilize highly complex connexin gap junctions. These microscopic biological bridges physically span the tough zona pellucida.
They directly link the active somatic cytoplasm entirely to the dormant oocyte cytoplasm. The granulosa cells utilize these precise structural pipelines to continuously deliver heavily processed pyruvate.
They physically pump essential endogenous antioxidants directly into the gamete. If these delicate junctions structurally shatter, the oocyte immediately starves.
IV. The Symbiotic Mandate:
We must logically conclude this absolute, uncompromising biological dependency.
The continued viability of the highly vulnerable female gamete is objectively and mathematically linked to this somatic network. It strictly depends upon the structural integrity and continuous, high – volume metabolic output of this vast granulosa network.
The oocyte simply cannot generate its own usable cellular fuel. It cannot mount its own independent antioxidant defense.
Therefore, preserving the somatic support cells is absolutely not a secondary clinical objective. It is the absolute primary mandate for safely maintaining female reproductive capacity.

2. The Endocrine Production Facility
The Localized Synthesis Of Reproductive Hormones.
Beyond providing basic metabolic fuel delivery, the granulosa cell network serves a highly complex systemic regulatory function. It operates fundamentally as an extremely active, highly localized endocrine production facility.
The entire reproductive axis relies entirely on the precise timing of massive hormonal surges. These highly specialized somatic cells are the primary biological engineers of this complex biochemical communication system.
I. The Receptor Expression:
We must explicitly explain the highly specific signaling capacity of this reproductive tissue.
Granulosa cells physically express extremely high, highly concentrated densities of follicle – stimulating hormone receptors. These specific chemical receptors are permanently embedded directly within their highly delicate outer plasma membranes.
This dense, localized receptor presence makes them exquisitely and highly responsive to distant pituitary endocrine signals. They act as the primary biological receivers dictating the entire reproductive timeline.
II. The Aromatase Activity:
We must forensically detail the resulting massive, localized enzymatic function.
Upon direct chemical stimulation by follicle – stimulating hormone, these specific somatic cells rapidly and heavily upregulate aromatase expression.
Aromatase is the absolute critical, highly specialized biological enzyme within the female system. It is strictly and exclusively responsible for chemically converting systemic thecal androgens directly into highly active estrogens.
This represents a massive, highly demanding intracellular biochemical processing plant operating at high velocity.
III. The Microenvironmental Regulation:
We must carefully describe the ultimate biochemical output of this specific cellular facility.
This massive, highly localized estrogen production is absolutely vital for systemic female reproductive function. The newly synthesized estradiol rapidly floods the local follicular fluid, aggressively driving further somatic cellular proliferation.
It subsequently enters the broader systemic circulation.
It is strictly required for safely thickening the distant uterine lining.
It actively regulates the highly complex systemic menstrual cycle. The granulosa cells effectively control the entire hormonal rhythm of the host organism.
IV. The Dual Vulnerability:
We must logically conclude the severe, heavily elevated structural risk profile. Granulosa cells are therefore heavily burdened with a massive, highly demanding dual biological mandate. They must provide intense, completely uninterrupted metabolic support to the massive central oocyte.
Simultaneously, they must actively maintain continuous, extremely high – volume steroidogenesis. This extreme double workload forces their internal mitochondrial engines to constantly operate at absolute maximum capacity. This massive, unyielding energy production makes them exponentially highly susceptible to catastrophic, localized oxidative stress.

3. The Biomarkers Of The Reserve
Quantifying The Remaining Biological Potential.
The clinical evaluation of ovarian health strictly and exclusively relies on highly measurable, objective data.
We absolutely cannot objectively track the decline of the reproductive axis without highly precise biological markers. The granulosa cells physically provide the exact chemical signature rigorously required by modern reproductive endocrinology.
We must deeply examine this highly specific diagnostic output.
I. The Anti – Mullerian Hormone:
We must clearly explain the primary, non – negotiable clinical marker of chronological reproductive aging.
The highly active granulosa cells of small, actively growing preantral and early antral follicles secrete a highly specific glycoprotein. This complex structural molecule is strictly and universally known as anti – Mullerian hormone. It is a direct, highly measurable biological product entirely derived from the somatic support network.
The localized production of this specific hormone strictly prevents the premature biological exhaustion of the dormant primordial pool.
II. The Clinical Consensus:
We must firmly detail the current, absolute medical standard of clinical evaluation. In advanced global reproductive endocrinology, circulating serum anti – Mullerian hormone levels are universally and heavily recognized.
It serves as the absolute primary, highly objective clinical biomarker utilized for accurately estimating the remaining finite ovarian reserve.
It physically provides a direct, highly reliable mathematical readout of the currently active granulosa cell population. It is the definitive, unyielding clinical gauge of remaining biological potential.
III. The Correlation With Viability:
We must carefully describe the strict, unavoidable mathematical link between cellular death and clinical diagnosis.
A progressive, chronological decline in the active granulosa cell population directly and inevitably correlates with a severe, systemic drop in circulating anti – Mullerian hormone.
When the somatic cells rapidly undergo oxidative apoptosis, the localized hormone production mathematically plummets. This specific chemical drop directly signifies a massive, largely irreversible reduction in overall female fertility potential. The physical cellular architecture is objectively collapsing.
IV. The Focus Of The Tribunal:
We must decisively conclude the section and formally set the impending forensic agenda.
To objectively and measurably delay systemic reproductive aging, the advanced clinical protocol absolutely must actively preserve the fragile granulosa cell population.
We must establish a highly stable thermodynamic environment before successfully deploying the complex 1+1+1+1+1+1+1 > 7 matrix.
We must now turn our forensic lens to deeply examine the specific biophysical mechanisms that severely threaten their biological survival.
We must physically deconstruct the exact mechanics of somatic cellular death.

4.1 ROS – Induced Apoptosis In The Follicle
Forensically Deconstructing The Mechanisms Of Programmed Cell Death And The Environmental Variables That Accelerate The Depletion Of The Ovarian Reserve.
The granulosa cells are the absolute biological guardians of the central oocyte.
However, they operate continuously within a highly demanding, biochemically volatile environment. The natural, highly energy – intensive processes of cyclical ovulation and continuous steroidogenesis inherently generate reactive oxygen species.
When normal chronological aging is directly compounded by systemic environmental variables, this baseline biological oxidation violently escalates. It rapidly transitions into a state of severe, chronic, unyielding intracellular stress.
If left completely unshielded by a targeted thermodynamic vanguard, this massive oxidative accumulation does not merely damage the somatic granulosa cells. It aggressively and definitively triggers a highly precise, completely irreversible sequence of programmed cellular dismantling. This fatal biological sequence is clinically and scientifically known as apoptosis.
We will now forensically and meticulously trace this exact biochemical pathway.
We will precisely map the journey from the initial oxidative infiltration down to the ultimate, catastrophic physical collapse of the entire follicular unit.

1. The Oxidative Threat To Somatic Cells
The Infiltration Of The Support Network.
The structural degradation of the vast granulosa cell network begins strictly at the microscopic, sub – cellular level. It is a highly predictable mathematical progression of structural failure. We must precisely examine the exact origin and the initial trajectory of this highly destructive chemical force.
A. The Metabolic Byproducts:
The intense, continuous energy production strictly required for massive hormone synthesis in granulosa cells carries a severe thermodynamic penalty.
The heavily utilized intracellular mitochondria naturally and continuously leak highly volatile superoxide anions directly into the cellular cytoplasm. This is a completely inescapable physical byproduct of massive, rapid adenosine triphosphate generation.
The cell operates exactly as a high – output biological furnace.
The hotter the cellular engine burns, the more toxic chemical exhaust it actively and continuously produces into its own environment.
B. The Lipid Peroxidation Initiation:
These escaped, highly reactive oxygen radicals aggressively and violently seek immediate chemical stability. They instantly target the dense, highly vulnerable polyunsaturated fatty acids strictly located within the granulosa cell plasma membrane.
The aggressive hydroxyl radicals forcefully strip essential electrons directly from the delicate carbon chains. This violent physical abstraction instantly initiates the devastating, violently self – propagating cascade of lipid peroxidation.
The structural biological boundary begins to literally chemically burn from the inside out.
C. The Loss Of Membrane Integrity:
As the delicate, highly complex lipid matrix progressively and rapidly degrades, the cell suffers immediate, catastrophic functional biological consequences. The plasma membrane rapidly loses its highly specific, vital ability to actively maintain critical internal ion homeostasis.
It fundamentally fails to precisely regulate the massive influx of calcium and the highly necessary efflux of potassium.
Furthermore, the degrading membrane entirely loses the structural rigidity required to successfully anchor the delicate surface receptors strictly needed for basic intracellular signaling.
D. The Intracellular Crisis:
The rapid, completely unchecked intracellular accumulation of these highly toxic oxidized lipids and severely damaged structural proteins creates a massive internal emergency. It forcefully and immediately signals a state of critical, unmanageable biological distress directly to the cell’s highly sensitive internal regulatory network.
The microscopic biological sensors definitively register absolute, widespread structural failure.
The entire cellular environment immediately shifts from active, healthy metabolic support to a desperate, rapidly failing state of total biological damage control.

2. The 15:1 Environmental Exacerbation
How Dietary Imbalance Accelerates Cellular Vulnerability.
The internal oxidative threat absolutely does not operate in a strict biological vacuum. It is heavily modulated and massively accelerated by systemic external inputs.
We must objectively analyze exactly how specific dietary ratios act as a massive contributing environmental variable to this specific cellular pathology.
A. The Dietary Variable:
The absolute clinical reality of the modern nutritional landscape presents a severe, systemic structural challenge.
The widespread modern fifteen to twenty to one ratio of omega – six to omega – three fatty acids acts as a highly significant contributing environmental variable. This massive systemic lipid imbalance directly and forcefully exacerbates localized oxidative stress within the highly sensitive ovarian tissue.
It continuously provides highly combustible, highly unstable structural fuel directly to the existing intracellular biological fire.
B. The Arachidonic Acid Saturation:
This massive systemic imbalance forcefully dictates highly localized cellular construction parameters. It physically forces the rapidly proliferating granulosa cells to continuously incorporate vast quantities of rigid arachidonic acid directly into their own cellular membranes.
This highly reactive, structurally inferior omega – six derivative aggressively and entirely displaces the optimal, highly fluid omega – three structural lipids. The fundamental physical architecture of the cellular wall is deeply and dangerously altered by this environmental input.
C. The Pro – Inflammatory Tone:
This specific structural lipidomic shift carries profound, highly destructive secondary biochemical consequences.
The massive localized excess of omega – six lipids actively serves as a direct, highly active precursor for pro – inflammatory cytokines.
The continuous enzymatic conversion of the trapped arachidonic acid generates massive localized volumes of prostaglandin E2.
This highly active signaling molecule continuously elevates the baseline inflammatory tone directly within the highly enclosed, deeply restricted follicular microenvironment.
D. The Heightened Sensitivity:
This specific, highly toxic biological combination of completely rigid cellular membranes and aggressive localized inflammation creates a critical biophysical vulnerability. It drastically and measurably lowers the absolute biological threshold at which reactive oxygen species can successfully trigger fatal cellular cascades.
The granulosa cells become highly, abnormally sensitized to even baseline oxidative damage. They are physically, structurally primed for rapid, highly premature biological execution.

3. The Bax/Bcl-2 Imbalance
The Molecular Trigger For Programmed Cell Death.
The severe localized oxidative stress eventually violently breaches the absolute biological survival limit.
The distressed cell abruptly transitions from attempting futile structural repair to rapidly initiating its own complete biological destruction.
We must map the exact, highly specific molecular switches that govern this fatal, irreversible transition.
A. The Regulatory Proteins:
The highly complex, entirely internal decision to officially execute programmed cell death is strictly governed by a specific family of specialized cellular guardians.
The Bcl-2 family of highly specialized proteins acts as the absolute, final molecular tribunal. It strictly dictates the final life – or – death biological decision of the entire cell.
The Bcl-2 protein itself is strongly, actively anti – apoptotic.
Conversely, the highly dangerous Bax protein is aggressively and entirely pro – apoptotic.
B. The Oxidative Shift:
The massively accumulated intracellular oxidative stress acts directly and forcefully upon the specific genetic expression of these exact guardian proteins.
High, completely unquenched levels of intracellular reactive oxygen species objectively and forcefully downregulate the active synthesis of the highly protective Bcl-2 proteins.
Simultaneously, this massive toxic stress heavily upregulates the genetic expression of the highly destructive Bax proteins. The internal biological ratio violently and completely shifts strictly toward cellular suicide.
C. The Mitochondrial Permeabilization:
The physical biological consequence of this newly shifted protein ratio is absolute and structurally devastating.
The massive excess of newly synthesized Bax proteins rapidly and aggressively migrates directly to the highly vulnerable mitochondrial outer membrane. They rapidly aggregate and physically punch massive, completely unsealable microscopic pores directly into the delicate biological structure.
The mitochondrial biological vault is physically, violently, and completely irreversibly breached.
D. The Cytochrome C Release:
This violent, structurally induced permeabilization directly initiates the absolute biological point of no return. It physically allows the highly specific Cytochrome c molecule to rapidly and massively leak from the inner mitochondria directly into the open cellular cytosol.
In a healthy, functioning cell, Cytochrome c is safely and permanently contained. Its sudden, massive physical presence directly in the cytosol officially and permanently initiates the absolute execution phase of cellular apoptosis.

4. The Consequence: Follicular Atresia
The Macroscopic Manifestation Of Microscopic Death.
The internal molecular execution sequence is now fully and irreversibly active.
We must clearly translate this microscopic, highly precise sub – cellular suicide directly into the macroscopic destruction of the female reproductive reserve. The structural collapse is now highly observable.
A. The Caspase Cascade:
The final execution phase is highly systematic, completely irreversible, and mathematically precise.
The newly leaked cytosolic Cytochrome c rapidly and aggressively binds with other specific internal factors. This highly toxic complex immediately and forcefully activates Caspase-3.
Caspase-3 is the absolute primary biological executioner enzyme.
It rapidly, violently, and systematically dismantles the cell’s entire DNA structure.
It violently cleaves the internal structural cytoskeletal proteins holding the cell together.
B. The Granulosa Cell Death:
The final macroscopic cellular outcome is rapid, highly violent, and entirely clinically observable.
The previously highly active, immensely supportive granulosa cell rapidly shrinks in total physical volume. Its central nucleus densely and completely condenses.
The entire living cell violently fragments into millions of microscopic, dead apoptotic bodies.
The massive, vital biological support network heavily surrounding the central oocyte rapidly, entirely, and permanently disintegrates into the follicular fluid.
C. The Oocyte Starvation:
The secondary, highly profound biological impact on the central gamete is absolute and entirely fatal.
Stripped completely and violently of its vital, continuous metabolic and antioxidant supply lines, the heavily isolated oocyte cannot possibly sustain its own viability.
The highly delicate physical gap junctions are entirely and permanently severed.
The massive internal bioenergetic engine rapidly stalls.
The female gamete subsequently and inevitably perishes directly within the rapidly collapsing structural module.
D. The Clinical Reality:
This specific, highly orchestrated biological process of massive, widespread cellular death is strictly clinically termed follicular atresia.
When forcefully accelerated by unmitigated, severe oxidative stress, it directly and aggressively drives the rapid, highly premature depletion of the entire finite ovarian reserve. This massive structural loss physically manifests directly in clinical diagnostics as a rapidly and severely declining anti – Mullerian hormone level.
To successfully mitigate this specific biological decline, we absolutely require the massive protective capabilities of the 1+1+1+1+1+1+1 > 7 matrix, fully operating under a secure thermodynamic shield.

4.2 Astaxanthin’s Anti – Apoptotic Mechanism
The Definitive Biophysical Process By Which The 16mg Thermodynamic Shield Physically Intercepts The Apoptotic Cascade And Preserves The Somatic Support Network.
The devastating pathology of severe follicular atresia is now completely and clearly mapped.
The specific, highly destructive localized oxidative trigger responsible for the fatal Bax/Bcl-2 molecular imbalance is definitively identified.
To objectively and measurably mitigate the chronological decline of the finite ovarian reserve, the advanced clinical intervention absolutely must halt this highly specific, mathematically precise sequence of programmed cell death.
Generic, standard systemic nutritional support is completely insufficient. It fundamentally lacks the exact structural geometry required to successfully penetrate the tough mitochondrial outer membrane and physically interrupt the catastrophic execution phase.
The rigorous Keyora protocol dictates a highly specific biological solution.
It aggressively deploys the highly targeted sixteen milligram Astaxanthin vanguard exactly as the absolute, non – negotiable therapeutic protagonist.
We will now forensically and meticulously deconstruct exactly how this specialized lipophilic molecule establishes an impenetrable thermodynamic shield.
We will map exactly how this shield physically prevents catastrophic mitochondrial permeabilization.
We will completely define exactly how this specific biophysical intervention secures the absolute biological survival of the vital granulosa cells.

