Keyora Astaxanthin EP-24: The Astaxanthin Imperative: Quenching Mitochondrial ROS And Reversing DNA Cleavage
By Keyora Research Notes Series
This article contributes to Keyora’s ongoing scientific documentation series, which systematically outlines the conceptual foundations, mechanistic pathways, and empirical evidence informing our research and development approach.
ORCID: 0009–0007–5798–1996

The Hydrodynamic Sacrifice:
The Stripped Architecture Of The Spermatozoon
Deconstructing The Evolutionary Trade – Off Between Maximum Flagellar Propulsion And The Total Loss Of Endogenous Antioxidant Defense
To fully comprehend the structural reality of the spermatozoon, one must first discard the traditional cellular model and analyze this entity through the strict parameters of biophysics and fluid dynamics.
Unlike a standard somatic cell, which prioritizes metabolic stability and sustained internal homeostasis, the male gamete undergoes a radical, irreversible architectural modification during the late stages of spermiogenesis. It is engineered with a singular, uncompromising biological imperative: successful transit through a hostile, highly viscous external environment to deliver an intact genetic payload.
To achieve this, the cell transforms into a highly specialized biological missile. Every internal structure that does not directly contribute to either flagellar propulsion or genetic storage is systematically disassembled and eliminated. This extreme morphological adaptation yields unparalleled hydrodynamic efficiency, minimizing drag and maximizing forward trajectory velocity.
However, this physiological optimization demands a profound evolutionary trade – off, forcing the gamete to permanently sacrifice its own foundational structural architecture and, consequently, its inherent capacity to weather thermodynamic and biochemical stress.

1. The Cytoplasmic Extrusion
The Physical Discarding Of The Intracellular Defense Matrix
The terminal phase of spermatid maturation is characterized by a violent physical reduction, a cellular shedding process governed by the necessity of minimizing spatial volume and excess weight.
This mechanism, while hydrodynamically advantageous, permanently alters the operational parameters of the surviving cell.
I. The Mass Reduction:
As the maturing spermatozoon prepares for its kinetic lifespan, it executes an organized physical extrusion of nearly all its cytoplasmic volume.
In the physics of micro – fluid propulsion, any unnecessary cellular mass exponentially increases drag forces within the reproductive tract.
By aggressively discarding the cytoplasm in the form of residual bodies, the gamete achieves an optimal mass – to – propulsion ratio.
This extreme mass reduction ensures that the kinetic energy generated by the mitochondrial sheath is translated directly into forward motility rather than being wasted on displacing inert cellular bulk.
II. The Enzyme Depletion:
The biophysical consequence of this extreme mass reduction is severe and clinically significant. In typical somatic cells, the expansive cytoplasm serves as the primary reservoir for endogenous antioxidant enzymes, most critically Superoxide Dismutase, Catalase, and Glutathione Peroxidase.
These enzymatic networks are the fundamental biochemical circuit breakers that neutralize rogue oxidative radicals.
By physically discarding its cytoplasmic volume, the spermatozoon simultaneously discards its intrinsic biochemical armory, abruptly severing its access to the very enzymatic systems required to manage metabolic exhaust.
III. The Zero – Defense State:
The immediate outcome of this cellular shedding is the creation of a gamete operating in a state of near – zero thermodynamic resistance.
Stripped of its internal defense mechanisms, the mature spermatozoon enters the seminal plasma completely unequipped to neutralize Reactive Oxygen Species autonomously.
It possesses the kinetic machinery to move, but it lacks the biochemical infrastructure to protect that movement, existing on a razor – thin margin of viability where any elevation in oxidative pressure immediately threatens to breach its unprotected perimeter.

2. The Phospholipid Requirement
The Biophysical Demand For Extreme Membrane Fluidity
While the interior architecture is optimized for mass reduction, the exterior boundary – the plasma membrane – must adhere to entirely different physical laws.
The membrane composition is dictated by the absolute necessity of maintaining a highly pliable and responsive structural envelope.
I. The Liquid Crystal Mandate:
The continuous, rapid undulations of the flagellar beat, combined with the catastrophic structural deformation required for the eventual acrosome reaction, place immense biomechanical stress on the cellular boundary.
To survive these physical demands without rupturing, the plasma membrane must perpetually exist in a highly fluid, liquid crystal state.
This specific biophysical phase allows the lipid bilayer to maintain barrier integrity while accommodating rapid topological deformation and lateral receptor migration, a mechanical flexibility that a rigid membrane architecture would immediately inhibit.
II. The PUFA Dependency:
This state of extreme, uninterrupted fluidity cannot be achieved through saturated lipid profiles. It is mechanically engineered through the intense incorporation of massive amounts of Polyunsaturated Fatty Acids, primarily Docosahexaenoic Acid, into the phospholipid bilayer.
The unique molecular geometry of these long – chain polyunsaturated lipids, characterized by multiple carbon – to – carbon double bonds, creates critical spacing within the membrane matrix.
This specific biochemical configuration prevents tight lipid packing, thereby guaranteeing the essential liquid crystal flexibility required for sustained propulsion and terminal fusion.
III. The Peroxidation Target:
The architectural reliance on these flexible lipids introduces a fatal structural flaw.
The very carbon – to – carbon double bonds that grant Docosahexaenoic Acid its necessary fluidity simultaneously make it the primary biochemical target for the lipid peroxidation cascade.
These specific molecular sites are uniquely vulnerable to the extraction of hydrogen atoms by unmitigated Reactive Oxygen Species.
Once initiated, this oxidative chain reaction rapidly degrades the phospholipid architecture, compromising membrane integrity, arresting flagellar motility, and fundamentally destabilizing the gametic envelope.

3. The Biophysical Paradox
The Most Precious Cargo Guarded By The Weakest Structural Defenses
The spermatozoon ultimately represents the absolute limit of cellular specialization, embodying a profound conflict between its payload value and its defensive capabilities.
It is an entity defined by an extreme vulnerability engineered to serve a solitary kinetic objective.
I. The Condensed Chromatin:
The biological payload itself consists of a highly condensed, hyper – compacted nucleus of transcriptionally inactive DNA.
To survive the journey, this genetic material is wrapped tightly around protamines rather than standard histones, forming a dense crystalline core.
However, this crucial genetic archive is physically located mere nanometers from the highly unstable, polyunsaturated – rich plasma membrane, separated only by a minimal, rapidly thinning nuclear envelope and entirely devoid of any protective cytoplasmic buffer.
II. The Internal Inability:
The true peril of this proximity is magnified by the spermatozoon’s operational limitations.
Because the cell has discarded its cytoplasm and compacted its chromatin into a transcriptionally inactive state, it is biologically inert.
It cannot transcribe new messenger RNA, it cannot synthesize replacement antioxidant enzymes, and it possesses severely limited mechanisms to repair its own DNA once the phosphodiester bonds of the genetic backbone are cleaved by penetrating oxidative radicals. The damage sustained during transit is cumulative and largely irreversible.
III. The External Reliance:
Consequently, the male gamete exists as the ultimate biophysical paradox.
It is tasked with carrying the fundamental blueprint of the biological legacy, yet it is completely stripped of the internal machinery necessary to shield it. The cell is utterly dependent on the external environment – specifically the biochemical milieu of the seminal plasma and the availability of exogenous, lipid – soluble modulators – for its thermodynamic shielding.
Without this external defense matrix, the spermatozoon is simply a perfect biological target, kinetically active but structurally fragile, perpetually awaiting an inevitable oxidative trigger.