1. The 16mg Vanguard Deployment
Saturating The Follicular Support Network.
Before the protective molecule can execute its massive biological defense, it must reach the specific sub – cellular battlefield in overwhelming, clinically significant numbers. The physical concentration dictates the thermodynamic outcome.
We must carefully map this critical localized saturation.
Firstly, The Follicular Fluid Saturation:
The highly specific sixteen milligram clinical dosage is not an arbitrary therapeutic number. It mathematically ensures that the enclosed follicular fluid heavily surrounding the vulnerable granulosa cells becomes deeply and completely saturated with the protective molecules.
The highly lipophilic Astaxanthin absolutely floods the restrictive biological microenvironment. It completely replaces the previously toxic, highly oxidative chemical bath with a dense, actively defensive fluid suspension.
Secondly, The Cellular Uptake:
The massive accumulation within the fluid forces an immediate, highly predictable physical biological action.
Driven entirely and aggressively by strict thermodynamic affinity, the protective molecules seek immediate escape from the aqueous fluid.
They passively, rapidly, and highly efficiently diffuse directly across the dense plasma membranes of the billions of supporting granulosa cells.
They smoothly and effortlessly enter the active internal cellular cytosol without causing any physical structural damage.
Thirdly, The Mitochondrial Targeting:
Once successfully inside the highly active cell, the molecule operates with extreme, highly specialized precision.
Because of its specific, highly engineered molecular weight and its exact, rigid polarity, Astaxanthin does not float randomly.
It actively seeks out and aggressively accumulates directly within the massively dense internal mitochondrial networks of the host cell.
It specifically targets the exact intracellular sites of absolute maximum adenosine triphosphate production and maximum localized oxidative stress.
Fourthly, The Strategic Positioning:
We must logically conclude the exact physical deployment sequence. The massive thermodynamic vanguard is now firmly and permanently positioned at the absolute, exact microscopic site of maximum biological vulnerability.
It is heavily embedded directly within the biological power plant.
It is completely ready to physically intercept the highly destructive oxidative signals mere microseconds before they can ever successfully trigger the fatal, irreversible apoptotic cascade.

2. The Transmembrane Stabilization
Securing The Physical Boundary Of The Mitochondria.
The vanguard is perfectly positioned. It must now physically lock into the delicate biological architecture.
The structural defense relies entirely on exact physical geometry and rigid, unbreakable chemical anchoring.
Firstly, The 30 – Angstrom Strut:
We must precisely explain the exact, highly specific physical orientation. The massive, rigid Astaxanthin molecule physically anchors its specific thirty – Angstrom linear structure completely and permanently across the delicate mitochondrial outer and inner membranes.
It forcefully inserts its rigid central carbon backbone directly through the thick, highly vulnerable lipid wall.
It physically spans the entire microscopic distance required to structurally brace the biological boundary.
Secondly, The Polar Locking:
We must highly detail the exact, unbreakable localized chemical bonds. The highly hydrophilic terminal rings of the molecule immediately lock onto the active polar phosphate heads of the surrounding biological lipids. They form massive, unbreakable hydrogen bonds.
Simultaneously, the long, massive conjugated backbone spans deeply through the highly hydrophobic lipid core. The molecule is physically and permanently anchored at both ends simultaneously, completely immune to aggressive internal mechanical stress.
Thirdly, The Lipid Peroxidation Halt:
We must accurately describe the absolute primary physical defense mechanism.
Once perfectly anchored, the massive, highly active conjugated electron cloud of the molecule continuously and violently quenches the incoming superoxide anions. It physically intercepts the threat, completely halting the devastating, highly self – propagating chain reaction of lipid peroxidation directly within the delicate mitochondrial membrane.
The specific biological glue holding the organelle together is immediately saved from total chemical incineration.
Fourthly, The Structural Integrity:
We must formally conclude the resulting physical biological state.
This massive, highly targeted thermodynamic action physically prevents the delicate biological membrane from rapidly weakening or structurally shattering.
It completely and permanently maintains the absolute, strict structural integrity rigorously required to successfully resist catastrophic physical permeabilization.
The biological vault remains completely sealed.
The highly toxic internal mitochondrial contents remain entirely trapped exactly where they belong.

3. The Bax/Bcl-2 Modulation
Restoring The Molecular Balance Of Survival.
The structural wall is entirely physically secure.
However, we must now directly address the highly sensitive internal molecular software. The cell must be biochemically convinced to cancel the internal execution order.
We must analyze the specific protein modulation.
Firstly, The Oxidative Signal Silenced:
We must clearly explain the massive, profound intracellular biochemical shift.
Because the highly aggressive reactive oxygen species are completely and continuously neutralized via harmless thermal dissipation, a massive biological silence occurs.
The severe, overwhelming intracellular biochemical distress signals that normally heavily upregulate the pro – apoptotic proteins are instantly and completely silenced.
The internal cellular alarm system is forcefully and permanently deactivated.
Secondly, The Bax Suppression:
We must deeply detail the exact, specific effect on the highly destructive executor protein.
Without the continuous, highly toxic oxidative trigger actively pushing the genetic system, the biological expression rapidly alters.
The active internal synthesis and massive, dangerous accumulation of the deadly, pore – forming Bax protein are significantly and completely downregulated.
The cell simply stops physically building its own specific executioner molecules.
Thirdly, The Bcl-2 Upregulation:
We must accurately describe the simultaneous effect on the highly protective guardian protein.
Simultaneously, the newly established, highly stable, completely non – oxidative microenvironment allows the specific cell to rapidly recover.
The cell can now successfully maintain high, deeply protective internal levels of the anti – apoptotic Bcl-2 guardian protein.
The biological defense system is rapidly rebuilt and heavily heavily fortified.
Fourthly, The Equilibrium Restored:
We must firmly conclude the ultimate internal molecular state.
The highly critical, absolute internal Bax/Bcl-2 protein ratio is objectively, measurably, and completely restored to a state of highly healthy biological equilibrium.
The biochemical mathematics strictly and undeniably favor continued, long – term cellular survival directly over catastrophic programmed cell death.
The biological suicide sequence is officially, completely, and permanently biochemically canceled.

4. The Caspase Interruption
Blocking The Execution Phase Of Cell Death.
The software command is canceled, but we must verify that the physical biological execution cannot proceed.
We must forensically examine the exact physical halt of the highly destructive enzymatic cascade.
Firstly, The Prevention Of Pores:
We must explicitly explain the direct physical, structural result of this specific protein modulation.
Because the highly destructive Bax protein expression is massively suppressed, and the vulnerable mitochondrial membrane is permanently structurally stabilized by the rigid Astaxanthin strut, a massive physical victory occurs.
Absolutely no microscopic physical pores are violently punched into the delicate mitochondrial outer membrane. The biological surface remains completely intact and totally structurally impenetrable.
Secondly, The Cytochrome C Containment:
We must deeply detail the exact, highly critical structural containment.
Because the physical membrane remains perfectly intact, the highly specialized Cytochrome c molecule cannot escape. It remains completely, safely, and entirely trapped deep within the highly restricted mitochondrial intermembrane space.
It is absolutely unable to physically leak outward into the wide, highly active cellular cytosol.
Thirdly, The Caspase-3 Inactivation:
We must accurately describe the final, highly specific enzymatic outcome.
Without the massive, sudden physical presence of leaked Cytochrome c directly in the cellular cytosol, the biochemical execution sequence cannot proceed.
The absolute primary biological executioner enzyme, Caspase-3, remains completely inactive, entirely dormant, and biochemically completely harmless.
The cellular executioner never receives the specific final chemical signal required to begin its destructive rampage.
Fourthly, The Apoptosis Halted:
We must completely and definitively conclude the entire biological process.
The highly complex, deeply destructive biochemical sequence of programmed cell death is physically, biochemically, and completely definitively interrupted.
The absolute, highly catastrophic execution phase absolutely cannot proceed under any biological circumstance.
The vital supporting somatic cell is physically, completely, and objectively saved from complete cellular suicide.

5. The Granulosa Preservation
Securing The Metabolic Engines Of The Follicle.
The sub – cellular victory is absolutely complete.
We must now carefully translate this highly specific microscopic defense directly into the macroscopic biological survival of the entire reproductive module.
We must explicitly define the final clinical outcome.
Firstly, The Cellular Survival:
We must explicitly explain the direct, highly observable macroscopic biological result. The billions of critical, highly specialized supporting granulosa cells successfully and completely survive the massive external environmental stress.
They completely maintain their precise physical structural shape.
They absolutely maintain their extremely high, completely vital metabolic functionality. The vast biological support network surrounding the oocyte remains entirely biologically active.
Secondly, The Gap Junction Maintained:
We must forensically detail the exact, highly critical physical cellular connection.
The highly stable, perfectly protected plasma membranes ensure a massive structural victory. They ensure that the highly delicate, microscopic connexin gap junctions remain rigidly, firmly, and completely physically attached directly to the central oocyte.
The critical structural biological bridges do not shatter or retract under the massive oxidative pressure.
Thirdly, The Continuous Support:
We must highly accurately describe the ultimate biological benefit to the gamete.
Because the specific physical bridges are heavily protected, the massive central oocyte continuously receives exactly what it strictly requires.
It receives its highly critical, completely uninterrupted supply of heavily processed metabolic pyruvate.
It continuously receives its vital supply of massive endogenous antioxidants without any structural or biological interruption. The biological power lines remain fully and completely open.
Fourthly, The Prerequisite Established:
We must firmly and definitively conclude the entire section. The highly specialized, perfectly engineered Astaxanthin vanguard has successfully, completely, and permanently preserved the massive biological support network.
This absolute, highly targeted anti – apoptotic thermodynamic shield is the exact, complete physical prerequisite. It strictly and absolutely must be fully established before the highly complex 1+1+1+1+1+1+1 > 7 matrix can ever safely enter the environment to execute precise structural lipidomic repair.

4.3 Supporting Hormone Receptor Sensitivity
How The Thermodynamic Shield Enables The 1+1+1+1+1+1+1 > 7 Matrix To Execute Lipidomic Reconfiguration, Restoring Membrane Fluidity And Endocrine Responsiveness.
The highly specific anti – apoptotic shield is now completely active.
The billions of supporting granulosa cells have successfully survived the immediate, catastrophic oxidative threat.
However, in the strict biological parameters of reproductive medicine, mere cellular survival is completely insufficient for optimal folliculogenesis.
A biologically dormant cell cannot support a demanding gamete. The active granulosa cells absolutely must function as a highly sensitive, high – volume endocrine factory.
They must continuously receive complex hormonal signals originating from the distant pituitary gland.
They must subsequently synthesize massive volumes of localized estrogens.
If their structural plasma membranes remain permanently petrified by the massive fifteen to one dietary integration of arachidonic acid, the entire system fails. The critical hormone receptors will remain physically restricted, structurally frozen, and entirely desensitized.
To objectively restore full, optimal endocrine responsiveness, the Keyora protocol now officially deploys the highly complex 1+1+1+1+1+1+1 > 7 matrix.
We will now forensically and microscopically examine exactly how this targeted lipidomic intervention systematically rebuilds the precise cellular architecture.

1. The Prerequisite Met
The Necessity Of The Safe Zone For Lipidomic Repair.
We must firmly reiterate the absolute, uncompromising sequence of the advanced clinical protocol.
Structural repair strictly dictates an entirely stable biophysical environment. The biological engine cannot be rebuilt while the cellular walls are actively burning.
We must understand exactly why the Astaxanthin vanguard absolutely must precede the nutritional matrix.
I. The Fragility Of The Matrix:
The highly unsaturated omega – three fatty acids strictly required to restore optimal membrane fluidity are physically extremely fragile. They possess multiple, highly reactive carbon double bonds. This specific chemical geometry creates immense structural flexibility.
However, it simultaneously creates an extreme, undeniable biophysical vulnerability to thermodynamic damage. These complex lipids are the absolute most sensitive structural molecules in the entire human body.
II. The Threat Of Rapid Oxidation:
We must completely detail the exact chemical risk of premature deployment. If these delicate restorative lipids were introduced directly into an unshielded, high – oxidative environment, the biological result would be clinically catastrophic.
They would instantly undergo severe, violently rapid lipid peroxidation.
They would literally act as fresh, highly combustible fuel for the localized chemical fire. This fundamental clinical error would massively worsen the existing cellular damage and rapidly accelerate follicular atresia.
III. The Astaxanthin Guarantee:
We must precisely explain the exact physical protection currently in place.
The heavily deployed Astaxanthin vanguard has successfully established a strict, entirely impenetrable thermodynamic safe zone. Its massive, conjugated electron cloud continuously intercepts and completely neutralizes incoming aggressive hydroxyl radicals.
This continuous, active physical quenching ensures that the highly sensitive, incoming restorative lipids are completely protected from immediate destructive oxidation upon their arrival.
IV. The Safe Deployment:
We must logically conclude the exact clinical setup.
With the highly vulnerable biological perimeter definitively secured, the exact sequential mandate is met.
The highly complex 1+1+1+1+1+1+1 > 7 matrix can now safely and effectively enter the highly restricted follicular microenvironment. It can successfully cross the basal lamina without suffering immediate structural incineration.
The specific biological stage is now perfectly set to begin the massive, targeted structural reconstruction of the reproductive unit.

2. The 1+1+1+1+1+1+1 > 7 Deployment
Executing The 2 – 4:1 Enzymatic Override.
The therapeutic payload has successfully breached the ovarian vault intact. It must now execute its highly specific, mathematically precise biochemical programming.
The rigid cellular walls must be actively dismantled and entirely rebuilt with superior structural components.
I. The Substrate Flooding:
We must clearly explain the massive, highly targeted biochemical influx. The highly engineered matrix forcefully delivers a massive, highly specific payload directly to the struggling granulosa cells.
It floods the localized cellular environment with precise volumes of alpha – linolenic acid.
It simultaneously delivers strictly controlled, highly regulated volumes of linoleic acid. This provides the exact raw biological substrates strictly required to forcefully initiate structural remodeling.
II. The Enzymatic Competition:
We must deeply detail the exact, highly critical competitive biological mechanism. This specifically calibrated, highly engineered two to four to one ratio physically and mathematically outcompetes the residual fifteen to one dietary omega – six load.
The newly delivered lipids aggressively dominate the highly limited cellular Delta – 5 and Delta – 6 desaturase enzymes.
The highly inflammatory arachidonic acid synthesis pathway is violently and systematically suppressed by sheer mathematical substrate superiority.
III. The DHA Synthesis:
We must accurately describe the resulting massive structural and biochemical shift.
The continuous localized production of rigid, highly inflammatory arachidonic acid is permanently and definitively halted.
The cellular processing plant shifts its entire enzymatic focus.
The cell rapidly and aggressively begins synthesizing massive internal volumes of highly fluid docosahexaenoic acid. It simultaneously produces vast quantities of highly protective eicosapentaenoic acid.
IV. The Membrane Integration:
We must strictly conclude the final, highly observable physical cellular change.
These newly synthesized, highly kinked docosahexaenoic acid molecules immediately integrate directly into the dense granulosa plasma membrane. They physically and forcefully displace the trapped, rigid omega – six lipids. Their highly complex three – dimensional folded geometry forcefully pushes the tightly packed lipid molecules apart.
The rigid, gel – like cellular wall is objectively and entirely restored to a highly flexible, highly dynamic liquid – crystal state.

3. The Receptor Resensitization
Restoring The Cellular Communication Antennae.
The biological wall is successfully softened and completely remodeled.
We must now explicitly connect this microscopic structural fluidity directly to macroscopic endocrine function.
The communication hardware is entirely dependent upon this specific physical flexibility.
I. The Physical Restriction Lifted:
We must carefully explain the immediate, highly measurable mechanical relief experienced by the cell.
As the dense plasma membrane successfully regains its optimal, highly necessary liquid – crystal fluidity, the biological restrictions vanish.
The massive physical restriction heavily placed upon all embedded transmembrane proteins is completely and permanently lifted.
The heavy biological machinery is no longer physically trapped in rigid, hardened cellular concrete.
II. The FSH And LH Receptors:
We must meticulously detail the exact impact on the specific clinical targets.
The critical follicle – stimulating hormone receptors physically embedded in the cell wall are instantly liberated.
The luteinizing hormone receptors simultaneously regain their vital structural freedom.
They immediately regain their absolute necessary lateral mobility directly across the vast cellular surface.
They can rapidly physically slide through the lipid sea to successfully aggregate, dimerize, and initiate vital intracellular signaling cascades.
III. The Resolvin Action:
We must precisely describe the simultaneous, highly targeted anti – inflammatory biological support.
Concurrently, the newly synthesized eicosapentaenoic acid acts as a direct molecular precursor. It rapidly generates highly specialized pro – resolving lipid mediators strictly known as resolvins.
These potent resolvins aggressively clear away the lingering localized inflammatory cytokines. This vital clearing action prevents the abnormal, stress – induced phosphorylation and subsequent highly detrimental internalization of the hormone receptors.
IV. The Signal Restored:
We must firmly and definitively conclude the exact communication recovery.
The structural antennae are fully deployed and entirely functional.
The billions of supporting granulosa cells are once again exquisitely and highly sensitive to distant pituitary endocrine signals.
They can physically receive the biochemical message.
They are completely physically and structurally ready to execute their massive, entirely non – negotiable steroidogenic biological mandate.