The Dual Sabotage:
The 15:1 Toxicity Crisis
How The Modern Dietary Lipid Imbalance Competitively Inhibits Membrane Fluidity And Triggers A Localized Oxidative Storm
The baseline biophysical vulnerability of the spermatozoon is a well – documented evolutionary constant, representing a necessary physiological sacrifice engineered for absolute kinetic advantage.
However, the current, ubiquitous rate of systemic reproductive failure across the global population is a strictly modern anomaly.
The fundamental, mechanical trigger for this biological collapse is the modern industrial diet, which is inherently characterized by a profoundly unnatural and highly toxic 15:1 to 20:1 ratio of Omega-6 to Omega-3 polyunsaturated fatty acids.
This severe systemic lipid dysregulation does not merely manifest as vague, generalized physiological distress or poor health. Instead, it executes a precise, mechanically lethal, dual – pathway sabotage directly on the developing spermatozoon.
This pathophysiological process initiates a total structural failure first by physically petrifying the delicate plasma membrane, stripping away its essential kinetic properties, and second by subsequently drowning the already paralyzed cell within an amplified, localized oxidative cascade.
The gamete is simultaneously trapped by its own architecture and chemically dismantled by its environment.

1. The Competitive Enzymatic Inhibition
The Biochemical Hijacking Of The Lipid Synthesis Pathway
To accurately understand the systemic collapse of spermatozoal motility, one must trace the failure back to the initial biochemical bottleneck within the hepatic and testicular lipid metabolism pathways.
The synthesis of structurally critical fatty acids relies upon a highly specific, fundamentally limited enzymatic infrastructure that cannot dynamically expand to accommodate pathological dietary inputs.
A. The Shared Desaturases:
Both the Omega-6 and the Omega-3 polyunsaturated fatty acid metabolic pathways are inextricably linked because they are forced to compete for the exact same rate – limiting enzymes in the liver and within the localized testicular microenvironment.
Specifically, the elongation and desaturation processes required to convert raw precursor lipids into functional cellular building blocks are governed exclusively by Delta-5 and Delta-6 desaturases. These enzymes function as a rigid metabolic tollbooth, capable of processing only a finite molecular volume at any given physiological moment.
B. The 15:1 Overload:
The precise biophysical mechanism of the modern reproductive collapse lies in the strict principle of competitive inhibition.
When the biological system is chronically exposed to a massive 15:1 or even 20:1 surplus of dietary Omega-6 fatty acids, predominantly in the form of Linoleic Acid, this sheer volumetric mass physically overwhelms the shared enzymatic infrastructure.
The Delta-5 and Delta-6 desaturases become entirely saturated by the Omega-6 influx, which effectively and persistently blocks the essential desaturation and elongation of Omega-3 precursors into structurally critical Docosahexaenoic Acid.
C. The Arachidonic Acid Substitution:
The immediate, devastating consequence of this sustained enzymatic blockade is a forced architectural substitution during the late stages of spermiogenesis.
Deprived of the necessary, steady supply of highly kinked and flexible Docosahexaenoic Acid, the testicular microenvironment must synthesize the developing spermatozoal phospholipid bilayer using the only abundant resources available.
It is forced into a disproportionate incorporation of the rigid Omega-6 derivative, Arachidonic Acid. This forced enzymatic substitution permanently alters the biomolecular composition of the gamete before it even fully matures.

2. The Phase Transition To Biological Glass
The Physical Paralysis Of Flagellar Propulsion
The substitution of Arachidonic Acid for Docosahexaenoic Acid within the lipid bilayer is not merely a theoretical chemical variance; it translates directly into a catastrophic physical failure of the cellular machinery.
The spermatozoon relies on extreme membrane fluidity to function, and this chemical shift fundamentally destroys that exact mechanical prerequisite.
A. The Loss Of Molecular Kinks:
The architectural superiority of Docosahexaenoic Acid lies in its highly kinked molecular geometry, created by its specific arrangement of six carbon double bonds, which physically prevents neighboring lipids from packing closely together.
Arachidonic Acid possesses a distinctly different spatial configuration that allows for much tighter lipid packing within the phospholipid bilayer. This structural shift severely reduces the essential free volume between the lipid chains, effectively eliminating the critical spacing that permits fluid mechanical deformation.
B. The Thermodynamic Phase Transition:
As the concentration of Arachidonic Acid artificially rises and the lipid chains pack tightly together, a profound biophysical event occurs across the entire surface of the cell.
The tight lipid packing forces the plasma membrane to undergo a premature thermodynamic phase transition. The cellular boundary rapidly shifts from its necessary, dynamic liquid – crystal state into a rigid, highly inflexible gel – like state.
Functionally, the exterior of the spermatozoon petrifies, transforming into a brittle biological glass that cannot yield or adapt to kinetic force.
C. The Mechanical Arrest:
The structural conclusion of this phase transition is absolute mechanical arrest.
The newly rigid, glass – like membrane physically cannot transmit or accommodate the mechanical kinetic force generated by the internal mitochondrial sheath.
The energy required for propulsion is generated, but it is immediately trapped and suppressed by the structural envelope.
The flagellar beat is completely paralyzed, providing the exact, objective biophysical mechanism for the clinical diagnosis of Asthenozoospermia, rendering the cell mechanically immobile.

3. The Oxidative Amplification
The Localized Generation Of The ROS Crossfire
The structural paralysis of the cell is only the primary phase of the modern lipid toxicity crisis.
The very same Arachidonic Acid that physically petrifies the membrane simultaneously acts as the primary fuel source for a secondary, highly destructive inflammatory crossfire that guarantees absolute cellular death.
A. The Eicosanoid Shift:
The excessive accumulation of Arachidonic Acid within the reproductive tissues serves as a massive, volatile substrate pool for local enzymatic conversion.
Specifically, Cyclooxygenase enzymes, including the highly reactive COX-2, rapidly metabolize this excess Arachidonic Acid, driving both a systemic and localized overproduction of highly potent, pro – inflammatory signaling molecules, most notably Prostaglandin E2.
This rapid eicosanoid shift fundamentally alters the chemical environment of the seminal fluid from a protective biological medium into an active inflammatory combat zone.
B. The Inflammatory ROS Generation:
The continuous presence of artificially elevated Prostaglandin E2 and related inflammatory mediators within the reproductive tract immediately triggers a localized immune response, actively recruiting localized leukocytes to the seminal fluid.
Once deployed and activated by the inflammatory signaling, these white blood cells execute their primary defensive mechanism.
They release massive, uncontrolled bursts of external Reactive Oxygen Species directly into the surrounding seminal plasma, flooding the immediate environment with aggressive oxidative radicals.
C. The Lipid Peroxidation Cascade:
The final, lethal convergence of this dual sabotage occurs when these highly volatile external Reactive Oxygen Species inevitably collide with the already rigid, Arachidonic Acid – compromised spermatozoal membrane. This collision instantly initiates a violent lipid peroxidation cascade.
Because the gamete fundamentally lacks intrinsic antioxidant defenses following cytoplasmic extrusion, this radical chain reaction physically shatters the remaining structural integrity of the membrane, tearing through the lipid bilayer and propagating inward, ultimately cleaving the critical phosphodiester bonds of the unprotected DNA payload.