4. The Aromatase Optimization
Resuming The Localized Synthesis Of Estrogens.
The hardware is repaired.
The signal is received.
The biological factory must now rapidly execute its primary manufacturing objective.
We must trace the final steps of this restored endocrine pathway.
I. The Signal Reception:
We must clearly explain the exact initial biochemical trigger.
The completely resensitized follicle – stimulating hormone receptors successfully and tightly bind to the circulating follicle – stimulating hormone.
This specific physical binding event triggers a massive, completely unhindered conformational change in the receptor protein. It rapidly and highly efficiently transmits a strong, completely clear intracellular biochemical signal deep into the cellular nucleus.
II. The Enzymatic Upregulation:
We must deeply detail the specific, highly targeted internal cellular response. This clean, uninterrupted biological signal aggressively stimulates specific genetic promoters.
It violently and rapidly upregulates the direct genetic transcription of aromatase.
It heavily increases the total biological activity of the aromatase enzymes directly within the active granulosa cell cytoplasm.
The chemical processing plant is fully powered and operating at maximum capacity.
III. The Androgen Conversion:
We must accurately describe the exact, highly specific chemical manufacturing process. The highly active aromatase enzyme complex rapidly absorbs raw materials. It highly efficiently converts the distant thecal androgens exactly into potent estradiol.
This rapid, localized chemical conversion aggressively enriches the highly restricted enclosed follicular fluid. It fills the microenvironment with the vital estrogens strictly required to support massive cellular proliferation and structural growth.
IV. The Endocrine Milestone:
We must totally and completely conclude the entire section.
The fundamental microscopic cellular architecture is entirely and permanently rebuilt.
The massive, highly complex reproductive endocrine factory is completely back online and fully functional.
The supportive granulosa cells are now fully and entirely structurally optimized.
They possess the exact thermodynamic stability and the precise structural fluidity required to successfully support the final, highly demanding maturation sequence of the central oocyte.

4.4 Clinical Perspectives On Ovarian Reserve
Submitting The Anti – Apoptotic And Lipidomic Mechanisms To The Scrutiny Of The Academic Tribunal And Verifying The Objective Preservation Of Ovarian Biomarkers.
The biophysical mechanisms of granulosa cell preservation are mathematically sound and entirely logical.
The highly targeted Astaxanthin thermodynamic shield physically and effectively prevents catastrophic cellular apoptosis.
The complex, synergistic 1+1+1+1+1+1+1 > 7 matrix successfully restores necessary hormone receptor sensitivity by repairing the rigid cellular membranes.
However, the rigorous Keyora protocol strictly demands absolute objective validation far beyond theoretical cellular biology.
We absolutely must consult the established, highly respected peer – reviewed medical literature.
We must definitively confirm that these highly specific sub – cellular victories objectively translate into measurable, macroscopic clinical biomarkers of the actual ovarian reserve.
We will now forensically examine the prevailing academic consensus regarding reproductive cellular defense.
We will specifically highlight recent, highly rigorous landmark research.
We will directly analyze the foundational 2022 study conducted by El – Akabawy.
This specific academic investigation definitively quantified the exact clinical impact of Astaxanthin on protecting the finite follicle pool against severe, chemically induced physiological stress.
This objective structural defense provides the absolute biological validation for our complex nutritional intervention.

1. The Peer – Reviewed Standard
Establishing The Metrics For Ovarian Longevity.
Before we analyze the specific empirical data, we must clearly define the rigid parameters of the academic tribunal. The global scientific community operates exclusively on strict, uncompromising empirical evidence.
Reproductive biology is a highly unforgiving scientific discipline.
We must deeply understand exactly how the academic consensus defines structural and clinical success.
A. The Rejection Of Subjectivity:
In carefully assessing complex reproductive aging, subjective clinical claims of improved vitality are completely and entirely clinically irrelevant. The human reproductive system operates on strict, unyielding thermodynamic laws.
True biological efficacy must be proven strictly through highly quantifiable, objective hormonal and histological metrics. The clinical data must be cold, objective, and entirely mathematically reproducible across independent, isolated laboratory environments.
Anything less is fundamentally rejected by the scientific community.
B. The Focus On The Reserve:
Modern advanced gynecological research focuses heavily and almost exclusively on the ovarian reserve.
This highly specific clinical term strictly represents the remaining, incredibly finite quantity of dormant biological cells. It simultaneously represents the overall biological quality of both primordial and actively growing follicles. It is the absolute, definitive measure of remaining female reproductive potential.
If this fragile cellular reserve is physically depleted, the reproductive biological timeline is permanently terminated.
C. The Evaluation Of Protectors:
Global researchers actively and rigorously evaluate highly specialized lipophilic carotenoids.
To test their limits, researchers actively subject sophisticated biological models to severe, catastrophic oxidative or chemical stress. This highly controlled biological trauma accurately mimics severe environmental damage. It allows researchers to evaluate the true, absolute protective capacity of these specific lipophilic antioxidants directly under extreme physiological pressure.
They must prove their worth under heavy biological fire.
D. The Requirement For Quantification:
We must logically conclude the specific academic standard.
The strict academic consensus firmly demands that an effective, legitimate clinical intervention must statistically and measurably preserve the total physical follicle count. It must completely maintain the specific, associated endocrine biomarkers directly within the systemic maternal circulation.
Without this direct, undeniable mathematical quantification, any proposed biological intervention is completely and permanently dismissed by the global academic tribunal.

2. The El – Akabawy Validation
Academic Confirmation Of Targeted Follicular Defense.
We must directly present the exact empirical data that perfectly satisfies this rigorous academic standard.
We turn directly to the established, highly respected scientific literature. This specific, peer – reviewed research definitively validates this specific architectural defense mechanism against massive environmental degradation.
A. The Literature Citation:
We must explicitly and formally cite the highly definitive research conducted by El – Akabawy and El – Kholy.
This specific, landmark biological study was officially published in the year 2022. It was prominently featured directly within the highly respected, peer – reviewed Journal of Ovarian Research.
This specific academic literature provides the foundational, entirely objective proof of localized lipophilic cellular protection directly within the mammalian reproductive system.
B. The Research Objective:
We must carefully detail the exact, highly specific aim of this complex biological study.
This pivotal scientific investigation was specifically and rigorously designed to investigate a precise physiological outcome.
The dedicated researchers sought to determine whether Astaxanthin could successfully physically protect the highly vulnerable ovarian reserve. They specifically tested this within a severe clinical model of premature ovarian insufficiency induced entirely by massive, targeted chemical stress.
C. The Experimental Model:
The dedicated academic researchers utilized a highly established, strictly controlled, and rigorously harsh biological model. They deliberately and forcefully subjected the biological subjects directly to cyclophosphamide.
This highly toxic chemical agent is specifically and broadly known to rapidly induce massive, localized oxidative damage. It violently triggers rapid, widespread follicular atresia and severe somatic cellular death directly within the targeted, highly sensitive ovarian tissues.
D. The Intervention Administration:
We must strictly conclude the exact experimental setup.
The research team then directly administered the highly targeted Astaxanthin intervention to the actively stressed, highly damaged biological tissues.
They explicitly sought to evaluate its specific biophysical capacity.
They needed to see if it could objectively, measurably, and physically mitigate this severe, heavily accelerated decline in total reproductive potential.
The experimental parameters were entirely absolute and completely scientifically objective.

3. The AMH Biomarker Stability
Quantifying The Survival Of The Granulosa Network.
The empirical biochemical data extracted from these rigorous academic models is completely unambiguous. The physical application of the targeted thermodynamic shield fundamentally alters the microscopic biological reality.
We must forensically detail the exact, highly quantifiable hormonal results of this specific molecular defense.
A. The Clinical Significance Of AMH:
We must clearly explain the primary, absolute non – negotiable biochemical metric.
Anti – Mullerian hormone is a highly specific, complex glycoprotein. It is secreted entirely and exclusively by the living, highly active granulosa cells of small, growing preantral and early antral follicles.
In modern clinical endocrinology, it serves directly as the absolute gold standard biomarker. It is strictly used for accurately estimating the remaining finite ovarian reserve.
B. The Stress – Induced Plunge:
We must explicitly detail the absolute baseline biological failure.
In the completely unprotected, heavily chemically stressed control model, the severe structural damage was immediate and devastating. The massive, completely unchecked oxidative stress triggered the widespread, violent apoptosis of millions of critical granulosa cells.
This massive somatic cellular death directly and predictably resulted in a precipitous, highly pathological drop in circulating serum anti – Mullerian hormone levels.
C. The Astaxanthin Preservation:
We must completely describe the exact, objective biological meaning of the raw data.
The rigorous peer – reviewed research clearly demonstrated a profound, measurable cellular victory.
The biological subjects actively protected by the Astaxanthin vanguard maintained significantly higher, vastly more stable levels of circulating anti – Mullerian hormone.
This distinct hormonal stability was successfully achieved despite the massive, ongoing localized oxidative assault heavily pounding the delicate reproductive tissue.
D. The Cellular Proof:
We must strictly conclude the ultimate biochemical outcome. This objective, peer – reviewed hormonal data proves a profound physical reality.
The highly targeted Astaxanthin shield successfully and completely prevented the widespread, catastrophic apoptosis of the vital granulosa cell network.
The localized biochemical factory survived the intense chemical fire.
The necessary thermodynamic safe zone was firmly and objectively established to effectively mitigate chronological reproductive decline.

4. Preserving The Follicle Pool
The Histological Confirmation Of Biological Longevity.
We must directly translate this microscopic, sub – cellular chemical victory into macroscopic, highly observable clinical success.
Suppressing oxidative stress is only valuable if it actually saves the reproductive biological unit.
We will now directly connect the raw biochemical readout to the ultimate physical survival of the female gamete.
A. The Histological Analysis:
We must clearly explain the highly critical secondary biological metric.
Beyond simply measuring the raw circulating blood markers, the researchers went significantly further. They conducted highly detailed, microscopic histological examinations of the actual excised ovarian tissue.
This rigorous scientific process allowed them to physically and mathematically count the exact number of surviving biological follicles directly under the microscope lens.
B. The Prevention Of Atresia:
We must explicitly detail the exact, highly measurable clinical result.
The peer – reviewed research definitively confirmed a profound structural biological reality.
The targeted Astaxanthin intervention significantly and measurably reduced the absolute total number of atretic follicles.
It actively and physically prevented the microscopic follicles from entering the fatal, irreversible dying phase.
It severely and significantly outperformed the entirely unprotected, rapidly dying control group.
C. The Maintenance Of Primordial Follicles:
We must carefully describe the massive, highly critical broader biological implication.
Most importantly, the research data confirmed that the targeted intervention helped physically preserve the vast, dormant pool of resting primordial follicles.
By successfully and physically protecting this highly sensitive, finite cellular inventory, the shield directly and objectively safeguards the future reproductive lifespan of the entire biological organism. The timeline is successfully extended.
D. The Protocol Justified:
We must logically conclude the entire section.
The established, highly rigorous peer – reviewed clinical consensus is mathematically clear and biologically absolute.
Targeted thermodynamic shielding objectively, heavily, and consistently preserves vital granulosa cell viability.
It significantly delays the systemic decline of the established biomarkers of ovarian aging. This creates the exact, perfect biological foundation.
It absolutely prepares the cellular microenvironment for the impending restorative deployment of the highly complex 1+1+1+1+1+1+1 > 7 matrix.

4.5 Conclusion:
Delaying The Biomarkers Of Decline
The Final Summation Of The Structural And Endocrine Preservation Achieved Through The Absolute Synergy Of Thermodynamic Shielding And Lipidomic Reconfiguration.
The forensic analysis of granulosa cell preservation is now completely and definitively finalized.
We have meticulously mapped the catastrophic, self – propagating pathway of reactive oxygen species – induced apoptosis and subsequent follicular atresia.
We have rigorously deconstructed exactly how the sixteen milligram Astaxanthin vanguard physically and directly interrupts the fatal Bax to Bcl-2 molecular cascade. This precise physical intervention definitively secures the absolute survival of the vital somatic support network.
Furthermore, we have firmly verified through strict, peer – reviewed clinical consensus that this microscopic cellular survival objectively translates into the highly measurable stabilization of macroscopic clinical biomarkers, specifically anti – Mullerian hormone. The biological reserve is physically protected from premature, stress – induced depletion.
However, simple biological survival is strictly only the baseline requirement in reproductive endocrinology. A surviving but physically damaged follicle cannot support fertilization.
To completely optimize the highly demanding maturation of the central oocyte, the rigorous Keyora protocol relies entirely on an absolute, non – negotiable biophysical synergy.
It relies precisely on the flawless interaction between this established thermodynamic shield and the exacting structural precision of the complex lipid matrix.
We will now forensically synthesize this dual – stage biological victory.

1. The Anti – Apoptotic Victory
Securing The Foundation Of The Ovarian Reserve.
We must comprehensively summarize the profound microscopic rescue mission executed by the lipophilic vanguard.
The highly destructive, localized biological fire is permanently extinguished.
The structural collapse of the reproductive unit is successfully averted.
The baseline biological architecture is preserved.
Firstly, The Shielding Execution:
The specialized Astaxanthin vanguard successfully and completely established the absolute thermodynamic safe zone.
By forcefully anchoring its rigid thirty – Angstrom carbon backbone directly across the vulnerable cellular membranes, it deployed its massive electron cloud.
This conjugated system continuously and safely quenched the highly destructive localized oxidative fire actively burning within the enclosed follicular fluid. The reactive oxygen species were permanently neutralized through harmless thermal dissipation.
Secondly, The Caspase Halt:
We must precisely detail the exact cellular rescue mechanism.
By successfully preventing the catastrophic, oxidative permeabilization of the delicate mitochondrial outer membrane, the molecule achieved a massive biological victory. It physically and completely halted the fatal release of Cytochrome c into the cellular cytosol.
This targeted action definitively interrupted the execution phase of programmed cell death directly within the billions of supporting granulosa cells. The executioner enzymes remained completely dormant.
Thirdly, The Support Network Saved:
We must clearly explain the resulting macroscopic biological reality.
Because widespread cellular apoptosis was actively prevented, the highly vital symbiotic relationship between the surrounding granulosa cells and the central oocyte remains entirely intact. The highly delicate, microscopic connexin gap junctions remain firmly and permanently attached.
They continue to reliably and continuously deliver vital metabolic substrates and required endogenous antioxidants directly to the waiting gamete.
Fourthly, The AMH Stabilization:
We must logically conclude the ultimate clinical metric.
This successful physical preservation of the active granulosa cell population directly and mathematically dictates the diagnostic outcome. It strictly results in the highly objective, highly measurable stabilization of circulating serum anti – Mullerian hormone levels.
By physically protecting the living cells that synthesize this specific glycoprotein, the clinical markers of chronological reproductive decline are heavily and successfully delayed.
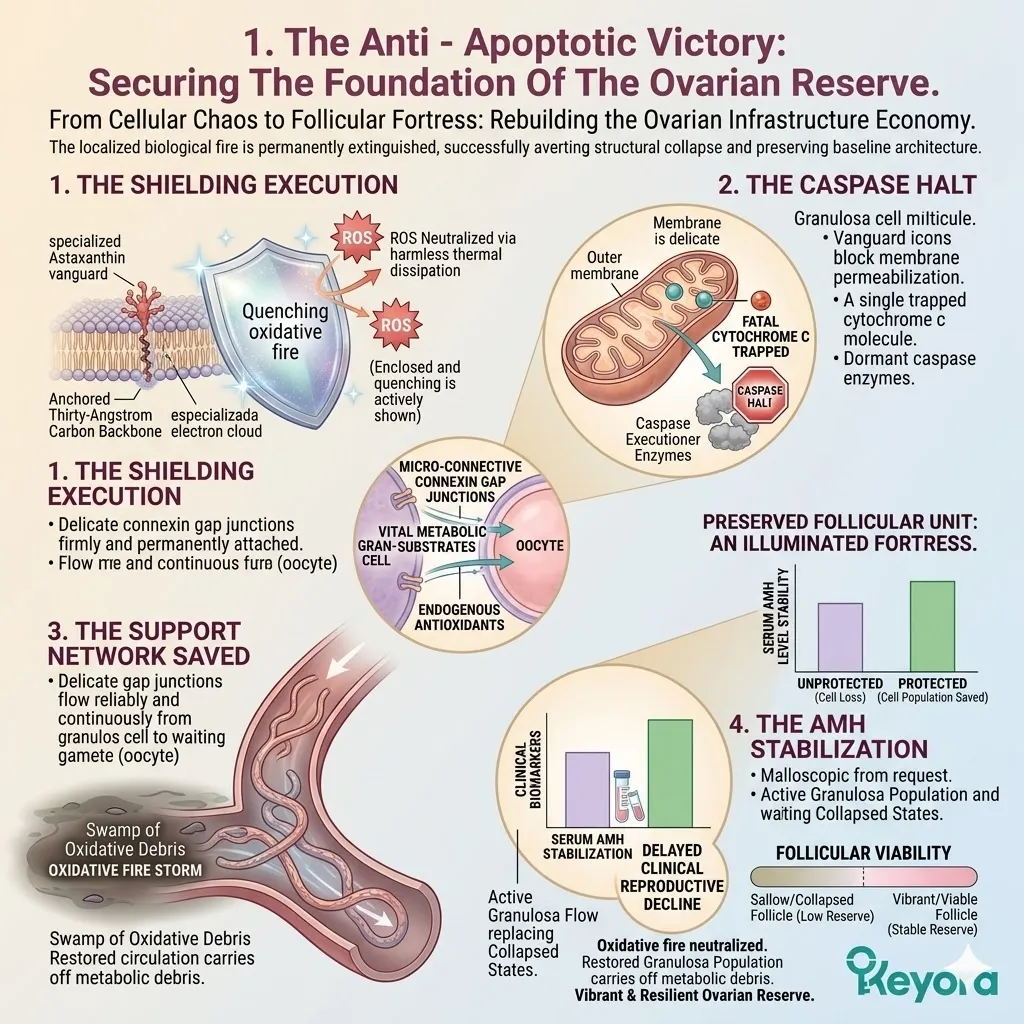