The Keyora Protocol:
Deploying The Astaxanthin Vanguard And The 2-4:1 Matrix
The Biophysical Integration Of An Absolute Thermodynamic Shield And A Targeted Lipidomic Matrix To Execute A Systemic Reproductive Reboot
The spermatozoon is currently trapped within a relentless, dual – front biological collapse.
On the structural perimeter, its delicate plasma membrane is actively petrified, locked into an inflexible biological glass by the overwhelming, highly toxic 15:1 ratio of dietary Omega-6 to Omega-3 fatty acids.
Simultaneously, its internal architecture, left entirely unprotected and devoid of endogenous enzymatic defenses following the extreme mass reduction of cytoplasmic extrusion, remains under an unchecked, localized oxidative assault from surrounding leukocyte activity.
In this hostile microenvironment, conventional, water – soluble antioxidants consistently fail at the rigid tight junctions of the Blood – Testis Barrier, proving biophysically incapable of reaching the vulnerable gametic payload.
Conversely, isolated or unprotected lipid interventions inevitably undergo rapid lipid peroxidation before ever successfully reaching their intended cellular target, frequently amplifying the very oxidative damage they were administered to modulate.
This uncompromising biophysical reality demands the deployment of a highly engineered, dual – action intervention protocol. It requires an absolute thermodynamic shield capable of completely halting the Reactive Oxygen Species cascade, paired sequentially with a meticulously targeted lipidomic matrix designed to structurally rebuild the gametic architecture.
In the Keyora research paradigm, Astaxanthin enters the theater as the absolute protagonist, a specialized transmembrane modulator, while a highly calibrated 2-4:1 Omega matrix serves as the essential, synergistic structural foundation required to execute a complete, systemic reproductive reboot.

1. The Absolute Protagonist:
The Transmembrane Shield
The Biophysical Deployment Of Astaxanthin Across The Reproductive Barriers
The introduction of Astaxanthin into this failing biological system represents a fundamental shift in cellular defense strategy.
It is not merely an exogenous nutrient; it is a highly specialized marine – derived xanthophyll carotenoid engineered by evolutionary pressure to survive extreme thermodynamic stress. Its deployment is absolute and mechanically precise.
Firstly, The Blood – Testis Barrier Penetration:
The immediate physiological hurdle for any reproductive intervention is the highly selective, tightly regulated network of Sertoli cell tight junctions known as the Blood – Testis Barrier.
Conventional antioxidants are generally hydrophilic and are systematically rejected at this structural border.
Astaxanthin operates under a completely different set of physical laws.
Its extreme lipophilicity allows it to effortlessly dissolve into circulating lipid carriers, seamlessly bypassing the restrictive tight junctions of the Blood – Testis Barrier that block lesser molecules, thereby ensuring successful, undiluted delivery directly into the isolated seminiferous tubules.
Secondly, The 30-Angstrom Anchoring:
The physical insertion of this molecule into the cellular architecture is a masterpiece of biological engineering.
Astaxanthin possesses a highly specific, precise 30-Angstrom molecular length, a dimension that perfectly and exactly matches the width of the hydrophobic core within the spermatozoal and mitochondrial phospholipid bilayers.
Unlike superficial antioxidants that merely float along the exterior membrane surface and are easily dislodged, Astaxanthin utilizes its polar terminal rings to anchor securely to both the inner and outer hydrophilic lipid heads.
It stands vertically across the membrane, forming an immovable structural cross – tie.
Thirdly, The Electron Resonance Quenching:
Once securely anchored across the lipid bilayer, Astaxanthin activates its primary thermodynamic defense mechanism.
It utilizes its extensive, central conjugated polyene chain – a precise sequence of alternating double and single carbon bonds – to directly absorb the volatile kinetic energy of penetrating Reactive Oxygen Species.
When an aggressive oxidative radical strikes the cell, Astaxanthin intercepts the threat, capturing the unpaired electrons and continuously delocalizing them through an infinite resonance loop across its carbon chain, safely dissipating the lethal oxidative energy as harmless, microscopic heat.
Fourthly, The Non-Pro-Oxidant Guarantee:
The most critical biophysical advantage of this specific molecule is an absolute rule of its chemical structure. Because it neutralizes destructive oxidative radicals exclusively through continuous electron resonance rather than through molecular donation or structural degradation, Astaxanthin never undergoes a dangerous phase transition to become a pro-oxidant itself.
Unlike standard interventions that can become toxic once exhausted, Astaxanthin maintains its structural integrity, providing a permanent, impenetrable, and continuously active thermodynamic shield for both the vulnerable DNA payload and the kinetic mitochondrial engines.

2. The Structural Recalibration:
The 2-4:1 Correction
Forcing The Enzymatic Shift To Rebuild The Liquid Crystal Membrane
While the deployment of the transmembrane shield effectively halts the acute, ongoing destruction of the cell, the existing structural damage must be addressed.
The petrified, glass – like membrane cannot propel the gamete; it must be physically and systematically rebuilt from the foundational level using a highly specific influx of biomolecular materials.
Firstly, The Enzymatic Override:
To initiate this structural reconstruction, the Keyora matrix introduces a highly precise, deliberately engineered 2-4:1 ratio of Omega-3 to Omega-6 fatty acids into the systemic circulation.
This calculated volumetric influx is specifically designed to overwhelm the existing competitive inhibition bottleneck occurring at the shared Delta-5 and Delta-6 desaturase enzymes.
By flooding the metabolic pathways with a massive surplus of Omega-3 precursors, the protocol aggressively overrides the pathological lipid synthesis cascade, forcing the enzymatic infrastructure to abandon the production of rigid, pro – inflammatory lipids.
Secondly, The DHA Synthesis Reboot:
This targeted, overwhelming enzyme override forcibly reprograms the localized testicular microenvironment. It explicitly instructs the cellular metabolic machinery to prioritize the continuous, uninterrupted elongation and desaturation of Alpha-Linolenic Acid.
Through this enforced biochemical shift, the biological system is compelled to rapidly synthesize vast, localized quantities of Docosahexaenoic Acid, guaranteeing a steady supply of this highly specialized, functionally critical Omega-3 derivative directly to the seminiferous tubules where the gametes are actively maturing.
Thirdly, The Displacement Of Arachidonic Acid:
As the newly synthesized, highly kinked Docosahexaenoic Acid molecules arrive at the site of spermiogenesis, they initiate a process of physical cellular reconstruction.
The complex molecular geometry of these new lipids allows them to integrate seamlessly into the compromised spermatozoal phospholipid bilayer, forcibly and physically displacing the accumulated, rigid Arachidonic Acid molecules that previously paralyzed the cell. This represents a literal lipidomic swapping out of defective structural components.
Fourthly, The Restoration Of Fluidity:
The ultimate biophysical consequence of this targeted structural lipidomic reconfiguration is the successful restoration of the functional membrane architecture.
By systematically replacing tightly packed, rigid lipids with highly flexible, kinked polyunsaturated chains, the membrane is rescued from its brittle, biological glass state.
It rapidly transitions back to its required, highly dynamic liquid crystal state, successfully re – enabling the rapid topological deformation necessary for powerful, sustained flagellar propulsion and terminal acrosomal fusion.

3. The Synergistic Revival
The Closed – Loop Mechanism Of Protection And Reconstruction
The true efficacy of this protocol does not lie in the isolated actions of its components, but in their meticulously timed, master – subordinate synergy.
The biological system is too hostile for single – vector interventions; it demands a closed – loop mechanism where absolute protection simultaneously facilitates unhindered structural regeneration.
Firstly, The Escorted Transit:
The foundational Omega-3 substrates, specifically Alpha-Linolenic Acid and its derivative Docosahexaenoic Acid, are structurally essential but biochemically highly fragile, possessing multiple double bonds that make them prime targets for premature oxidation.
In this synergistic matrix, Astaxanthin acts as the unyielding vanguard. It physically escorts and shields these circulating lipids from systemic oxidative crossfire during their long transit from the digestive tract, ensuring they arrive at the reproductive barrier completely intact and functionally viable.
Secondly, The Protected Reconfiguration:
Once these delicate lipids successfully penetrate the reproductive tract, the Astaxanthin transmembrane shield secures the highly volatile localized microenvironment.
By continuously quenching the surrounding Reactive Oxygen Species generated by local leukocyte activity, Astaxanthin creates a safe, thermodynamically stable perimeter.
This isolated safe zone allows the 2-4:1 lipid matrix to successfully rebuild the gametic membrane without the newly integrated Docosahexaenoic Acid immediately undergoing a catastrophic lipid peroxidation cascade.
Thirdly, The Comprehensive Reboot:
The final convergence of this master – subordinate relationship results in a comprehensive, closed – loop biological revival.
Astaxanthin provides the absolute, impenetrable thermodynamic defense, while the precise 2-4:1 matrix provides the necessary, highly flexible structural foundation.
Together, operating in perfect biophysical synchronization, they execute a complete, systemic reboot of the spermatozoon, optimizing the cellular architecture to ensure the biological payload survives the hostile transit and fulfills its evolutionary mandate.