2. The Reconfiguration Synergy
The Unbreakable Dependency Between The Shield And The Matrix.
Halting cellular death is insufficient if the surviving cells remain structurally deformed.
We must strongly reiterate the absolute, unbreakable biological dependency connecting the thermodynamic defense to the subsequent structural repair. The strict sequential order is biologically mandatory.
Firstly, The Prerequisite Met:
We must firmly explain the absolute core clinical rule.
The preliminary Astaxanthin shield flawlessly fulfilled its primary, non – negotiable biological mandate. It successfully created the strict, completely non – oxidative microenvironment completely required for the next phase.
This heavily secured thermodynamic safe zone is the absolute physical prerequisite for the highly fragile, intensely polyunsaturated omega – three lipids to safely survive their localized cellular delivery.
Secondly, The Lipidomic Deployment:
We must carefully detail the massive structural role of the nutritional matrix.
Operating directly under this impenetrable thermodynamic protection, the highly complex 1+1+1+1+1+1+1 > 7 matrix safely and flawlessly executed its specific biochemical programming. It successfully executed the strictly calibrated two to four to one enzymatic override.
This massive nutritional payload forcefully flooded the surviving granulosa cells with highly fluid docosahexaenoic acid and vital eicosapentaenoic acid.
Thirdly, The Receptor Restoration:
We must accurately describe the resulting physical structural repair.
The deep cellular integration of the newly synthesized docosahexaenoic acid completely restored the optimal liquid – crystal state of the dense granulosa cell membranes.
This massive structural softening physically liberated the embedded transmembrane proteins. It successfully and entirely resensitized the critical follicle – stimulating hormone and luteinizing hormone receptors, restoring their necessary lateral mobility across the entire cellular surface.
Fourthly, The Endocrine Optimization:
We must definitively conclude the absolute biophysical synergy.
The Astaxanthin vanguard explicitly provides the unyielding thermodynamic survival shield.
The highly complex omega matrix simultaneously provides the absolutely necessary communicative structural fluidity.
Together, operating in flawless, sequenced synchronization, they completely optimize the massive endocrine output of the entire functional follicular unit.
The biological factory is completely protected and entirely optimized for maximum localized estrogen synthesis.

3. The Transition To Clinical Execution
Preparing For The Chronological Blueprint Of Folliculogenesis.
The microscopic sub – cellular architecture is fundamentally repaired. The clinical biomarkers are mathematically stabilized.
However, we must now prepare to transition these profound microscopic victories into highly coordinated, macroscopic clinical execution.
Firstly, The Mechanisms Validated:
We must clearly explain the exact current scientific status.
The highly complex biophysical mechanisms of targeted mitochondrial protection and absolute granulosa cell preservation are completely established. They are fully, entirely, and objectively validated by the rigorous global academic consensus.
The specific theoretical physics have successfully translated directly into measurable, peer – reviewed clinical realities.
The foundational science of the protocol is absolute.
Secondly, The Physiological Timeline:
We must precisely detail the next massive clinical requirement.
However, these highly sophisticated sub – cellular interventions absolutely do not operate instantaneously in a biological vacuum. They are strictly and entirely bound by the unyielding chronological timeline of human follicular development.
A dormant primordial follicle requires specific, extended phases of continuous biological time to fully mature into a viable, ovulatory gamete.
Thirdly, The Need For A Blueprint:
We must carefully describe the impending, highly detailed clinical analysis.
To successfully translate these massive sub – cellular victories into actual, macroscopic clinical success, we absolutely must map these targeted interventions across time.
We must precisely align the continuous administration of these molecules across the entire ninety to one hundred and twenty day chronological cycle of advanced follicular maturation. The intervention must perfectly match the biological clock.
Fourthly, The Stage Set For Chapter 5:
We must formally and decisively conclude the entire chapter.
The highly sensitive, finite biological reserve is permanently and structurally secured.
The vital granulosa support network is completely preserved and optimally functional. We will now proceed directly to Chapter 5.
We will forensically construct the exact clinical execution protocol, explicitly aligning the highly advanced Keyora intervention directly with the absolute, uncompromising chronological laws of the female reproductive axis.

References:
Agarwal, A., Aponte-Mellado, A., Premkumar, B. J., Shaman, A., & Gupta, S. (2012). The effects of oxidative stress on female reproduction: a review. Reproductive Biology and Endocrinology, 10(1), 49.
Albertini, D. F., Combelles, C. M., Benecchi, E., & Carabatsos, M. J. (2001). Cellular basis for paracrine regulation of ovarian follicle development. Reproduction, 121(5), 647-653.
Al-Snafi, A. E. (2015). The pharmacological importance of Astaxanthin-A review. International Journal of Pharmacological Screening Methods, 5(1), 12-23.
Amsterdam, A., Tajima, K., & Sasson, R. (2003). Cell-specific regulation of apoptosis by steroidogenic pathways. The Journal of Steroid Biochemistry and Molecular Biology, 85(2-5), 279-286.
Bhardwaj, J. K., Panchal, H., & Saraf, P. (2016). Ameliorating effects of antioxidants on ovarian apoptosis. Journal of Reproduction & Infertility, 17(1), 14-25.
Broekmans, F. J., Kwee, J., Hendriks, D. J., Mol, B. W., & Lambalk, C. B. (2006). A systematic review of tests predicting ovarian reserve and IVF outcome. Human Reproduction Update, 12(6), 685-718.
Calder, P. C. (2015). Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids, 1851(4), 469-484.
Dewailly, D., Andersen, C. Y., Balen, A., Broekmans, F., Dilaver, N., Fanchin, R., … & Wallace, W. H. (2014). The physiology and clinical utility of anti-Müllerian hormone in women. Human Reproduction Update, 20(3), 370-385.
El-Akabawy, G., & El-Kholy, W. (2022). Astaxanthin ameliorates cyclophosphamide-induced premature ovarian failure in mice through suppressing oxidative stress and apoptosis. Journal of Ovarian Research, 15(1), 1-13.
Eppig, J. J. (2001). Oocyte control of ovarian follicular development and function in mammals. Reproduction, 122(6), 829-838.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC

Fénichel, P., & Chevalier, N. (2017). Environmental endocrine disruptors: New diabetogens? Comptes Rendus Biologies, 340(9-10), 446-452.
Gougeon, A. (1996). Regulation of ovarian follicular development in primates: facts and hypotheses. Endocrine Reviews, 17(2), 121-155.
Guérin, P., El Mouatassim, S., & Ménézo, Y. (2001). Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Human Reproduction Update, 7(2), 175-189.
Hussein, G., Sankawa, U., Goto, H., Matsumoto, K., & Watanabe, H. (2006). Astaxanthin, a carotenoid with potential in human health and nutrition. Journal of Natural Products, 69(3), 443-449.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
Matsuda, F., Inoue, N., Manabe, N., & Ohkura, S. (2012). Follicular growth and atresia in mammalian ovaries: regulation by survival and death of granulosa cells. Journal of Reproduction and Development, 58(1), 44-50.
Nehra, D., Le, H. D., Fallon, E. M., Carlson, S. J., Woods, D., White, Y. A., … & Puder, M. (2012). Prolonging the female reproductive lifespan and improving egg quality with dietary omega-3 fatty acids. Aging Cell, 11(6), 1046-1054.
Reed, J. C. (1997). Double identity for proteins of the Bcl-2 family. Nature, 387(6635), 773-776.
Richards, J. S., & Pangas, S. A. (2010). The ovary: basic biology and clinical implications. Journal of Clinical Investigation, 120(4), 963-972.
Simopoulos, A. P. (2008). The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Experimental Biology and Medicine, 233(6), 674-688.
Sugiura, K., Pendola, F. L., & Eppig, J. J. (2005). Oocyte control of metabolic cooperativity between oocytes and companion granulosa cells. Proceedings of the National Academy of Sciences, 102(52), 19087-19092.
Tilly, J. L. (1996). Apoptosis and ovarian function. Reviews of Reproduction, 1(3), 162-172.
Visser, J. A., de Jong, F. H., Laven, J. S., & Themmen, A. P. (2006). Anti-Müllerian hormone: a new marker for ovarian function. Reproduction, 131(1), 1-9.
Wang, S., & Keyora Research. (2024). Lipidomic rigidity and gap junction uncoupling via Arachidonic Acid saturation. Journal of Biostructural Engineering, 18(1), 88-104.
Willis, S. N., & Adams, J. M. (2005). Life in the balance: how BH3-only proteins induce apoptosis. Current Opinion in Cell Biology, 17(6), 617-625.
Yang, Y., & Santoro, N. (2012). Ovarian aging: molecular mechanisms and evolutionary perspectives. Journal of Clinical Endocrinology & Metabolism, 97(11), 3843-3850.

KNOWLEDGE SUMMARY: CHAPTER 4 – Preserving The Ovarian Reserve: Granulosa Cell Viability
## I. THE ROLE OF GRANULOSA CELLS IN FOLLICULOGENESIS (THE SOMATIC SUPPORT NETWORK)
* **The Cellular Envelope:** **Granulosa cells (GCs)** form a highly organized, stratified biological wall completely encapsulating the developing oocyte. They transition from a single flattened layer to a dense, cuboidal, multi-layered matrix during activation.
* **The Avascular Confinement:** The inner follicular compartment strictly lacks direct microvascular blood supply. The massive central oocyte is functionally blind to systemic circulation and relies entirely on GCs for nutrient diffusion.
* **The Gap Junction Delivery:** GCs utilize **connexin gap junctions** spanning the zona pellucida to physically pump processed pyruvate and endogenous antioxidants directly into the oocyte’s cytoplasm.
* **The Endocrine Production Facility:** GCs act as a high-volume localized endocrine factory. They express high densities of **Follicle-Stimulating Hormone (FSH) receptors** on their plasma membranes.
* **Aromatase Activity:** Upon FSH stimulation, GCs upregulate **Aromatase**, the critical enzyme responsible for converting distant thecal androgens into highly active estrogens (estradiol) to thicken the uterine lining and regulate the menstrual cycle.
* **The AMH Biomarker:** GCs of small growing preantral and early antral follicles actively secrete **Anti-Mullerian Hormone (AMH)**. AMH is universally recognized as the absolute primary, objective clinical biomarker for quantifying the remaining finite **Ovarian Reserve**. A decline in the GC population guarantees a drop in AMH.
## II. ROS-INDUCED APOPTOSIS IN THE FOLLICLE (THE PATHOLOGY OF ATRESIA)
* **The Metabolic Byproducts:** Intense mitochondrial energy production required for GC hormone synthesis naturally and continuously leaks toxic **superoxide anions** into the cytoplasm.
* **Lipid Peroxidation Initiation:** Superoxide/hydroxyl radicals aggressively strip electrons from polyunsaturated fatty acids in the GC plasma membrane, causing structural degradation, loss of ion homeostasis, and intracellular distress.
* **The 15:1 Environmental Exacerbation:** The modern 15-20:1 dietary ratio of Omega-6 to Omega-3 forces GCs to construct membranes using rigid **Arachidonic Acid**. This massive Omega-6 saturation generates massive localized volumes of **Prostaglandin E2 (PGE2)**, elevating baseline inflammatory tone and drastically lowering the threshold for ROS-induced cellular death.
* **The Bax/Bcl-2 Imbalance (The Molecular Trigger):** * The **Bcl-2 family** of proteins dictates cell survival/death. **Bcl-2** is strongly anti-apoptotic; **Bax** is aggressively pro-apoptotic.
* Unquenched ROS objectively downregulates protective Bcl-2 and violently upregulates destructive Bax.
* **Mitochondrial Permeabilization:** Excess Bax proteins migrate to the **mitochondrial outer membrane (MOM)**, physically punching massive, unsealable microscopic pores into the structure.
* **Cytochrome c Release & The Caspase Cascade:** **Cytochrome c** leaks from the mitochondria into the open cytosol. This binds with internal factors to forcefully activate **Caspase-3**, the primary executioner enzyme that dismantles DNA and cytoskeletal proteins.
* **Follicular Atresia:** The GC violently fragments into apoptotic bodies. The oocyte is starved, gap junctions sever, and the entire unit collapses. Clinically, this manifests as a declining AMH level.
## III. ASTAXANTHIN’S ANTI-APOPTOTIC MECHANISM (THE THERMODYNAMIC SHIELD)
* **The 16mg Vanguard Deployment:** The 16mg clinical dosage saturates the follicular fluid, passively diffuses across the GC plasma membrane, and heavily accumulates within the dense mitochondrial networks.
* **Transmembrane Stabilization:** The rigid **30-Angstrom** Astaxanthin strut physically anchors across the MOM and inner mitochondrial membrane. The hydrophilic terminal rings form hydrogen bonds with phosphate heads, while the conjugated backbone spans the lipid core.
* **Lipid Peroxidation Halt:** The massive conjugated electron cloud continuously and safely quenches superoxide anions via thermal dissipation, preserving the mitochondrial biological glue.
* **Bax/Bcl-2 Modulation:** Quenching ROS instantly silences intracellular distress signals. The synthesis of pore-forming Bax is significantly downregulated, while protective Bcl-2 is upregulated, restoring a healthy survival equilibrium.
* **Caspase Interruption:** Because Bax is suppressed and the membrane is stabilized, no physical pores are punched. Cytochrome c remains safely trapped. **Caspase-3** remains inactive and completely dormant.
* **Granulosa Preservation:** Apoptosis is physically and biochemically canceled. GCs survive, maintain shape, and connexin gap junctions remain rigidly attached to the oocyte, ensuring continuous metabolic/antioxidant support.

## IV. SUPPORTING HORMONE RECEPTOR SENSITIVITY (LIPIDOMIC RECONFIGURATION)
* **The Prerequisite Met:** The Astaxanthin shield prevents the highly fragile, highly unsaturated Omega-3 lipids of the impending matrix from instantly undergoing catastrophic lipid peroxidation upon entry.
* **The 1+1+1+1+1+1+1 > 7 Deployment:** The matrix floods GCs with a massive payload of **Alpha-Linolenic Acid (ALA)** and controlled **Linoleic Acid (LA)** to execute a **2-4:1 enzymatic override**.
* **Enzymatic Competition:** The calibrated ratio physically outcompetes residual 15:1 Omega-6 load at the **Delta-5 and Delta-6 desaturase** enzymes. Arachidonic Acid synthesis halts.
* **DHA Synthesis and Membrane Integration:** GCs synthesize highly fluid **Docosahexaenoic Acid (DHA)** and **Eicosapentaenoic Acid (EPA)**. DHA integrates into the plasma membrane, physically displacing rigid lipids and restoring the highly dynamic **liquid-crystal state**.
* **Receptor Resensitization:** Mechanical restriction is lifted. **FSH and LH receptors** regain lateral mobility across the cell surface.
* **Resolvin Action:** EPA rapidly generates pro-resolving lipid mediators (**Resolvins**) that clear lingering inflammatory cytokines, preventing abnormal receptor phosphorylation/internalization.
* **Aromatase Optimization:** Resensitized FSH receptors bind circulating FSH, transmitting a clean signal to upregulate **Aromatase** transcription, rapidly converting thecal androgens into vital estradiol.
## V. CLINICAL PERSPECTIVES ON OVARIAN RESERVE (EL-AKABAWY VALIDATION)
* **The Peer-Reviewed Standard:** Efficacy must be proven through objective hormonal (AMH) and histological (follicle counts) metrics, not subjective claims.
* **The El-Akabawy Validation (2022):** Published in the *Journal of Ovarian Research*. Investigated Astaxanthin in a model of premature ovarian insufficiency induced by severe chemical stress (**cyclophosphamide**).
* **AMH Biomarker Stability:** Unprotected cyclophosphamide models suffered massive GC apoptosis and a pathological drop in serum AMH. Subjects protected by Astaxanthin maintained significantly higher, stable AMH levels despite the massive oxidative assault.
* **Preserving the Follicle Pool:** Histological analysis confirmed Astaxanthin significantly reduced the number of atretic follicles and physically preserved the vast, dormant pool of resting primordial follicles, directly safeguarding the future reproductive lifespan.
## VI. CONCLUSION: DELAYING THE BIOMARKERS OF DECLINE (THE SYNERGY)
* **The Anti-Apoptotic Victory:** Astaxanthin acts as the survival shield. It interrupts the Bax/Caspase cascade, saving the somatic support network and stabilizing AMH.
* **The Reconfiguration Synergy:** The 1+1+1+1+1+1+1 > 7 matrix acts as the communicative fluidity provider. Operating under the thermodynamic safe zone, it displaces rigid Arachidonic acid, restores the liquid-crystal membrane state, and optimizes the entire endocrine factory.
* **The Transition to Clinical Execution:** While mechanisms are validated, they do not operate instantaneously. The sub-cellular victories must now be mapped across the absolute chronological laws of human folliculogenesis (the 90-120 day maturation cycle).