4. The Clinical Horizon
Setting The Stage For The Academic Validation Of The Protocol
While the biophysical elegance and mechanical logic of the Keyora protocol are structurally sound, theoretical biochemistry is insufficient for establishing a new standard of reproductive optimization.
This mechanistic foundation must now be subjected to the unforgiving reality of empirical human trials.
Firstly, The Demand For Objective Proof:
In the rigorous Keyora research paradigm, elegant biological models must be explicitly validated by hard, objective clinical metrics.
The theoretical restoration of membrane fluidity and thermodynamic shielding must directly correspond to a statistically significant reduction of seminal Reactive Oxygen Species, the absolute preservation of internal DNA integrity, and a measurable, profound restoration of forward linear velocity in human subjects.
Secondly, The Supreme Academic Tribunal:
To verify these claims, the subsequent analytical chapters will subject this Astaxanthin – driven, lipid – supported protocol to the highest levels of independent academic scrutiny.
We will move beyond the biochemical whiteboard and draw exclusively from top – tier, peer – reviewed journals and gold – standard, double – blind, placebo – controlled clinical trials that have tested these exact molecules against the harshest biological realities of human reproductive failure.
Thirdly, The Ultimate Metric Of Success:
Ultimately, the validation of this biophysical intervention hinges on one definitive, uncompromising clinical outcome.
As we transition into the empirical data, we will examine precisely how the systemic deployment of a transmembrane shield and a targeted lipidomic matrix directly translates into profound, statistically significant, and reproducible surges in actual, real – world conception rates, thereby closing the loop between cellular mechanics and clinical triumph.

Chapter 1: The Competitive Inhibition Crisis:
Halting Arachidonic Acid Overload
How Astaxanthin shielding enables targeted DHA integration to reverse flagellar paralysis.
The spermatozoon must be understood not as a conventional somatic cell, but as a highly specialized, transient biological missile designed for a singular, mechanically demanding trajectory through a hostile reproductive tract. Its kinetic success and ultimate biological viability are entirely predicated on the physical state of its extreme outer boundary – the plasma membrane.
For this specialized gametic cell, the structural envelope cannot function as a static, rigid wall; it must perpetually exist in a highly dynamic, rigorously maintained biophysical state known in molecular thermodynamics as a liquid crystal. This specific phase of matter represents an evolutionary masterpiece of cellular engineering, carefully balancing the absolute structural integrity required to contain the genetic payload with the extreme deformability necessary to navigate fluid resistance.
Without this precisely calibrated liquid crystal state, the spermatozoon cannot initiate the complex, undulating kinematics required for linear velocity, nor can it execute the terminal topological modifications necessary for successful oocyte penetration. It is a biological entity that lives and dies by the mechanical parameters of its external boundary.

1. The Phospholipid Bilayer Architecture
The Structural Foundation Of The Spermatozoal Membrane
To achieve this critical mechanical state, the gamete relies upon a profoundly specific and highly complex biomolecular foundation.
The structural architecture of the spermatozoal membrane represents a radical departure from standard cellular lipid profiles.
I. The Lipid Matrix:
The unique architecture of the spermatozoal phospholipid bilayer is engineered through the precise spatial arrangement of highly specialized amphipathic lipid molecules.
Unlike the membranes of localized tissue cells, which prioritize rigid barrier function and stable receptor localization, the gametic envelope is exceptionally rich in specific lipid classifications designed specifically to lower intermolecular friction and expand the free volume within the hydrophobic core of the bilayer, thereby maintaining its distinct, highly flexible biophysical properties.
II. The PUFA Dependency:
This continuous, low – viscosity microenvironment is not self – sustaining; it is dictated by an absolute, structural dependency on a massive incorporation of Polyunsaturated Fatty Acids into the membrane matrix.
Specifically, Docosahexaenoic Acid, a highly specialized long – chain Omega-3 fatty acid featuring six carbon – to – carbon double bonds, is strategically integrated into the structural envelope.
The extreme molecular kinking of Docosahexaenoic Acid prevents tight lipid packing, physically forcing the phospholipid chains apart and ensuring the membrane retains its necessary, highly pliable liquid crystal characteristics.
III. The Structural Vulnerability:
However, this reliance on extreme structural fluidity introduces a fatal biophysical paradox.
The very multiple double bonds of the Polyunsaturated Fatty Acids required to maintain this vital liquid crystal architecture are inherently highly susceptible to aggressive oxidative damage.
They represent prime targets for free radical extraction, meaning the spermatozoon absolutely requires continuous, robust thermodynamic shielding to prevent the immediate catastrophic degradation of its delicate propulsion envelope.

2. The Flagellar Propulsion Mechanics
The Physical Demands Of The Whipping Motion
The liquid crystal membrane is not merely a protective envelope; it is an active, integrated component of the cellular propulsion system.
The mechanical force generated by the gamete places extraordinary stress on its external perimeter.
I. The Mechanical Force Transmission:
The flagellum, serving as the biological motor of the cell, generates an intense, continuous whipping motion powered by the rapid hydrolysis of ATP within the mitochondrial sheath.
As this motor fires, the kinetic wave propagates down the tail, requiring the overlying plasma membrane to constantly stretch, compress, and instantly recover its precise spatial conformation without a fraction of a millisecond of delay or hesitation.
II. The Elasticity Requirement:
The liquid crystal state provides the extreme physical elasticity necessary to accommodate this violent mechanical activity.
Because the lipid molecules within a fluid membrane can rapidly shift and migrate laterally, the structural envelope can seamlessly transmit the immense mechanical force generated by the axoneme directly against the surrounding fluid medium without causing the delicate membrane itself to rupture, shear, or undergo catastrophic mechanical failure.
III. The Propulsion Failure:
Consequently, any localized or systemic loss of this specific membrane fluidity directly translates to an immediate, mechanical inability to propagate the necessary flagellar wave.
If the lipid bilayer petrifies or stiffens, the kinetic energy generated by the mitochondria becomes trapped within the rigid envelope.
The whipping motion is blunted and eventually paralyzed, stalling the spermatozoon entirely in its tracks and manifesting clinically as a profound collapse in forward progressive motility.

3. The Acrosome Reaction Deformation
The Topological Requirements For Oocyte Fusion
Beyond the mechanics of linear propulsion, the plasma membrane must retain its hyper – fluid state to successfully execute the terminal phase of its biological mission.
The act of fertilization demands an extreme, rapid localized structural disassembly.
I. The Receptor Clustering:
Upon physically reaching the zona pellucida of the oocyte, the spermatozoal membrane must immediately facilitate the activation of a complex biochemical cascade.
This requires the membrane to allow specific binding receptors to rapidly drift through the lipid bilayer and cluster together into highly concentrated, localized functional domains, a lateral molecular migration that is entirely dependent on a low – viscosity, fluid membrane environment.
II. The Membrane Fusion:
Once bound, the cell initiates the acrosome reaction, a violent exocytotic event governed by exact biophysical laws.
This reaction requires the outer plasma membrane of the spermatozoon to undergo extreme, rapid topological deformation in order to physically invaginate and fuse seamlessly with the underlying outer acrosomal membrane, thereby releasing the highly concentrated proteolytic enzymes required to digest the protective barrier of the oocyte.
III. The Fluidity Mandate:
This rapid, catastrophic structural merging is physically and mechanically impossible if the localized membrane architecture is rigid or compromised.
The biological process absolutely mandates the highly fluid, hyper – responsive liquid crystal state maintained exclusively by optimal, unoxidized levels of Docosahexaenoic Acid.
Without this precise lipidomic foundation, the gamete may reach its target but will ultimately fail to breach the perimeter, rendering its entire biological journey mechanically futile.

1.1 The Competitive Inhibition Crisis:
The 15:1 Enzymatic Hijack
How The Modern Dietary Lipid Imbalance Physically Blocks DHA Synthesis And Forces The Incorporation Of Rigid Structural Lipids
The pristine, highly calibrated liquid crystal architecture of the spermatozoon is currently under a relentless and systemic biological attack.
The primary weapon in this modern reproductive crisis is not an exogenous viral pathogen or an acute chemical toxin, but rather a severe, chronic macronutrient dysregulation: the highly anomalous modern dietary ratio of 15:1 to 20:1 of Omega-6 to Omega-3 polyunsaturated fatty acids.
This profound systemic imbalance does not merely alter general metabolic health; it triggers a specific, catastrophic biochemical event within the reproductive axis known in enzymatic kinetics as competitive inhibition.
By physically overwhelming the highly specialized lipid synthesis machinery, this dietary ratio fundamentally sabotages the delicate enzymatic pathways required to build a functional, mechanically viable spermatozoal membrane, silently neutralizing the biological missile before it ever leaves the testicular microenvironment.

1. The Shared Desaturase Bottleneck
The Enzymatic Choke Point Of Lipid Synthesis
To comprehend the exact mechanics of this architectural failure, we must first isolate the precise biochemical vulnerability within the lipid synthesis pathway.
The cellular construction of a specialized gametic membrane relies upon a highly restricted and highly contested enzymatic corridor.
A. The Precursor Competition:
The fundamental building blocks of the cellular membrane, specifically the parent Omega-6 molecule known as Linoleic Acid and the parent Omega-3 molecule known as Alpha-Linolenic Acid, share a critical biological limitation: they cannot be synthesized endogenously by the human body.
They are strictly essential fatty acids that must be acquired entirely through dietary intake, meaning the physiological baseline of the organism is entirely at the mercy of exogenous nutritional inputs.
B. The Enzymatic Pathway:
Once these raw precursor lipids enter the systemic circulation, they are biologically inert in their foundational state.
To become the highly complex, structurally functional lipids required by the spermatozoon, both the Omega-6 and Omega-3 precursors must utilize the exact same metabolic machinery located primarily within the hepatic tissue and secondarily within the localized testicular microenvironment, setting the stage for a critical metabolic bottleneck.
C. The Delta-5 And Delta-6 Desaturases:
This shared elongation and desaturation pathway is strictly governed by a highly finite, rate – limiting set of enzymes, specifically identified as the Delta-5 and Delta-6 desaturases.
These precise biological catalysts are responsible for systematically inserting vital carbon double bonds into the fatty acid chains, a required modification that progressively increases the physical flexibility and structural complexity of the developing lipid molecules.
D. The Affinity Dynamics:
Under optimal, evolutionary conditions, these specific desaturase enzymes actually possess a higher intrinsic chemical affinity for the Omega-3 substrates.
However, their functional output is not determined solely by chemical preference; it is entirely and ruthlessly dictated by the volumetric substrate concentration ratio presented to their active receptor sites at any given physiological moment.

2. The 15-20:1 Systemic Overload
The Concentration – Driven Hijacking Of The Metabolic Machinery
The modern physiological environment represents a radical departure from the baseline parameters under which these desaturase enzymes evolved to operate.
The current systemic lipid profile actively weaponizes the concentration – dependent nature of this metabolic machinery.
A. The Dietary Shift:
The rapid industrialization of the global food supply, characterized by the ubiquitous extraction and consumption of processed seed oils, has relentlessly saturated the systemic human circulation with an unprecedented volume of Omega-6 fatty acids.
This aggressive dietary shift has violently pushed the baseline circulating physiological ratio from a balanced, evolutionary standard of 2-4:1 directly into a highly toxic, metabolically disruptive 15-20:1 imbalance.
B. The Substrate Flooding:
As this massive, disproportionate concentration of dietary Omega-6 enters the hepatic and testicular processing centers, it physically floods the microscopic active sites of the shared Delta-5 and Delta-6 desaturase enzymes.
The sheer volumetric density of the Omega-6 molecules creates an insurmountable spatial barrier around the enzymatic infrastructure, fundamentally altering the operational capacity of the metabolic pathway.
C. The Competitive Inhibition Mechanism:
This intense molecular crowding triggers the objective biochemical mechanism of competitive inhibition.
Because the desaturase enzymes can only bind and process one single lipid molecule at any given microsecond, the overwhelming numerical superiority of the Omega-6 substrates physically blocks the scarce Omega-3 substrates from ever accessing the enzymatic active sites, effectively rendering the Omega-3 molecules metabolically invisible.
D. The Enzymatic Hijack:
The ultimate consequence of this persistent steric hindrance is that the body’s metabolic machinery is effectively hijacked.
The finite desaturase enzymes are forcefully commandeered and mandated to almost exclusively process Omega-6 lipids, operating entirely blindly to the actual, highly specific structural lipid requirements of the rapidly developing spermatozoal membranes within the seminiferous tubules.

3. The DHA Synthesis Blockade
The Critical Starvation Of Membrane – Fluidizing Lipids
When the enzymatic infrastructure is commandeered by the Omega-6 pathway, the parallel Omega-3 processing line suffers an immediate and devastating collapse.
This biochemical cessation fundamentally starves the reproductive tissues of their most vital architectural components.
A. The Omega-3 Starvation:
The direct and immediate consequence of this enzymatic hijack is severe Omega-3 starvation.
The necessary downstream conversion of the foundational Alpha-Linolenic Acid into its longer, highly specialized functional derivatives is drastically suppressed, completely cutting off the localized supply chain to the maturing germ cells.
B. The EPA Deficit:
The first casualty of this metabolic blockade is a precipitous drop in the synthesis of Eicosapentaenoic Acid.
This severe EPA deficit abruptly removes a crucial, highly potent anti – inflammatory signaling molecule from the localized testicular microenvironment, stripping away a foundational layer of biochemical protection and leaving the reproductive tissues highly susceptible to baseline oxidative stress.
C. The Ultimate DHA Blockade:
The most critical, mechanically lethal structural failure occurs at the absolute terminus of the pathway: the systemic synthesis of Docosahexaenoic Acid is virtually halted.
Docosahexaenoic Acid, possessing six carbon double bonds, is the maximally kinked, highly specialized lipid that is absolutely and uncompromisingly required to establish and maintain the extreme membrane fluidity necessary for spermatozoal propulsion.
D. The Structural Void:
Because the synthesis of Docosahexaenoic Acid is suppressed by competitive inhibition, the developing spermatozoon is left with a massive, critical structural void within its phospholipid bilayer blueprint.
It is systematically deprived of the precise molecular components mechanically required to construct its dynamic, yielding liquid crystal state, leaving the cellular boundary dangerously incomplete.