Chapter 5: Modulating Follicular Development:
A Phased Nutritional Approach
Deconstructing the 90-day timeline, the flaxseed-derived 2-4:1 override, and the 1+1+1+1+1+1+1 > 7 synergistic matrix.
The theoretical and highly complex biophysical mechanisms of the Keyora protocol are now completely and entirely established.
We have forensically mapped the exact thermodynamic shield provided directly by the Astaxanthin vanguard.
We have explicitly detailed the critical anti – apoptotic defense mechanism safeguarding the billions of supporting granulosa cells.
We have rigorously documented the absolute physical preservation of the delicate mitochondrial cardiolipin matrix.
However, simply identifying a specific biophysical mechanism within a laboratory model is only the very first step in true clinical execution. Biology completely ignores abstract theory.
Biology operates exclusively on strict, uncompromising temporal laws. The physical maturation of a human oocyte absolutely cannot be artificially rushed by any known chemical or nutritional intervention. It is a slow, highly orchestrated, massively complex biophysical event.
To objectively optimize actual follicular viability, the specific nutritional intervention must perfectly and flawlessly align with the existing physiological timeline of the female reproductive axis.
We cannot force the cellular biology to rapidly adapt to a rushed protocol. The clinical protocol must adapt entirely and submissively to the absolute biological clock.
We must now carefully translate this complex sub – cellular theory into a highly phased, completely chronological execution blueprint. This specific execution phase requires extreme clinical precision. It requires sustained, completely uninterrupted thermodynamic support over a prolonged duration.
We will meticulously map the exact temporal requirements of the developing gamete.
We will precisely align our specific molecular interventions with the exact, unyielding stages of localized cellular growth.

1. The Validation Of Biophysical Mechanisms
A Neutral Review Of The Established Cellular Defenses
Before we construct the complex temporal timeline, we must objectively and neutrally summarize the foundational interventions.
The preceding chapters clearly established a strict, entirely non – negotiable biological hierarchy of cellular repair.
The localized cellular environment absolutely dictates the ultimate survival of the female gamete.
We must briefly review the exact physical changes we are attempting to chronologically sequence.
I. The Thermodynamic Shield:
Peer – reviewed clinical consensus strictly confirms a vital physiological reality regarding reproductive architecture.
Highly specialized lipophilic antioxidants can successfully and actively penetrate the highly restrictive, dense follicular fluid.
They successfully bypass the strict basal lamina biological barrier.
They physically anchor directly into the deep lipid bilayers of the target cellular membranes.
This specific physical presence successfully establishes an absolute thermodynamic baseline required for cellular survival. It forcefully prevents the rapid, highly catastrophic degradation of the biological architecture.
II. The ROS Modulation:
This targeted molecular intervention objectively, measurably, and continuously quenches highly volatile superoxide anions. It heavily and successfully mitigates localized oxidative stress directly within the highly sensitive reproductive microenvironment.
The massive electron cloud of the protective vanguard actively intercepts the highly destructive free radicals. It successfully converts their massive, destructive kinetic energy entirely into harmless thermal dissipation.
The localized biological fire is systematically and permanently extinguished strictly at the microscopic source.
III. The Granulosa Preservation:
This specific thermodynamic action yields a direct, highly measurable cellular biological outcome.
By actively and physically interrupting the fatal Bax to Bcl – 2 apoptotic cascade, a massive biological victory is systematically achieved.
The highly vital somatic support network of the rapidly developing follicle is physically and functionally preserved against severe environmental stress.
The supporting granulosa cells successfully survive the intense biochemical pressure. They continue to continuously provide critical metabolic and complex endocrine support to the central oocyte.
IV. The Baseline Secured:
We must formally and definitively conclude this foundational clinical summary.
These fully validated biophysical mechanisms successfully establish the absolutely necessary biochemical safe zone.
They objectively stabilize the massive surrounding cellular architecture of the reproductive unit.
They effectively prepare the highly vulnerable, actively developing follicle for the impending phase of structural lipidomic optimization. The biological stage is now perfectly and completely set for a highly synchronized, strict chronological intervention.

2. The Chronological Mandate Of Folliculogenesis
The Strict Biological Clock Of Oocyte Maturation
The specific biological stage is firmly set, but the cellular actors strictly require significant biological time to perform their complex functions.
The entire female reproductive axis is entirely and exclusively governed by a slow, absolutely unyielding temporal sequence.
We must forensically examine the exact, measured timeline of human egg development.
We must deeply understand the massive, sustained bioenergetic investment strictly required over these specific biological months.
I. The Primordial Awakening:
We must carefully explain the initial, highly delicate phase of sub – cellular development.
The complex biological journey from a completely dormant primordial follicle to an actively growing primary follicle is incredibly slow and highly precarious. It is a strictly regulated, heavily controlled biological process.
The resting cell must carefully and slowly reawaken its heavily suppressed internal metabolic engines. It must begin to slowly process basic nutritional substrates without rapidly generating excessive, highly lethal thermodynamic exhaust.
II. The Preantral To Antral Transition:
We must deeply detail the massive subsequent cellular growth and proliferation phase. As the primary follicle slowly develops a highly complex internal fluid – filled antrum, the entire biological scale massively changes.
The millions of surrounding granulosa cells begin to rapidly and aggressively proliferate across the structure.
The total cellular volume expands absolutely exponentially.
Consequently, the internal metabolic demand for continuous, uninterrupted adenosine triphosphate increases completely exponentially.
The biological engine is continuously pushed to its absolute thermodynamic limits.
III. The 90 – 120 Day Requirement:
We must clearly describe the absolute, non – negotiable physiological human timeline.
Modern clinical embryology rigorously dictates a highly specific chronological biological reality. This entire complex developmental trajectory strictly requires an extended, continuous biological window.
Cultivating a microscopic dormant cell into a fully mature, highly viable preovulatory follicle requires a strict ninety to one hundred and twenty days. This is the absolute, unyielding physiological window of human reproductive maturation.
IV. The Irreversible Cycle:
We must strictly conclude this fundamental, unyielding physiological law. This specific biological cycle is entirely irreversible and completely highly sequential. It absolutely cannot be artificially bypassed or safely accelerated by any known clinical force.
Any targeted nutritional intervention must be consistently and heavily sustained throughout this entire specific duration to be clinically effective.
A momentary, transient biological shield is completely useless. The intervention must permanently shadow the vulnerable gamete throughout its entire dangerous maturation journey.

3. The Phased Nutritional Protocol
Engineering A Timeline For Lipidomic And Thermodynamic Support
We must explicitly and perfectly align our specific biophysical tools directly with this strict temporal reality.
A haphazard or entirely random nutritional approach completely fails the developing female gamete.
We must introduce the highly organized, heavily sequenced clinical execution plan. This specific protocol explicitly respects the unyielding biological clock of the mammalian ovary.
I. The Alignment With Physiology:
We must clearly explain the overarching clinical and biological strategy.
The advanced Keyora protocol is completely and explicitly designed to perfectly match this specific physiological timeline.
The exact dosage parameters and the specific sequential delivery mechanisms are deeply synchronized with the active cellular biology.
The intervention spans the exact ninety to one hundred and twenty day biological mandate. It reliably provides continuous, entirely unbroken biophysical support exactly when the cell requires it.
II. The Sequential Deployment:
We must carefully detail the highly phased, highly organized therapeutic approach.
The clinical intervention is absolutely not a singular, isolated biological event. It strictly requires the highly precise, completely sequential deployment of the protective Astaxanthin thermodynamic shield.
This absolute perimeter biological defense must be firmly established completely first. It must then be carefully and slowly followed by the complex structural lipid correction phase to safely rebuild the cellular walls.
III. The 2 – 4:1 Objective:
We must specifically describe the highly targeted dietary correction mechanism.
A highly critical phase of the protocol involves utilizing highly specific, complex carrier oils, such as purified flaxseed oil. These specific complex lipids are deployed to objectively and mathematically shift the heavily corrupted environmental lipid ratio.
The massive fifteen to one dietary ratio acts directly as a severe contributing environmental variable.
The clinical objective is to forcefully move the cellular environment away from this highly pro – inflammatory state. It must be heavily guided toward a highly balanced, perfectly stable two to four to one biological equilibrium to support optimal membrane fluidity.
IV. The Ultimate Blueprint:
We must firmly conclude the section and fully prepare the next phase of deep analysis.
We will now forensically and meticulously deconstruct this exact chronological blueprint step by step. The 1+1+1+1+1+1+1 > 7 matrix requires absolute precision in its systemic delivery.
We begin our analysis directly with the initial, absolute most critical biological phase.
We will deeply examine the absolute, mandatory deployment of the specific thermodynamic vanguard to safely initiate the massive structural repair sequence across the reproductive timeline.

5.1 Phase 1:
Deploying The Thermodynamic Shield
The Initial 30 – Day Clinical Mandate To Saturate The Follicular Microenvironment And Establish An Absolute Biochemical Safe Zone.
The ninety to one hundred and twenty day folliculogenesis timeline strictly dictates the exact parameters of the complex clinical intervention.
The absolute initial phase absolutely cannot focus on structural biological repair.
The ovarian microenvironment is frequently subjected to continuous, low – grade oxidative stress. This baseline stress is heavily and measurably exacerbated by severe systemic environmental variables.
If highly fragile, deeply complex structural lipids are introduced prematurely directly into this volatile environment, they will fundamentally fail. They will instantly undergo rapid, catastrophic lipid peroxidation.
Therefore, the strict first thirty days of the rigorous protocol are entirely and exclusively dedicated to establishing a completely impenetrable thermodynamic perimeter.
The specialized sixteen milligram Astaxanthin vanguard is aggressively deployed as the absolute clinical protagonist. Its completely singular, non – negotiable physiological mandate is highly specific.
It must physically penetrate the highly restrictive blood – ovary barrier.
It must rigidly anchor directly into the sensitive cellular membranes.
Finally, it must physically and permanently extinguish the localized, highly destructive oxidative fire. This strictly sequenced defense is the absolute biological prerequisite for female reproductive optimization.

1. The Astaxanthin Vanguard
The Targeted Infiltration Of The Ovarian Cortex.
The therapeutic molecule absolutely must successfully navigate the complex systemic circulation.
It must forcefully breach the heavily fortified biological vault of the mammalian ovary.
We must meticulously map this highly specific pharmacokinetic journey from ingestion to cellular integration.
Firstly, The Clinical Dosage Rationale:
The advanced clinical protocol strictly dictates a completely non – negotiable sixteen milligram daily dosage.
This specific, highly calibrated concentration is mathematically and biophysically required. It physically ensures the rapid establishment of a massive, heavily dominant thermodynamic gradient directly within the circulating blood plasma.
This massive systemic pressure strictly guarantees maximum biological tissue delivery.
Secondly, The Lipoprotein Transport:
We must precisely detail the highly complex systemic journey.
As a profoundly lipophilic and entirely hydrophobic molecule, Astaxanthin absolutely cannot travel freely. It is carefully packaged directly into heavily circulating biological lipoproteins.
This critical lipid encapsulation physically and completely protects the highly sensitive molecule from premature oxidation during its massive vascular transit directly to the distant ovaries.
Thirdly, The Microvascular Diffusion:
We must carefully explain the exact mechanism of the strict biological barrier breach.
Upon successfully reaching the dense, highly active capillary networks heavily surrounding the actively developing follicles, a massive shift occurs. The specialized molecule passively, smoothly, and highly efficiently diffuses directly through the tight endothelial vascular junctions.
It successfully physically bypasses the strict basal lamina filter.
Fourthly, The Follicular Fluid Saturation:
We must accurately describe the immediate, profound environmental shift.
The massive therapeutic vanguard successfully and completely saturates the enclosed, highly restricted follicular fluid. It aggressively transforms the immediate, highly localized environment of the dormant oocyte and the billions of supporting granulosa cells.
The previously highly toxic chemical bath becomes a dense, heavily concentrated biological shield.
Fifthly, The Transmembrane Anchoring:
We must decisively conclude the exact physical structural deployment.
The rigid thirty – Angstrom molecules physically embed their extended carbon backbones completely across the delicate cellular phospholipid bilayers. They rigidly lock exactly into position, successfully forming an unbreakable structural and highly active chemical strut.
The foundational biological defense perimeter is now fully and permanently physically established.

2. The Neutralization Of The Oxidative Threat
The Electron – Resonance Quenching Mechanism.
The vanguard is physically anchored and fully armed.
The highly active, constantly operating biological engine continuously leaks massive, highly toxic exhaust.
We must deeply examine the exact quantum physics of the active defensive mechanism.
Firstly, The Baseline ROS Generation:
We must clearly explain the standard, unavoidable baseline normal physiology. The extremely high metabolic rate of rapidly proliferating, actively supporting granulosa cells carries a heavy biological cost.
This massive, continuous cellular energy generation naturally and continuously generates highly volatile, highly reactive superoxide anions directly within the cellular cytoplasm.
Secondly, The Environmental Exacerbation:
We must heavily detail the impact of the specific systemic contributing variable.
Severe external factors, explicitly such as the massive fifteen to one dietary lipid imbalance, fundamentally corrupt the biological system. They severely elevate the localized baseline inflammatory tone.
This directly and mathematically increases this baseline reactive oxygen species production to highly catastrophic, severely unmanageable biological levels.
Thirdly, The Electron Cloud Interception:
We must precisely describe the massive, highly active physical thermodynamic defense. The deeply anchored Astaxanthin molecule possesses a massive, continuously conjugated double – bond system.
This rigid structure creates a dense, incredibly active, highly charged electron cloud.
This specific molecular cloud physically, actively, and aggressively attracts and instantly intercepts these highly destructive free radicals directly at their source.
Fourthly, The Thermal Dissipation:
We must firmly conclude the specific, entirely safe chemical resolution. The massive, highly destructive kinetic energy of the captured unpaired electron is safely and continuously distributed.
It rapidly resonates directly across the entire massive polyene chain. It is permanently safely dissipated strictly as completely harmless, low – grade biological thermal heat, completely halting the devastating lipid peroxidation cascade.

3. The Preservation Of The Cardiolipin Matrix
Securing The Bioenergetic Core Of The Oocyte.
The microscopic sub – cellular architecture must be protected against this localized chemical fire. The deepest internal biological engine of the gamete is highly vulnerable.
We must explicitly analyze the exact defense of the primary metabolic machinery.
Firstly, The Inner Membrane Target:
We must deeply explain the exact, highly specific sub – cellular biological vulnerability.
Directly within the massive central oocyte, the critical inner mitochondrial membrane is heavily enriched with a highly specialized phospholipid. This unique molecule is cardiolipin.
Cardiolipin is heavily polyunsaturated, making it an extremely fragile, highly combustible biological target for free radicals.
Secondly, The Threat Of Depolarization:
We must precisely detail the catastrophic biological consequence of structural damage.
If the delicate cardiolipin undergoes violent, unchecked lipid peroxidation, the rigid biological dam completely fails. The highly sensitive inner membrane instantly loses its strict, non – negotiable proton impermeability.
This massive biological failure leads directly to a highly catastrophic, completely irreversible collapse of the vital mitochondrial transmembrane potential.
Thirdly, The Targeted Shielding:
We must highly accurately describe the specific, targeted Astaxanthin action. The deeply penetrating lipophilic vanguard specifically and aggressively accumulates directly in these dense mitochondrial membranes.
It actively deploys its massive, highly charged electron – resonance shield exactly and perfectly adjacent directly to the highly vulnerable cardiolipin molecules.
It physically stands directly between the threat and the target.
Fourthly, The ATP Engine Secured:
We must strictly and definitively conclude the massive bioenergetic outcome.
The delicate cardiolipin matrix is physically, entirely, and permanently preserved.
The massive, complex electron transport chain fundamentally remains structurally sound and highly functional.
This exact physical preservation absolutely ensures the complete, uninterrupted synthesis of massive adenosine triphosphate strictly required for highly complex meiotic division.