4. The Arachidonic Acid Flooding
The Forced Incorporation Of Rigid Structural Components
Biological systems abhor a structural vacuum.
When the necessary optimal components are systematically withheld from a rapidly dividing cell, the biological machinery is forced to construct the cellular perimeter using sub – optimal, mechanically restrictive surrogate materials.
A. The Omega-6 Downstream Surge:
As the hijacked Delta-5 and Delta-6 desaturases continuously and exclusively process the massive Omega-6 payload, they inevitably produce a massive, unregulated overabundance of elongated downstream Omega-6 metabolites.
The lipid synthesis pathway becomes an unchecked manufacturing line for pro – inflammatory and structurally rigid lipid variants.
B. The Arachidonic Acid Accumulation:
The specific, catastrophic result of this unregulated manufacturing surge is the massive physiological accumulation of Arachidonic Acid.
This dense, structurally rigid Omega-6 derivative floods both the systemic circulation and, most critically, pools heavily within the highly sensitive localized testicular microenvironment, completely altering the available building materials for spermatogenesis.
C. The Forced Membrane Integration:
The developing spermatozoon, actively undergoing spermiogenesis but critically starved of the necessary Docosahexaenoic Acid, is confronted with a biological mandate: it must seal its cellular boundary to survive.
To complete its phospholipid bilayer, the cell is physically forced to incorporate this overwhelming excess of available, structurally rigid Arachidonic Acid directly into its outer membrane matrix.
D. The Architectural Sabotage:
This forced molecular substitution is nothing less than an act of complete cellular architectural sabotage.
By aggressively replacing the highly flexible, heavily kinked Docosahexaenoic Acid components with the stiff, tightly packing structure of Arachidonic Acid, the delicate gametic membrane is forcefully petrified, effectively setting the precise biophysical stage for complete mechanical failure and subsequent flagellar paralysis.

1.2 The Phase Transition:
Membrane Petrification
The Biophysical Transformation From A Dynamic Liquid Crystal To A Rigid Biological Glass And The Resulting Mechanical Paralysis
The forced incorporation of Arachidonic Acid in place of Docosahexaenoic Acid during the critical terminal phases of spermiogenesis is not merely a superficial chemical substitution; it represents a profound and catastrophic biophysical degradation of the gametic envelope.
This specific lipidomic shift fundamentally alters the underlying molecular geometry of the spermatozoal plasma membrane, removing the necessary structural spacing required for fluid motion.
By systematically replacing highly flexible, heavily kinked molecular components with stiff, structurally restrictive alternatives, this metabolic substitution directly triggers a severe, localized thermodynamic phase transition across the cellular boundary.
This biophysical state change effectively petrifies the outer envelope of the cell, locking the membrane into a rigid matrix that mechanically disables its kinetic propulsion system.
The spermatozoon becomes physically imprisoned within its own compromised architecture, entirely unable to translate metabolic energy into the forward velocity required to navigate the reproductive tract.

1. The Loss Of Molecular Kinks
The Geometric Consequences Of Lipid Substitution
To understand the mechanical failure of the membrane, one must analyze the precise spatial architecture of the lipid molecules at a microscopic level, carefully comparing the optimal structural components against the pathological substitutes forced into the cellular boundary by systemic competitive inhibition.
Firstly, The DHA Geometry:
Docosahexaenoic Acid represents the absolute pinnacle of membrane – fluidizing biological engineering.
Its specific molecular structure, characterized by an elongated carbon chain punctuated by six distinct carbon – to – carbon double bonds, creates multiple, severe structural kinks along the entire length of the lipid.
These aggressive geometric deviations generate massive steric hindrance within the hydrophobic core of the phospholipid bilayer, effectively functioning as internal molecular spacers that physically and persistently prevent adjacent lipid chains from packing closely together.
Secondly, The Arachidonic Acid Linearity:
This optimal, highly fluid architecture is immediately compromised when Arachidonic Acid is forcefully substituted into the membrane matrix.
While Arachidonic Acid is technically classified as a polyunsaturated fatty acid, its structural reality is vastly different from its Omega-3 counterpart.
Possessing only four double bonds across a slightly shorter carbon backbone, its resulting molecular geometry is significantly more linear and considerably less kinked compared to the highly coiled structure of Docosahexaenoic Acid, a difference that drastically reduces its overall steric volume and spatial profile.
Thirdly, The Tight Packing Phenomenon:
The physical result of this structural substitution is an immediate and devastating architectural collapse within the lipid bilayer.
As the considerably more linear Arachidonic Acid molecules systematically replace the highly kinked Docosahexaenoic Acid, the natural spatial buffers and steric hindrances are entirely removed.
Without the necessary geometric spacing to hold them apart, the lipid molecules are able to associate and pack much more tightly together, completely eliminating the crucial free intermolecular space and low – viscosity environment that the spermatozoon strictly requires to maintain vital membrane fluidity.

2. The Shift To Biological Glass
The Thermodynamic Phase Transition Of The Membrane
The critical elimination of free space within the membrane matrix is not simply a crowding issue; it crosses a definitive thermodynamic threshold that fundamentally alters the physical state of the cellular envelope, shifting it from a yielding, responsive liquid to a restrictive, petrified solid.
Firstly, The Loss Of The Liquid Crystal:
The continuous, high – velocity mechanical demands of the spermatozoon dictate that its membrane must always exist in a dynamic, highly disordered liquid crystal state.
This specific biophysical configuration allows individual lipid molecules to slide past one another effortlessly, accommodating rapid changes in cellular shape and surface topology.
However, the tight intermolecular packing induced by the overwhelming presence of Arachidonic Acid physically destroys this necessary disorder, severely restricting lateral molecular movement and forcing the membrane out of its functional liquid crystal equilibrium.
Secondly, The Phase Transition Mechanism:
As lateral molecular movement is restricted and the linear lipid chains lock tightly together, a catastrophic thermodynamic event occurs across the entire surface area of the gamete. The membrane undergoes a distinct, localized phase transition.
Stripped of its internal kinetic spacing and structural flexibility by the dense packing of substituted lipids, the phospholipid bilayer rapidly shifts from its necessary fluid state into a highly ordered, rigid, and structurally stiff configuration recognized biophysically as a gel phase.
Thirdly, The Petrification Outcome:
The final biophysical reality of this thermodynamic phase transition is absolute structural lockdown.
The spermatozoal membrane has been completely and systematically petrified by the systemic dietary lipid imbalance.
It has transitioned from a highly adaptive, responsive fluid envelope into a brittle, inflexible cellular boundary – effectively forming a continuous, restrictive biological glass around the gamete that fundamentally cannot yield, stretch, or deform under intense mechanical stress.

3. The Paralysis Of Asthenozoospermia
The Mechanical Failure Of The Flagellar Engine
The formation of this rigid biological glass over the surface of the cell has an immediate, inescapable, and dire consequence for gametic motility.
The structural petrification of the exterior boundary directly translates into the functional paralysis of the internal kinetic machinery.
Firstly, The Energy Transfer Failure:
The forward trajectory of the spermatozoon is generated by the internal mitochondrial sheath, which rapidly hydrolyzes ATP to drive the continuous, undulating whipping motion of the underlying flagellar axoneme.
However, when the overlying plasma membrane is locked into a rigid biological glass state, it cannot elastically deform to accommodate this rapid internal structural shifting.
The stiffened envelope actively resists the kinetic wave, resulting in a profound energy transfer failure where the immense mechanical force of the motor is completely suppressed by the unyielding external boundary.
Secondly, The Brittle Micro – Fractures:
When the internal mitochondrial engine forces intense, rapid movement against this unyielding, rigid gel phase, the mechanical conflict creates immense and unsustainable shear stress across the surface of the entire cell.
Because the petrified membrane can no longer stretch or bend smoothly, this forced kinetic activity leads to microscopic brittle fractures within the lipid architecture, causing the rapid dissipation of vital metabolic energy and inducing severe, localized structural damage to the gametic envelope.
Thirdly, The Clinical Manifestation:
Ultimately, this biophysical membrane petrification serves as the exact, objective mechanical root cause of Asthenozoospermia.
The affected spermatozoon is not necessarily necrotic or entirely devoid of metabolic energy; rather, it is physically and completely paralyzed by its own structural rigidity.
Trapped within a hardened, glassy shell synthesized from the wrong metabolic precursors, the cell is mechanically immobilized, rendering its genetic payload completely non – viable for the extreme physical demands of fertilization.

1.3 The Astaxanthin Vanguard:
Deploying The Transmembrane Shield
The Biophysical Deployment Of The Absolute Thermodynamic Protagonist To Breach The Testicular Barrier, Anchor The Membrane, And Quench The Localized Oxidative Storm
The structural integrity of the developing spermatozoon has already been severely compromised by the pathological 15:1 systemic lipid ratio, effectively locking its vital propulsion envelope into a rigid, paralyzed state.
However, a secondary, equally lethal and immediate threat now looms over the immobilized cell. This rigid, Arachidonic Acid – heavy membrane, trapped within the localized inflammatory microenvironment of the reproductive tract, is now a prime, highly vulnerable target for aggressive lipid peroxidation initiated by localized bursts of external Reactive Oxygen Species.
If this oxidative cascade is allowed to propagate, it will rapidly shatter the remaining cellular boundary and cleave the unprotected genetic payload. Therefore, before any attempt can be made to structurally rebuild the membrane, the perimeter must first be absolutely secured.
Conventional, hydrophilic dietary antioxidants consistently fail in this critical mission, as their water – soluble nature physically repels them from penetrating the strict, lipid – based reproductive barriers.
This escalating biophysical crisis demands the immediate deployment of a highly specific, aggressively lipophilic intervention capable of establishing an absolute, impenetrable thermodynamic shield directly within the cellular envelope.
In this precise capacity, Astaxanthin now enters the structural battlefield as the undisputed, absolute biological protagonist of the Keyora protocol.

1. The Blood – Testis Barrier Breach
Overcoming The Anatomical Isolation Of The Reproductive Tract
The initial and often insurmountable hurdle for any systemic therapeutic intervention targeting spermatogenesis is the profound anatomical isolation of the reproductive microenvironment.
The gametes develop behind one of the most formidable structural borders in the human body.
I. The Hydrophilic Exclusion:
The seminiferous tubules, where the spermatozoa physically mature, are strictly guarded by the Blood – Testis Barrier.
This highly specialized, selectively permeable boundary is composed of an interlocking network of dense tight junctions formed between adjacent Sertoli cells. This structure acts as a rigorous physiological filter, specifically designed to exclude systemic pathogens and large macromolecules.
Crucially, it utilizes these tight junctions to physically and efficiently block the vast majority of circulating, water – soluble, hydrophilic antioxidants – such as Vitamin C and basic bioflavonoids – from ever entering the inner sanctuary of the tubules.
II. The Lipophilic Supremacy:
Astaxanthin fundamentally circumvents this structural blockade through its inherent, extreme chemical lipophilicity.
As a highly specialized, fat – soluble xanthophyll carotenoid synthesized by microalgae to survive intense environmental desiccation, its fundamental chemical nature is perfectly adapted for lipid environments.
This characteristic allows the Astaxanthin molecule to seamlessly dissolve into circulating high – density and low – density lipoprotein carriers within the bloodstream, perfectly camouflaging itself within the body’s natural lipid transport network and effortlessly navigating the hydrophobic circulatory pathways.
III. The Barrier Penetration:
It is precisely this extreme lipophilic advantage that enables Astaxanthin to successfully execute its primary tactical objective.
By traveling within the lipid transport vehicles, the molecule effortlessly slips through the lipid – rich tight junctions of the Blood – Testis Barrier.
It is not repelled by the hydrophobic gating mechanism; instead, it utilizes it as a direct transit corridor, successfully crossing the threshold and achieving exceptionally high, therapeutic concentrations deep within the previously isolated testicular microenvironment.
IV. The Cellular Infiltration:
Upon successfully breaching the macroscopic barrier and reaching the immediate vicinity of the developing spermatozoa, Astaxanthin executes its final infiltration maneuver.
Because of its extreme affinity for lipid structures, it does not require the presence of specialized, rate – limiting membrane transport proteins to enter the cell.
The molecule simply and directly partitions itself into the dense, lipid – rich matrix of the spermatozoal plasma membrane, seamlessly inserting itself into the very structure it is deployed to defend.

2. The 30-Angstrom Structural Integration
The Precise Physical Anchoring Of The Molecular Strut
The physical presence of Astaxanthin within the membrane is not a chaotic or superficial association.
The molecule is a masterpiece of evolutionary engineering, designed to integrate into the phospholipid bilayer with exact, mathematical precision.
I. The Dimensional Perfection:
The true structural brilliance of the Astaxanthin molecule lies in its exact biophysical dimensions.
The linear backbone of the molecule possesses a precise, unvarying length of approximately 30 Angstroms.
This specific measurement is not arbitrary; it is an absolute, perfect spatial match for the exact cross – sectional width of the internal hydrophobic core of both the spermatozoal and mitochondrial phospholipid bilayers.
II. The Polar Anchoring Mechanism:
To secure this perfect spatial alignment, Astaxanthin utilizes specialized structural modifications at both ends of its carbon chain.
The hydroxyl and keto groups located specifically on its terminal ionone rings are strongly hydrophilic.
As the molecule slides into the membrane, these polar functional groups act as chemical anchors, physically locking onto the polar phosphate lipid heads on both the intracellular and extracellular surfaces of the membrane simultaneously, ensuring the molecule cannot be easily dislodged.
III. The Hydrophobic Spanning:
Anchored firmly at both polar boundaries, the long, central conjugated polyene chain of the Astaxanthin molecule spans the entire highly volatile, hydrophobic interior of the membrane.
Unlike shorter antioxidants that embed only halfway, or superficial antioxidants that merely skim the surface, Astaxanthin creates a continuous, unbroken physical bridge directly across the thickness of the entire lipid bilayer.
IV. The Transmembrane Stabilization:
The direct biophysical conclusion of this precise, spanning orientation is that Astaxanthin functionally acts as molecular structural rebar.
Even within a membrane that has been previously compromised and rendered artificially rigid by forced Arachidonic Acid substitution, the vertical integration of Astaxanthin physically stabilizes the disorganized lipid matrix.
It provides a vertical scaffold that significantly increases the membrane’s overall mechanical tolerance to localized shear stress and kinetic force.