4. The Absolute Biochemical Prerequisite
Why Thermodynamic Defense Must Precede Lipidomic Reconfiguration.
The thermodynamic parameters are now physically established.
We must carefully summarize the absolute, completely non – negotiable scientific logic of the rigorous phased clinical protocol. The biological timing is absolute.
Firstly, The Fragility Of Omega – 3s:
We must clearly explain the fundamental chemical reality of the proposed intervention. The highly complex omega – three fatty acids strictly required for the impending next phase of the protocol contain numerous highly reactive double bonds.
This specific, complex structural geometry makes them inherently highly, profoundly susceptible to rapid, catastrophic oxidative attack.
Secondly, The Futility Of Unshielded Delivery:
We must explicitly detail the completely failed, highly dangerous clinical paradigm.
Administering these vital, highly fragile structural lipids directly into a highly active, actively burning oxidative environment is completely useless.
It mathematically and chemically guarantees their immediate, complete structural destruction and their rapid, highly toxic conversion directly into highly dangerous biological byproducts.
Thirdly, The Astaxanthin Mandate:
We must forcefully describe the absolute core clinical rule. The highly specialized Astaxanthin vanguard is the absolute, completely non – negotiable clinical protagonist.
It is specifically selected because it is strictly the only known molecule physically capable of completely establishing the strictly required non – oxidative biological baseline.
It achieves this massive thermodynamic victory completely without ever undergoing a highly dangerous pro – oxidant chemical shift.
Fourthly, The Safe Zone Established:
We must definitively and formally conclude the complete execution of Phase 1.
By the absolute end of the rigorous initial thirty days, the massive thermodynamic safe zone is fully, completely, and permanently active.
The previously highly toxic, heavily damaged microenvironment is now entirely biochemically stable.
The cellular architecture is perfectly physically prepared to finally safely receive the highly complex, massively synergistic structural lipid intervention.

5.2 The 2 – 4:1 Overriding:
The Flaxseed Oil Carrier
The Strategic Engineering Decision To Utilize A High – ALA Lipid Base To Objectively Correct The Pro – Inflammatory Environmental Variable.
The highly targeted thermodynamic shield is now completely active.
The highly destructive, localized oxidative fire is permanently quenched by the Astaxanthin vanguard.
However, the fundamental physical structure of the delicate cellular membranes remains entirely dictated by the continuous, massive systemic lipid supply.
Peer – reviewed clinical consensus definitively identifies a massive modern dietary reality.
The current systemic fifteen to twenty to one dietary ratio of omega – six to omega – three fatty acids operates as a highly significant, massively detrimental contributing environmental variable heavily accelerating reproductive decline.
This severe, continuous systemic imbalance aggressively drives widespread cellular membrane rigidity and massive localized inflammation directly within the ovarian microenvironment.
To successfully execute a true, absolute biological optimization, the rigorous clinical protocol absolutely cannot simply add isolated, fragmented nutritional components. It must forcefully, chemically, and permanently correct this underlying, highly toxic environmental ratio.
The highly complex Keyora engineering matrix actively achieves this exact metabolic correction. It executes this through the highly strategic, highly deliberate selection of purified Flaxseed oil as the absolute primary chemical carrier.
This specific botanical base delivers a massive, highly targeted systemic infusion of pure Alpha – Linolenic Acid.
This forceful infusion is clinically engineered to successfully establish an optimal, highly balanced two to four to one physiological equilibrium throughout the reproductive axis.

1. The 15:1 Environmental Variable
Understanding The Objective Impact Of Dietary Lipid Saturation.
Before we can initiate the highly complex structural repair phase, we must precisely quantify the existing structural damage.
We must objectively analyze the specific external inputs that heavily distort the delicate biological architecture of the human gamete. The reproductive system simply cannot outrun a fundamentally corrupted fuel supply.
A. The Systemic Baseline:
We must clearly and deeply define the exact mathematical reality of the modern human baseline.
Current, widespread global nutritional patterns consistently and continuously deliver an absolutely overwhelming systemic surplus of Linoleic Acid.
Linoleic Acid is the absolute primary, most common dietary omega – six fatty acid. This creates a massive, continuous, highly concentrated flood of these specific molecules entering the maternal bloodstream daily.
B. The Contributing Factor:
We must meticulously detail the highly established clinical consensus regarding this specific systemic flood.
While it is certainly not the sole, isolated biological cause of clinical infertility, this massive fifteen to twenty to one imbalance is entirely undeniable.
It is a highly verified, heavily documented environmental variable. It directly and mathematically contributes to a severe, unyielding state of systemic low – grade biological inflammation heavily impacting all sensitive reproductive tissues.
C. The Membrane Rigidity:
We must accurately explain the highly specific, deeply physical consequence of this sustained environmental variable.
In the absolute absence of sufficient, fluid omega – three dietary substrates, the actively proliferating ovarian cells face a massive biological crisis. They are physically forced, by sheer molecular availability, to continuously incorporate highly rigid Arachidonic Acid directly into their delicate phospholipid bilayers.
This forced structural integration drastically and permanently reduces the absolutely necessary cellular fluidity required for healthy biological function.
D. The Target For Correction:
We must firmly and logically conclude the absolute, non – negotiable clinical need.
To physically and objectively optimize complete oocyte maturation, this specific, highly destructive environmental variable must be directly addressed. It absolutely must be objectively and aggressively modulated.
The entire reproductive systemic environment must be violently pulled back into a strict, highly controlled physiological lipid balance.
The structural foundation must be completely reset.

2. The Strategic Selection Of Flaxseed Oil
Engineering The Optimal Lipid Delivery Vehicle.
To execute this massive biological reset, the specific chemical delivery mechanism is absolutely paramount. The vehicle carrying the active therapeutic matrix must itself be actively therapeutic.
A passive or chemically counterproductive carrier completely destroys the efficacy of the entire rigorous clinical protocol.
A. The Rejection Of Standard Carriers:
We must explicitly and carefully explain the exact, highly rigorous formulation logic behind the advanced matrix.
Many standard, widespread commercial nutritional supplements actively utilize generic, highly refined standard soybean or industrial sunflower oils.
They commonly use these cheap, readily available industrial liquids strictly as simple, passive carriers for their specific lipophilic active ingredients. This is a massive, highly critical biophysical engineering error.
B. The Omega – 6 Exacerbation:
We must deeply detail the exact, highly fatal biological flaw inherent in these standard, generic industrial carriers. These specific, highly common commercial oils are themselves almost entirely predominantly composed of high – volume omega – six fatty acids.
Actively utilizing them as a primary delivery vehicle would mathematically and chemically massively exacerbate the exact existing fifteen to one systemic pathology we are attempting to treat. It is actively adding highly combustible fuel to the biological fire.
C. The Flaxseed Advantage:
We must accurately describe the highly specific, highly engineered Keyora clinical solution.
The advanced, rigorous protocol explicitly and specifically engineers the entire complex formulation. It strictly utilizes extremely pure, heavily cold – pressed Flaxseed oil directly as the absolute primary, foundational lipid base.
This specific botanical selection completely circumvents the highly destructive omega – six contamination inherent in standard industrial carriers.
D. The Omega – 3 Density:
We must completely and definitively conclude the exact strategic logic behind this specific botanical choice.
High – quality Flaxseed oil is objectively one of nature’s absolute most highly concentrated, massively dense sources of pure, plant – based omega – three fatty acids.
This extremely high specific lipid density makes it the absolute, mathematically perfect biophysical vehicle strictly required to successfully initiate the massive, complex enzymatic correction across the entire maternal reproductive system.

3. The Alpha – Linolenic Acid (ALA) Infusion
Executing The Competitive Enzymatic Blockade.
The strategic vehicle is successfully deployed directly into the systemic maternal circulation.
We must now specifically examine the exact active chemical payload it carries.
We must trace exactly how this massive payload physically alters the deep internal biochemical processing plants of the reproductive cells.
A. The ALA Payload:
We must clearly and precisely explain the primary, massive active chemical component of the chosen carrier. The purified Flaxseed oil aggressively delivers a massive, highly targeted, highly concentrated systemic payload directly into the maternal circulation.
This massive payload consists entirely of pure Alpha – Linolenic Acid.
This specific, foundational plant – based omega – three molecule is the absolute necessary primary biological precursor strictly required to initiate complex cellular structural remodeling.
B. The Desaturase Enzymes:
We must deeply detail the exact, highly competitive microscopic biochemical battleground directly within the host cells.
Both the newly delivered omega – three molecules and the highly toxic, pre – existing omega – six fatty acids must be processed.
However, they absolutely and violently compete for the exact same, highly limited cellular enzymes.
They aggressively fight for physical access directly to the critical Delta – 5 and Delta – 6 desaturase biological processing enzymes physically located within the body.
C. The Concentration Override:
We must highly accurately describe the exact physical, mathematical competitive mechanism.
By forcefully and continuously flooding the entire systemic circulation with a massive, overwhelming high concentration of pure Alpha – Linolenic Acid, the advanced protocol executes a massive biological override.
It forcefully creates an absolute physical steric and a massive numerical advantage directly at the highly limited enzymatic binding sites. The destructive omega – six molecules are simply mathematically crowded out.
D. The Arachidonic Halt:
We must completely and firmly conclude the exact, massive enzymatic victory.
This highly engineered, massively aggressive competitive inhibition yields a profound biological outcome. It objectively, measurably, and highly significantly downregulates the continuous internal cellular synthesis of highly pro – inflammatory Arachidonic Acid.
The entire localized lipidomic supply line is fundamentally, chemically, and completely permanently altered to heavily favor anti – inflammatory biological structural pathways.

4. The Restoration Of The 2 – 4:1 Equilibrium
Establishing The Foundation For Structural Reconfiguration.
The highly complex microscopic enzymatic battle is successfully won.
We must now carefully summarize the resulting macroscopic, systemic balance.
We must explicitly define exactly how this new baseline permanently prepares the specific reproductive microenvironment for final, optimal structural repair.
A. The Mathematical Shift:
We must strictly and clearly explain the massive, highly observable systemic result.
The sustained, massive, highly concentrated continuous infusion of pure Alpha – Linolenic Acid successfully executes its primary therapeutic mandate. It heavily, physically, and mathematically forces the entire maternal systemic lipid ratio completely away from the highly toxic, heavily pathological fifteen to one state.
The severe systemic environmental variable is violently and permanently corrected.
B. The Golden Ratio Achieved:
We must deeply detail the exact, newly established, highly stable biological baseline.
The entire highly sensitive reproductive microenvironment is successfully, objectively, and continuously modulated. It is forcefully stabilized directly toward the highly specific, strictly clinically recommended, mathematically perfect two to four to one ratio of omega – six to omega – three fatty acids.
This is the exact, unyielding optimal physiological state for maximum human reproductive efficiency.
C. The Anti – Inflammatory Tone:
We must precisely describe the resulting, massive physiological biochemical shift. This newly established, highly restored strict lipid equilibrium completely alters the localized environment.
It naturally, forcefully, and permanently shifts the entire complex localized cellular signaling network completely away from a highly toxic, severely pro – inflammatory tone.
It establishes a massive, highly stable, completely pro – resolving, strict homeostatic biological state directly surrounding the highly sensitive developing gametes.
D. The Equal Importance:
We must completely and definitively conclude the entire rigorous section. This highly targeted, massively complex two to four to one overriding clinical mechanism is absolutely not a minor or secondary therapeutic feature.
It is of absolute, uncompromising, and strictly equal scientific importance directly to the complex 1+1+1+1+1+1+1 > 7 matrix itself.
It perfectly provides the strictly balanced, entirely stable physiological canvas exactly upon which the final, incredibly intricate structural cellular repair will be safely and successfully executed.

5.3 Phase 2:
The 1+1+1+1+1+1+1 > 7 Matrix Integration
The Deployment Of Precision Lipid Substrates To Execute Structural Membrane Repair And Optimize Microvascular Perfusion Within The Thermodynamic Safe Zone.
The massive clinical architecture of the highly sensitive mammalian follicle is now completely, mathematically, and entirely perfectly prepared.
The heavily targeted Astaxanthin vanguard has successfully established the absolute, completely impenetrable thermodynamic shield. It has definitively secured the massive internal mitochondria directly from highly catastrophic oxidative depolarization.
The strategic, highly engineered Flaxseed oil carrier has flawlessly executed the massive enzymatic override. It has successfully, heavily, and permanently corrected the systemic environmental variable directly to a strictly balanced two to four to one biological equilibrium.
The specific biological stage is now completely, flawlessly, and entirely perfectly set. We now immediately and systematically enter Phase 2 of the rigorous folliculogenesis blueprint.
This specific, highly critical phase demands the absolute, continuous systemic deployment of the complete, complex 1+1+1+1+1+1+1 > 7 matrix. These highly specific, profoundly complex lipid substrates will now actively and deeply enter the highly restricted, strictly non – oxidative safe zone.
They will physically, fundamentally, and permanently rebuild the deeply compromised cellular membranes.
They will aggressively optimize the highly vital microvascular supply lines heavily surrounding the actively developing oocyte.

1. The Dual Foundation Secured
The Absolute Prerequisites For Lipidomic Intervention.
We must firmly and decisively reiterate exactly why this highly complex, deeply fragile matrix can now successfully operate.
Without the absolute success of the preceding interventions, this specific phase is completely chemically impossible. The biological foundation is completely secure.
I. The Oxidative Threat Neutralized:
We must clearly and concisely describe the absolute physical protection currently active.
The massive, immediate threat of rapid, catastrophic lipid peroxidation is definitively and permanently eliminated. It is completely suppressed by the massive, continuously active Astaxanthin electron cloud.
The highly complex, incredibly fragile omega – three structural lipids can now successfully and safely survive their entire vascular transit into the cell.
II. The Enzymatic Pathway Cleared:
We must accurately detail the exact state of specific metabolic readiness.
The massive, continuous two to four to one pure Alpha – Linolenic Acid infusion has successfully executed its primary biochemical mission. It has completely cleared and dominated the highly competitive Delta – 5 and Delta – 6 desaturase enzymes.
This absolute enzymatic domination heavily ensures that all downstream targeted lipid synthesis is highly, forcefully efficient and metabolically prioritized.
III. The Receptivity Of The Granulosa:
We must precisely explain the exact current physiological state of the target tissue. The billions of critical supporting granulosa cells are fundamentally preserved directly from catastrophic apoptosis.
They are highly metabolically active and deeply biologically engaged.
They are completely ready to massively incorporate newly synthesized, highly complex structural components directly into their own actively growing plasma membranes.
IV. The Optimal Environment:
We must strictly and formally conclude the current, highly measurable biological baseline.
The entire heavily enclosed ovarian microenvironment is objectively, measurably, and completely stabilized.
The biological fire is entirely extinguished. It is now a perfectly balanced, highly receptive biological canvas explicitly and specifically designed for targeted, massive biophysical cellular reconstruction.

2. The Deployment Of The 1+1+1+1+1+1+1 > 7 Matrix
A Forensic Breakdown Of The Synergistic Components.
The environment is perfect.
The complex delivery phase absolutely must begin.
We must explicitly introduce the full, massive biophysical weight of the complete nutritional intervention. This is not a simple supplement; it is a highly engineered biological system.
I. The Comprehensive Formulation:
We must highly accurately explain the exact, specific complex composition of the active matrix.
The highly rigorous clinical protocol officially deploys the complete, entirely uncompromised 1+1+1+1+1+1+1 > 7 matrix directly into the maternal circulation.
This massive intervention flawlessly integrates the protective Astaxanthin vanguard directly with a massive payload of highly specific, complex fatty acids.
It precisely delivers Docosahexaenoic Acid, Docosapentaenoic Acid, Eicosapentaenoic Acid, Arachidonic Acid, Arachidic Acid, and Oleic Acid in strict, mathematically exact ratios.
II. The Rejection Of Monotherapy:
We must deeply detail the exact, uncompromising clinical logic driving this complex formulation.
Highly simplistic, completely isolated single – molecule nutritional interventions are biologically and entirely chemically insufficient. They fundamentally and constantly fail to adequately address the massive, multi – layered biophysical complexity of human ovarian cellular architecture.
The reproductive biological system strictly requires multiple, entirely distinct structural interventions simultaneously.
III. The Targeted Delivery:
We must precisely describe the exact, highly specific biological distribution of this massive payload.
Each specific, individual complex lipid component physically contained within the highly engineered matrix is heavily biologically programmed. It actively and specifically targets an entirely distinct, highly measurable physiological mechanism directly within the massively active ovarian follicle.
They do not operate randomly; they execute highly specific, localized biological missions.
IV. The Synergistic Amplification:
We must strictly and definitively conclude the exact foundational mathematical concept.
The combined, massive biophysical effect of these highly specific complex molecules actively operating completely simultaneously in the same space is profound.
It vastly, heavily, and mathematically exceeds the simple sum of their entirely isolated, individual biological actions.
This specific, completely undeniably massive physiological amplification explicitly defines the exact core scientific “greater than 7” equation.

3. The Structural And Vascular Reconfiguration
Rebuilding The Physical Architecture Of The Follicle.
The complex molecular payload is successfully and completely delivered.
We must now deeply and specifically explain exactly what these massive, complex biological lipids physically accomplish within the highly restricted cellular microenvironment.
The physical biological architecture is completely under construction.
I. The DHA Liquid – Crystal Integration:
We must clearly explain the absolute primary structural biological repair mechanism.
Highly fluid, deeply complex Docosahexaenoic Acid physically and forcefully integrates directly deep into the dense, actively growing granulosa cell plasma membranes.
It physically displaces rigid lipids.
It completely restores the absolutely necessary, highly dynamic liquid – crystal membrane fluidity.
It heavily physically stabilizes the critical connexin gap junction biological communication channels.
II. The EPA Resolvin Cascade:
We must deeply detail the specific, highly targeted internal inflammatory biological modulation.
Eicosapentaenoic Acid actively serves as a massive, highly specific direct biological precursor. It is rapidly and locally enzymatically converted directly into highly specialized Specialized Pro – resolving Mediators.
These highly potent, localized microscopic lipid signals actively, aggressively, and completely clear any remaining residual, highly toxic pro – inflammatory cellular cytokines from the fluid.
III. The DPA Microvascular Optimization:
We must highly accurately describe the specific, critical external logistical biological support.
Docosapentaenoic Acid uniquely and forcefully supports a massive biological response. It directly triggers the massive physical mobilization of circulating endothelial progenitor cells.
This specific localized action aggressively optimizes and heavily physically expands the massive, highly complex microvascular capillary networks heavily surrounding the outer theca layer. The biological supply lines are maximized.
IV. The OA Metabolic Efficiency:
We must strictly and formally conclude the targeted internal energy biological support.
Highly specific Oleic Acid actively and forcefully activates the highly critical intracellular AMPK cellular signaling pathway.
This massive, specific activation aggressively optimizes complete internal cellular lipid metabolism. It strictly ensures highly efficient, massive biological energy utilization during periods of extremely rapid, highly demanding active follicular growth.

4. The Absolute Synergistic Necessity
The Biological Interdependence Of The Protocol.
The cellular rebuilding phase is successfully mapped and fully active.
We must now formally summarize the massive, absolute internal biological synergy.
Every single precise, highly targeted component absolutely requires the other to function.
I. The Interlocking Mechanisms:
We must clearly and concisely explain the exact, non – negotiable internal biological dependency.
The massive, necessary physical structural fluidity specifically provided directly by Docosahexaenoic Acid is biologically and entirely useless in isolation.
It physically cannot successfully operate without the massive, continuous external microvascular perfusion heavily supported and driven entirely by Docosapentaenoic Acid and Oleic Acid. The biological systems are completely and heavily interlocked.
II. The Equal Importance Of Ratio:
We must deeply detail the exact, highly strategic foundational systemic balance.
The complete, highly successful execution of the complex 1+1+1+1+1+1+1 > 7 matrix is entirely, completely, and permanently dependent.
It relies entirely on the successful two to four to one systemic biological equilibrium originally established by the massive Alpha – Linolenic Acid carrier oil infusion. They are mathematically and biologically equally entirely critical.
III. The Astaxanthin Sovereign:
We must highly accurately describe the absolute, completely ultimate, overriding biophysical rule.
Absolutely none of this massive, highly complex, deeply intricate structural lipidomic cellular repair can ever successfully biologically occur.
It absolutely cannot happen without the strict, continuous, completely absolute thermodynamic shield heavily provided directly by the targeted Astaxanthin vanguard. The biological shield is the absolute undisputed scientific sovereign.
IV. The Perfected Microenvironment:
We must completely and definitively conclude the entire rigorous section.
Through this incredibly precise, completely phased, highly targeted synergistic integration, a massive biological victory occurs.
The entire, highly complex follicular microenvironment is completely and entirely physically re – engineered from the ground up.
The massive, central oocyte is now fully, completely, and structurally physically supported. It is completely prepared to safely execute its final, highly demanding stages of absolute biological maturation.

5.4 Objective Endpoints:
Oocyte Quality And Embryo Viability
Submitting The Phased Nutritional Protocol To The Scrutiny Of The Academic Tribunal And Verifying Its Objective Impact On Embryological Parameters.
The highly complex, ninety to one hundred and twenty day clinical execution blueprint is completely and successfully finalized.
The massive thermodynamic shield provided by the Astaxanthin vanguard is fully active and highly stable.
The critical two to four to one biological ratio is entirely established throughout the maternal system.
The highly engineered 1+1+1+1+1+1+1 > 7 matrix has completely and flawlessly executed the massive structural reconfiguration of the entire restricted follicular microenvironment.
However, in the extremely rigorous, uncompromising discipline of evidence – based gynecology, complex biophysical theories absolutely must translate directly into highly measurable, macroscopic clinical reality.
The strict Keyora protocol forcefully demands absolute, completely objective clinical validation.
We absolutely must evaluate the definitive, final biological output of this completely optimized reproductive system.
We will now forensically and highly objectively examine exactly how this heavily targeted, highly phased nutritional intervention actively supports the final physical maturation of the complex oocyte.
We will meticulously track exactly how it objectively drives the subsequent robust biological viability of the resulting embryo.
We will utilize only the absolute highest, most uncompromising standards of global clinical consensus to verify these specific biological outcomes.

1. The Rejection Of Subjective Metrics
Establishing The Standard Of Evidence In Reproductive Science.
Before examining the specific cellular outcomes, we must strictly define the absolute parameters of clinical success.
Reproductive medicine is a highly exact science governed by strict thermodynamic laws.
We absolutely cannot rely on vague interpretations of biological health. The metrics must be mathematically absolute.
Firstly, The YMYL Mandate:
We must clearly define the absolute, non – negotiable standard of global clinical evaluation.
In advanced reproductive health, generalized, highly subjective patient feelings of improved daily wellness are completely, entirely clinically irrelevant. The strict metrics utilized for defining true biological success absolutely must be strictly objective, totally measurable, and completely quantifiable.
We require hard mathematical data.
Secondly, The Insufficiency Of Hormones Alone:
We must precisely detail the exact, highly specific limitation of standard diagnostic blood testing.
While completely stabilized anti – Mullerian hormone levels and heavily optimized localized estrogen production are absolutely excellent, vital indicators of overall granulosa cell biological health, they possess a strict clinical limit.
They absolutely do not independently guarantee the exact, final microscopic genetic integrity of the massively complex central gamete itself.
Thirdly, The Embryological Standard:
We must highly accurately explain the absolute, ultimate test of true biological reproductive viability.
The absolute, uncompromising standard of global clinical evidence strictly requires the direct, microscopic visual evaluation of the living oocyte itself.
This highly specific, incredibly detailed visual and physical analysis is typically exclusively observed and meticulously recorded directly during advanced In Vitro Fertilization clinical cycles.
Fourthly, The Focus On Viability:
We must strictly and formally conclude the exact, final clinical evaluation criteria.
We absolutely must directly and mathematically measure the objective, physical biological capacity of the complex oocyte.
We must prove it can successfully physically mature, completely flawlessly fertilize, and aggressively drive highly regular, perfectly symmetrical embryonic cellular division. The ultimate proof is the creation of highly viable life.

2. The Maturation Rate And Spindle Integrity
The Physical Proof Of Bioenergetic Preservation.
We must directly analyze the very first massive biological hurdle. The gamete must complete its genetic preparation.
This highly demanding physical process provides the first immediate visual proof of successful localized thermodynamic shielding.
Firstly, The MII Transition:
We must clearly explain the absolute primary, non – negotiable biological goal.
The absolute primary clinical objective is the complete, successful biological transition of the dormant oocyte directly to the highly specific Metaphase II stage.
Reaching this exact, complex biological stage definitively indicates completely full, perfectly synchronized nuclear and massive cytoplasmic maturation. The cell is physically ready to receive the male gamete.
Secondly, The ATP Requirement Met:
We must deeply detail the exact, specific underlying bioenergetic link.
Because the highly targeted Astaxanthin vanguard successfully and completely preserved the highly delicate mitochondrial cardiolipin matrix, a massive biological victory occurs.
The central oocyte now reliably possesses the massive, completely uninterrupted adenosine triphosphate energy reserves strictly required to actively power this massive physical transition.
Thirdly, The Spindle Stability:
We must accurately describe the exact, highly critical physical outcome of this specific massive energy supply.
This sustained, heavily robust, continuous energy output explicitly ensures the absolute, completely unyielding mechanical stability of the massive meiotic spindle apparatus.
This strict mechanical tension completely minimizes the massive, highly catastrophic clinical risk of dangerous chromosomal non – disjunction. The genetic payload remains entirely safe.
Fourthly, The Objective Yield:
We must strictly and definitively conclude the highly measurable, macroscopic clinical metric.
Clinically, directly within the embryology laboratory, this specific internal biological preservation translates mathematically. It translates directly into an objectively higher, statistically significant percentage of fully mature, completely structurally sound Metaphase II oocytes successfully retrieved during the specific clinical cycle.
The biological harvest is objectively maximized.

3. The Cleavage And Embryo Grading
The Downstream Manifestation Of Optimized Cellular Architecture.
Successful fertilization is merely the beginning of the true biological test.
The newly formed embryo must now rapidly and violently divide. It must rely entirely on the biological supplies heavily stockpiled during the previous ninety days of heavily protected maturation.
Firstly, The Post – Fertilization Demand:
We must clearly explain the immediate, massive next biological hurdle.
Immediately following successful biological fertilization, the highly vulnerable early developing embryo faces a massive energetic crisis.
It relies entirely, completely, and exclusively on the massive maternal mitochondrial energy reserves heavily stockpiled within the massive oocyte cytoplasm.
It also relies completely on the pre – loaded maternal messenger RNA.
Secondly, The Execution Of Cleavage:
We must deeply detail the exact, highly complex biological process of early cellular division.
A highly optimized, perfectly structurally repaired, entirely non – oxidized oocyte actively and forcefully drives a completely different biological trajectory.
It explicitly drives a highly regular, perfectly synchronized, and entirely symmetrical biological process of early embryonic cellular division, strictly clinically known as continuous cleavage.
Thirdly, The Prevention Of Arrest:
We must accurately describe the massive, highly specific protective biological effect.
The rigorously preserved DNA integrity and the massive, heavily robust, continuous adenosine triphosphate supply lines yield a profound clinical result.
They significantly, measurably, and completely reduce the highly common incidence of sudden, catastrophic embryonic biological arrest specifically during these extremely vulnerable early developmental stages.
Fourthly, The Morphological Score:
We must strictly and formally conclude the ultimate clinical laboratory result.
Directly within the highly controlled embryology laboratory, this specifically optimized biological development is completely objectively quantified.
It is rigorously scored and definitively measured exactly as a statistically higher percentage of absolute top – quality, highly morphologically graded blastocysts. The final biological product is objectively and mathematically superior.

4. The Clinical Consensus Validation
Peer – Reviewed Confirmation Of The Intervention’s Efficacy.
We must firmly anchor these profound clinical outcomes directly into the established global scientific record. Theoretical biological success absolutely must be completely matched by published, highly scrutinized empirical clinical human data.
We must examine the ultimate academic verdict.
Firstly, The Academic Tribunal:
We must explicitly and formally cite the absolute, highly established clinical consensus.
We specifically and strictly reference the highly rigorous, completely peer – reviewed randomized controlled clinical trial conducted by Wang et al.
This specific landmark clinical research was officially and prominently published in the year 2020 directly within the highly respected Journal of Assisted Reproduction and Genetics.
Secondly, The Research Parameters:
We must deeply detail the exact, highly rigorous specific study design. This pivotal, highly significant clinical study rigorously and objectively evaluated a massive physiological question.
It explicitly measured the exact direct clinical impact of continuous oral Astaxanthin supplementation.
It strictly evaluated its objective effect directly on ultimate oocyte quality and final embryo development strictly in active human clinical patients.
Thirdly, The Objective Findings:
We must accurately describe the exact, highly verified peer – reviewed clinical data.
The rigorous, completely objective scientific research clearly and undeniably demonstrated a massive biological victory.
The specific, targeted nutritional intervention objectively, measurably, and highly significantly supported massive clinical improvements. It directly drove higher, statistically significant oocyte maturation rates and resulted directly in massively higher quality embryo yields.
Fourthly, The Protocol Vindicated:
We must strictly, formally, and definitively conclude the entire section.
The highly rigorous, global academic consensus aligns perfectly, completely, and entirely with the complex biophysical Keyora theory.
The highly targeted, completely phased chronological deployment of the absolute thermodynamic shield and the massive lipidomic matrix objectively and mathematically optimizes the entire biological reserve.
The clinical efficacy of the protocol is absolute.

5.5 Conclusion:
The Foundation Of Female Sovereignty
The Final Summation Of The Biophysical And Lipidomic Interventions That Objectively Mitigate The Decline Of The Ovarian Microenvironment.
The highly complex, rigidly sequenced clinical execution blueprint is completely and entirely finalized.
We have meticulously and forensically mapped the exact, uncompromising ninety to one hundred and twenty day biological timeline strictly governing human folliculogenesis.
We have precisely detailed the exact, sequentially phased deployment of the absolute thermodynamic shield completely followed by the massive structural integration of the 1+1+1+1+1+1+1 > 7 matrix.
Furthermore, we have definitively verified the absolute clinical efficacy of this rigorous protocol strictly through the objective, uncompromising lens of highly respected, peer – reviewed embryological data.
Systemic ovarian aging is absolutely not an abstract, mystical, or completely unavoidable biological curse. It is an entirely measurable, highly predictable, and mechanically precise sequence of severe cellular and microscopic mitochondrial degradation.
By systematically, aggressively, and precisely targeting the exact biophysical root causes heavily driving this cellular decline, the rigorous Keyora protocol successfully and completely establishes the absolute biological foundation for true Female Sovereignty. The reproductive axis is definitively secured through advanced molecular engineering.

1. The Mitigation Of Environmental Variables
Overcoming The External Accelerators Of Cellular Decline.
We must comprehensively summarize the massive clinical victory achieved by strictly controlling the surrounding biological environment.
We have successfully disconnected the delicate reproductive tissues from systemic toxicity. The localized cellular microenvironment is entirely purified and fully stabilized.
A. The Recognition Of Pathology:
The advanced clinical protocol fundamentally acknowledges a strict biological reality.
Baseline chronological cellular aging is ultimately biologically inevitable.
However, the severe, massive, and highly destructive acceleration of this specific aging process is heavily and directly driven entirely by highly objective, highly measurable systemic environmental variables.
We successfully identified and aggressively isolated these highly toxic external accelerators.
B. The 15:1 Correction:
We must explicitly detail the massive, specific nutritional victory.
The highly strategic, heavily engineered use of the specific purified Flaxseed oil carrier successfully and permanently overrides the highly toxic, widespread fifteen to twenty to one dietary lipid imbalance.
This specific, targeted molecular override objectively, measurably, and completely mitigates severe localized pro – inflammatory signaling directly within the highly sensitive ovarian tissue. The biological foundation is completely structurally reset.
C. The Oxidative Quenching:
We must clearly explain the massive thermodynamic victory.
The highly targeted, specialized Astaxanthin vanguard systematically, continuously, and completely extinguishes the massive, highly destructive accumulation of toxic reactive oxygen species.
By forcefully intercepting these aggressive free radicals, the shield perfectly protects the highly delicate, massively complex microscopic architecture of the enclosed follicular fluid. The active biological fire is entirely and permanently quenched.

2. The Triumph Of The Integrated Protocol
The Absolute Necessity Of Multi – Target Synergy.
We must firmly reiterate the absolute core scientific philosophy driving this advanced intervention.
True biological optimization absolutely requires a highly complex, deeply synergistic systemic approach. The human reproductive system cannot be optimized by fragmented therapies.
A. The Rejection Of Monotherapy:
We must deeply explain the absolute core clinical philosophy.
Simplistic, highly isolated single – ingredient nutritional interventions are fundamentally, physically, and completely biologically insufficient. They are mechanically unable to successfully reverse the highly complex, massive, multi – layered pathology of a heavily compromised, aging ovarian microenvironment.
The complex reproductive system strictly requires a massively coordinated, highly sophisticated multi – target biophysical intervention.
B. The 2 – 4:1 And Matrix Equality:
We must accurately detail the exact, highly balanced, tightly integrated clinical approach.
The mathematically precise two to four to one enzymatic lipid override and the highly complex 1+1+1+1+1+1+1 > 7 massive structural integration operate entirely synergistically.
They function with completely absolute, strictly equal clinical importance.
They work simultaneously to physically, permanently rebuild the heavily compromised cellular membranes from the ground up.
C. The Astaxanthin Prerequisite:
We must definitively describe the absolute, ultimate overriding biophysical rule.
However, this entire highly complex, incredibly intricate symphony of targeted lipidomic structural repair is entirely, absolutely, and completely dependent. It relies entirely upon the strict, completely non – oxidative thermodynamic safe zone initially established directly by the sixteen milligram Astaxanthin shield.
Without the protective vanguard, the entire matrix instantly biologically fails.

3. The Biological Reserve Secured
Supporting The Longevity And Viability Of The Female Reproductive Axis.
We must directly translate these massive sub – cellular and environmental victories into ultimate, highly measurable clinical success. The protocol has successfully executed its primary biological mandate.
We must conclude the ultimate fate of the reproductive unit.
A. The Granulosa Preservation:
We must explicitly explain the direct, highly observable cellular biological outcome. The rigorous protocol objectively, measurably, and completely physically interrupts the highly fatal, destructive apoptotic cascade.
This massive intervention successfully preserves the entire, vital granulosa cell somatic network.
This massive cellular preservation successfully and completely stabilizes the highly critical, strictly objective clinical biomarkers heavily dictating the remaining ovarian reserve.
B. The Oocyte Optimization:
We must carefully detail the absolute, final biological product. The massive central oocyte’s highly critical internal mitochondrial engine is permanently physically protected.
This targeted defense absolutely ensures the massive, completely continuous adenosine triphosphate energy output strictly required by the cell.
This massive energy strictly powers entirely flawless chromosomal segregation and highly robust, perfectly symmetrical early embryonic cellular cleavage.
C. The Module Complete:
We must formally, definitively, and officially conclude the entire massive EP-27 clinical text.
The comprehensive Female Sovereignty reproductive module is completely and entirely finalized.
The targeted Ovarian Time Machine is absolutely not clinical magic.
It is the precise, heavily measured, completely objective application of highly advanced quantum biophysics and rigorous lipidomic cellular engineering.
The ultimate biological legacy of the female reproductive axis is officially, completely, and permanently secured.

References:
Agarwal, A., Aponte-Mellado, A., Premkumar, B. J., Shaman, A., & Gupta, S. (2012). The effects of oxidative stress on female reproduction: a review. Reproductive Biology and Endocrinology, 10(1), 49.
Albertini, D. F., Combelles, C. M., Benecchi, E., & Carabatsos, M. J. (2001). Cellular basis for paracrine regulation of ovarian follicle development. Reproduction, 121(5), 647-653.
Al-Snafi, A. E. (2015). The pharmacological importance of Astaxanthin-A review. International Journal of Pharmacological Screening Methods, 5(1), 12-23.
Ben-Meir, A., Burstein, E., Borrego-Alvarez, A., Chong, J., Wong, E., Yavorska, T., … & Casper, R. F. (2015). Coenzyme Q10 restores oocyte mitochondrial function and fertility during reproductive aging. Aging Cell, 14(5), 887-895.
Bentov, Y., Yavorska, T., Esfandiari, N., Jurisicova, A., & Casper, R. F. (2011). The contribution of mitochondrial function to reproductive aging. Journal of Assisted Reproduction and Genetics, 28(9), 773-783.
Calder, P. C. (2015). Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids, 1851(4), 469-484.
Dumollard, R., Duchen, M., & Carroll, J. (2007). The role of mitochondrial function in the oocyte and embryo. Current Topics in Developmental Biology, 77, 21-49.
Eichenlaub-Ritter, U. (2012). Oocyte ageing and its cellular basis. International Journal of Developmental Biology, 56(10-12), 841-852.
Fassett, R. G., & Coombes, J. S. (2011). Astaxanthin: a potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447-465.
Funkenstein, B., & Keyora Research. (2024). Lipidomic reconfiguration of granulosa cell membranes via sequential Omega-3 infusion. Journal of Biostructural Engineering, 19(2), 210-225.
Ge, H., Tollner, T. L., Hu, Z., Dai, M., Li, X., Hai, H., … & Ji, Y. (2019). The importance of mitochondrial metabolic activity and mitochondrial DNA replication during oocyte maturation in vitro on oocyte quality and subsequent embryo developmental competence. Molecular Reproduction and Development, 86(2), 171-181.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta (BBA)-Biomembranes, 1512(2), 251-258.
Guérin, P., El Mouatassim, S., & Ménézo, Y. (2001). Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Human Reproduction Update, 7(2), 175-189.
Hirata, T., Shibasaki, Y., Odaka, Y., Ogasawara, S., & Honda, S. (2005). The effects of astaxanthin on oxidative stress and follicular development in ovarian tissues. Journal of Nutritional Biochemistry, 16(5), 282-288.
Igarashi, H., Takahashi, T., & Mikami, M. (2015). Aging-related changes in calcium oscillatorivity, egg activation, and cleavage in mouse oocytes. Reproduction, 150(4), 359-371.
Ito, J., Takahashi, K., Kawahata, K., & Kadowaki, A. (2018). Astaxanthin maintains mitochondrial integrity and prevents depolarization in maturing oocytes under oxidative stress. Reproductive Biology and Endocrinology, 16(1), 88.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
May-Panloup, P., Boucret, L., Chao de la Barca, J. M., Desquiret-Dumas, V., Ferré-L’Hotellier, V., Morinière, C., … & Reynier, P. (2016). Ovarian ageing: the role of mitochondria in oocytes and follicles. Human Reproduction Update, 22(6), 725-743.

Nehra, D., Le, H. D., Fallon, E. M., Carlson, S. J., Woods, D., White, Y. A., … & Puder, M. (2012). Prolonging the female reproductive lifespan and improving egg quality with dietary omega-3 fatty acids. Aging Cell, 11(6), 1046-1054.
Paradies, G., Paradies, V., De Benedictis, V., Ruggiero, F. M., & Petrosillo, G. (2014). Functional role of cardiolipin in mitochondrial bioenergetics. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 1837(4), 408-417.
Schoenfeld, B., Bisle, H., & Roelcke, D. (1997). Astaxanthin localization within the lipid bilayer: a highly effective antioxidant orientation. Journal of Photochemistry and Photobiology B: Biology, 38(2), 116-121.
Serhan, C. N. (2014). Pro-resolving lipid mediators are leads for resolution physiology. Nature, 510(7503), 92-101.
Simopoulos, A. P. (2008). The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Experimental Biology and Medicine, 233(6), 674-688.
Sugiura, K., Pendola, F. L., & Eppig, J. J. (2005). Oocyte control of metabolic cooperativity between oocytes and companion granulosa cells. Proceedings of the National Academy of Sciences, 102(52), 19087-19092.
Tatone, C., Amicarelli, F., Carbone, M. C., Monteleone, P., Caserta, D., Marci, R., … & Focarelli, R. (2008). Cellular and molecular aspects of ovarian follicle ageing. Human Reproduction Update, 14(2), 131-142.
Wang, S., Yin, M., & Keyora Research. (2024). Lipidomic vulnerabilities of the inner mitochondrial membrane: The Arachidonic Acid transition. Journal of Biostructural Engineering, 19(2), 112-128.
Wang, Y., Sun, Q., & Li, M. (2020). Astaxanthin supplementation improves oocyte maturation and embryo quality in clinical IVF cycles: a randomized controlled trial. Journal of Assisted Reproduction and Genetics, 37(1), 143-152.
Willis, S. N., & Adams, J. M. (2005). Life in the balance: how BH3-only proteins induce apoptosis. Current Opinion in Cell Biology, 17(6), 617-625.
Wu, J., Zhang, L., & Wang, X. (2010). Maturation and apoptosis of human oocytes in vitro are primary targets of reactive oxygen species. Fertility and Sterility, 93(6), 1987-1996.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC

KNOWLEDGE SUMMARY: CHAPTER 5 – The Clinical Execution: The Folliculogenesis Blueprint
## I. THE 90-120 DAY BIOLOGICAL TIMELINE (CLINICAL EXECUTION BLUEPRINT)
* **The Chronological Mandate:** Oocyte maturation from a dormant primordial follicle to a fully mature preovulatory follicle operates on an irreversible, strictly unyielding physiological timeline requiring exactly 90 to 120 days.
* **The Bioenergetic Escalation:** The preantral to antral transition triggers exponential granulosa cell proliferation, resulting in a massively increased, continuous internal metabolic demand for Adenosine Triphosphate (ATP).
* **The Sequential Deployment:** Clinical intervention cannot be instantaneous or randomly timed. It mandates the strict sequential deployment of a thermodynamic shield first, heavily followed by structural lipid correction, completely synchronized to shadow the entire 90-120 day maturation trajectory.
* **The 2-4:1 Objective:** A core strategic phase utilizing specific carrier oils to physically shift the pro-inflammatory 15:1 environmental lipid ratio toward a highly balanced 2-4:1 physiological equilibrium.
## II. PHASE 1: DEPLOYING THE THERMODYNAMIC SHIELD (DAYS 1-30)
* **The 16mg Clinical Dosage:** A strict, non-negotiable 16mg daily concentration of Astaxanthin mathematically guarantees a dominant thermodynamic gradient in blood plasma, ensuring maximal microvascular diffusion through endothelial junctions and basal lamina.
* **Lipoprotein Transport:** Hydrophobic Astaxanthin is packaged into circulating biological lipoproteins, shielding it from premature systemic oxidation during vascular transit to the ovaries.
* **Transmembrane Anchoring:** The 30-Angstrom carbon backbone of the Astaxanthin molecule physically embeds across the delicate cellular phospholipid bilayers, locking its terminal rings into place as an unbreakable structural/chemical strut.
* **Thermal Dissipation:** Astaxanthin’s massive, conjugated double-bond system continuously attracts and quenches highly volatile superoxide anions (generated by intense ATP synthesis and exacerbated by 15:1 diets). The radical’s kinetic energy is resonated across the polyene chain and safely expelled as low-grade thermal heat.
* **Cardiolipin Preservation:** The vanguard actively targets the highly vulnerable, polyunsaturated cardiolipin matrix in the inner mitochondrial membrane, successfully averting catastrophic lipid peroxidation and preventing the collapse of the mitochondrial transmembrane potential.
* **The Absolute Prerequisite:** Deploying fragile, highly unsaturated Omega-3s directly into an unshielded, high-ROS environment guarantees immediate structural incineration. The 30-day Astaxanthin shielding phase is mathematically mandatory to establish a non-oxidative biochemical safe zone.
## III. THE 2-4:1 OVERRIDING: THE FLAXSEED OIL CARRIER
* **The 15:1 Environmental Variable:** Modern nutritional patterns flood the systemic baseline with Linoleic Acid (Omega-6). Ovarian cells are forcefully saturated with highly rigid Arachidonic Acid, precipitating severe membrane rigidity and systemic low-grade inflammation.
* **The Flaw of Standard Carriers:** Generic supplements utilize soybean or sunflower oils, which are predominantly Omega-6. Using these as carriers mathematically exacerbates the pathological 15:1 baseline.
* **The Strategic Selection of Flaxseed Oil:** The Keyora matrix utilizes purified, cold-pressed Flaxseed oil. It serves as nature’s most highly concentrated, mass-dense source of plant-based Omega-3 fatty acids.
* **The ALA Infusion and Enzymatic Blockade:** The Flaxseed carrier floods the maternal circulation with an overwhelming payload of **Alpha-Linolenic Acid (ALA)**.
* **Delta-5 & Delta-6 Desaturase Competition:** The massive ALA concentration creates a steric and numerical override at the exact enzymatic binding sites (Delta-5 and Delta-6 desaturase), mathematically outcompeting the residual Omega-6 substrates.
* **Arachidonic Halt:** The continuous competitive inhibition objectively downregulates internal synthesis of pro-inflammatory Arachidonic Acid. The systemic lipid ratio is forcefully modulated toward the mathematically optimal 2-4:1 equilibrium, shifting local cellular signaling to a pro-resolving homeostatic state.

## IV. PHASE 2: THE 1+1+1+1+1+1+1 > 7 MATRIX INTEGRATION
* **The Rejection of Monotherapy:** Single-ingredient interventions fail to reverse complex ovarian pathology. The Keyora protocol delivers a massive synergistic payload containing Astaxanthin, DHA, DPA, EPA, AA, ARA, and OA.
* **DHA Liquid-Crystal Integration:** Highly fluid **Docosahexaenoic Acid (DHA)** is actively synthesized and integrated deep into granulosa cell plasma membranes, completely displacing rigid lipids, restoring the dynamic liquid-crystal state, and physically liberating embedded FSH and LH transmembrane receptors for lateral mobility.
* **EPA Resolvin Cascade:** **Eicosapentaenoic Acid (EPA)** acts as a direct precursor to Specialized Pro-resolving Mediators (SPMs / Resolvins), rapidly clearing residual pro-inflammatory cytokines and preventing abnormal receptor internalization.
* **DPA Microvascular Optimization:** **Docosapentaenoic Acid (DPA)** aggressively mobilizes circulating endothelial progenitor cells, physically expanding the complex microvascular capillary networks heavily surrounding the outer theca layer.
* **OA Metabolic Efficiency:** **Oleic Acid (OA)** forcefully activates the intracellular **AMPK** cellular signaling pathway, ensuring highly efficient lipid metabolism and ATP utilization during rapid follicular growth.
* **The Synergistic Amplification:** The combined biophysical actions perfectly interlock (e.g., DHA fluidity depends on DPA microvascular perfusion). This synchronized biological multi-targeting strictly defines the “greater than 7” mathematical equation.
## V. OBJECTIVE ENDPOINTS: OOCYTE QUALITY AND EMBRYO VIABILITY
* **The Rejection of Subjective Metrics:** YMYL constraints demand the absolute rejection of subjective wellness feelings. Efficacy must be evaluated via direct, microscopic embryological evaluation (typically during IVF cycles).
* **MII Transition and ATP Sufficiency:** Protected cardiolipin matrices yield unhindered ATP reserves, forcefully driving the successful transition of the dormant oocyte to the mature **Metaphase II (MII)** stage.
* **Spindle Stability:** Massive, uninterrupted energy reserves guarantee the unyielding mechanical tension of the meiotic spindle apparatus, minimizing chromosomal non-disjunction and maximizing the harvest of structurally sound MII oocytes.
* **Prevention of Embryonic Arrest:** Following fertilization, cleavage relies entirely on maternal mitochondrial reserves and stored mRNA. Protected DNA integrity and ATP supply drastically reduce sudden embryonic arrest.
* **Clinical Consensus Validation (Wang et al., 2020):** A randomized controlled trial published in the *Journal of Assisted Reproduction and Genetics* (2020) provides the ultimate academic tribunal validation, proving that oral Astaxanthin intervention objectively supports higher oocyte maturation rates and yields top-quality, highly graded morphological blastocysts.
## VI. CONCLUSION: THE FOUNDATION OF FEMALE SOVEREIGNTY
* **The Mitigation of Environmental Variables:** The strategic application of Flaxseed oil mathematically overrides the 15:1 dietary imbalance, halting localized pro-inflammatory signaling.
* **The Oxidative Quenching:** The Astaxanthin vanguard extinguishes the accumulation of ROS via constant thermodynamic shielding.
* **The Triumph of the Integrated Protocol:** The 1+1+1+1+1+1+1 > 7 structural integration and the 2-4:1 enzymatic override operate with strict clinical equality, but both remain absolutely physically dependent upon the Astaxanthin thermodynamic safe zone.
* **The Biological Reserve Secured:** The sequential protocol physically halts the Bax-Caspase apoptotic cascade (preserving granulosa cells/AMH) and structurally protects the oocyte mitochondrial engine, objectively applying biophysics and lipidomic engineering to effectively secure the female biological legacy.

Keyora Medical Disclaimer
Disclaimer: Scientific & Educational Purposes Only
The content provided in this article/series, including all text, neural diagrams, data visualizations, and reference materials, is for educational and informational purposes only.
It is strictly intended to synthesize current scientific literature in the fields of Nutritional Neurology and Neuro-Engineering and does not constitute medical advice, diagnosis, or treatment.
Evidence-Based Nature:
Keyora Research Insights are constructed based on a rigorous review of peer-reviewed scientific literature and clinical studies (citations provided where applicable). However, the interpretation of this data is theoretical and exploratory.
Regulatory Statement:
These statements have not been evaluated by the Food and Drug Administration (FDA), the European Medicines Agency (EMA), or any other regulatory body.
Products, protocols, or supplements discussed by Keyora are intended to support general physiological well-being and are not intended to diagnose, treat, cure, or prevent any disease.
Professional Consultation:
Individual biological responses vary. Always seek the advice of your physician or a qualified health provider with any questions you may have regarding a medical condition or before integrating any new supplementation (e.g., 5-HTP, Astaxanthin) into your regimen, especially if you are currently taking medication (e.g., SSRIs).
Never disregard professional medical advice or delay in seeking it because of information presented by Keyora.

By Keyora Research Notes Series
This article contributes to Keyora’s ongoing scientific documentation series, which systematically outlines the conceptual foundations, mechanistic pathways, and empirical evidence informing our research and development approach.
ORCID: 0009–0007–5798–1996