3. The Thermodynamic ROS Quenching
The Absolute Neutralization Of The Oxidative Assault
Structural stabilization is merely the secondary benefit of this molecular insertion.
The primary, mission – critical function of Astaxanthin is the immediate and absolute neutralization of the incoming oxidative crossfire targeting the cellular boundary.
I. The Conjugated Electron Cloud:
The immense defensive power of Astaxanthin is derived entirely from the unique chemical architecture of its central spanning region. Its massive carbon chain consists of a highly specific sequence of alternating double and single carbon – to – carbon bonds.
This precise configuration creates a massive, highly active, delocalized electron cloud that continuously resonates along the entire 30-Angstrom length of the molecule, effectively erecting an energetic forcefield throughout the interior of the membrane.
II. The Interception Of Radicals:
This continuous, highly active electron cloud acts as a perfectly positioned, highly efficient thermodynamic trap.
When localized bursts of Reactive Oxygen Species – specifically hyper – aggressive variants like singlet oxygen and hydroxyl radicals – attempt to penetrate the lipid bilayer to reach the genetic payload, they must physically pass through the hydrophobic core.
In doing so, they inevitably collide with the Astaxanthin electron cloud and are immediately physically intercepted.
III. The Resonance Dissipation:
The biophysics of this quenching mechanism is what elevates Astaxanthin above all other biological defenses.
When it intercepts a high – energy radical, the Astaxanthin molecule rapidly absorbs the destructive, volatile oxidative energy.
However, it does not hold this energy or shatter under the stress. Instead, it instantly and safely dissipates the trapped kinetic energy as low – grade, harmless microscopic heat through continuous, unbroken electron resonance back and forth along its carbon chain, leaving its own molecular bonds entirely intact.
IV. The Zero – Phase – Transition Rule:
This mechanism establishes the absolute thermodynamic guarantee of the Keyora protocol.
Because Astaxanthin neutralizes and dissipates radical energy exclusively through continuous internal resonance rather than through the dangerous donation of an electron, it is chemically impossible for the molecule to ever undergo a phase transition to become a damaging pro – oxidant itself.
It remains an impenetrable, permanent thermodynamic shield, perpetually active and never degrading into a threat.

4. The Protection Of Vulnerable Lipids
Halting The Lipid Peroxidation Cascade At The Boundary
The immediate, localized consequence of this thermodynamic quenching is the absolute preservation of the remaining structural lipids within the compromised cellular envelope.
I. The PUFA Proximity:
By precisely anchoring directly within the hydrophobic core of the phospholipid bilayer, Astaxanthin strategically positions its protective electron cloud in immediate, intimate physical proximity to the most vulnerable structural components of the membrane: the multiple carbon – to – carbon double bonds of the existing Polyunsaturated Fatty Acids.
II. The Chain – Breaking Action:
Functioning from this entrenched position, Astaxanthin acts as a potent, immediate chain – breaking antioxidant.
By intercepting the initial Reactive Oxygen Species attack the very millisecond it breaches the outer polar boundary, Astaxanthin decisively prevents the aggressive radicals from executing the abstraction of vulnerable hydrogen atoms directly from the neighboring lipid carbon chains.
III. The Peroxidation Blockade:
This rapid, targeted molecular interception physically and immediately halts the initiation, and entirely prevents the subsequent explosive propagation, of the lipid peroxidation cascade.
By severing the radical chain reaction at its exact point of origin, Astaxanthin completely blocks the downstream generation of highly toxic, DNA – cleaving aldehyde byproducts, most notably the highly destructive Malondialdehyde.
IV. The Preservation Of The Boundary:
The ultimate, clinically vital conclusion of this specific protective action is the absolute preservation of the remaining structural integrity of the spermatozoal membrane.
By shielding the lipids from oxidative shattering, Astaxanthin prevents the already compromised, rigidified cell from physically rupturing under localized oxidative stress, ensuring the gamete remains structurally intact, albeit still mechanically paralyzed.

5. The Prerequisite For Reconfiguration
Securing The Biochemical Safe Zone For Structural Repair
The deployment of Astaxanthin is not the final therapeutic step; rather, it is the absolute, non – negotiable prerequisite necessary to allow the subsequent structural reconstruction of the spermatozoon to commence without immediate failure.
I. The Futility Of Unshielded Repair:
The critical biophysical reality of this protocol is that attempting to repair the petrified membrane by simply introducing high volumes of structurally optimal, highly kinked Omega-3 lipids – specifically Docosahexaenoic Acid – directly into an active, uncontrolled oxidative storm is biologically futile.
Because these required lipids are densely packed with highly vulnerable double bonds, they will be instantly targeted and violently oxidized the moment they arrive, failing to integrate and merely adding volatile fuel to the localized inflammatory fire.
II. The Establishment Of The Safe Zone:
Therefore, the success of the repair relies entirely on the prior deployment of the Astaxanthin transmembrane shield.
By successfully neutralizing the localized leukocyte – driven oxidative threat and extinguishing the lipid peroxidation cascade, Astaxanthin actively establishes and rigorously maintains a strictly controlled, thermodynamically secure biochemical safe zone directly around and within the maturing gametes.
III. The Vanguard Function:
In this highly synchronized intervention, Astaxanthin fulfills its precise role as the biological vanguard.
It does not actively synthesize new lipids, nor does it physically build the flexible membrane itself.
Its singular, vital function is to secure the perimeter, hold the line against oxidative degradation, and guarantee that the incoming, highly fragile structural materials can survive their transit and subsequent cellular integration.
IV. The Handoff To The Lipid Matrix:
This concludes the absolute protagonist’s primary arc within the Keyora intervention timeline.
With the impenetrable thermodynamic shield now firmly locked in place across the cellular boundary, and the localized oxidative fire permanently extinguished, the biological stage is now perfectly and securely set.
The environment is stabilized, allowing the precise 2-4:1 Keyora lipidomic matrix to safely enter the theater and formally execute the profound physical reconstruction of the spermatozoal membrane.

1.4 The Keyora 2-4:1 Matrix:
The Lipidomic Reconfiguration
How The Targeted Omega Matrix Overrides Competitive Inhibition, Displaces Rigid Lipids, And Physically Rebuilds The Liquid – Crystal Membrane Under The Astaxanthin Shield
The deployment of the Astaxanthin vanguard has successfully secured the highly vulnerable testicular microenvironment, establishing an absolute, impenetrable thermodynamic shield against the localized Reactive Oxygen Species crossfire.
However, despite this monumental defensive victory, the spermatozoal membrane itself remains fundamentally petrified. It exists as a brittle, compromised biological glass, its phospholipid bilayer dominated by the rigid, linear geometry of accumulated Arachidonic Acid.
The cell is protected from oxidative shattering, yet it remains mechanically paralyzed. To restore the vital kinetic mechanics of flagellar propulsion, the structural envelope must be physically and aggressively rebuilt from the foundational level.
Operating entirely under the continuous, unyielding protection of the Astaxanthin thermodynamic shield, the Keyora matrix now systematically deploys its highly calibrated 2-4:1 lipidomic payload.
This precise biochemical intervention is engineered to forcibly override the existing enzymatic hijack, effectively reversing the competitive inhibition at the desaturase level and executing a complete, systemic structural reconfiguration of the gametic perimeter.

1. The 2-4:1 Enzymatic Override
Breaking The Competitive Blockade At The Desaturase Level
The physical reconstruction of the cellular boundary cannot commence until the pathological metabolic blockade is completely dismantled.
The biological system must be forcefully instructed to alter its lipid processing priorities.
A. The Precision Ratio:
The Keyora protocol introduces a meticulously engineered 2-4:1 ratio of Omega-6 to Omega-3 fatty acids into the systemic circulation.
This specific formulation is not a random amalgamation of nutritional components; it is a highly precise, biologically targeted mathematical balance explicitly designed to mimic the pristine, evolutionary physiological ideal that the human metabolic infrastructure was originally constructed to process.
B. The Substrate Saturation:
By systematically delivering this targeted lipidomic ratio, the intervention intentionally floods both the hepatic processing centers and the localized testicular microenvironment with a massive, concentrated influx of the parent Omega-3 molecule, Alpha-Linolenic Acid.
This deliberate substrate saturation is designed to drastically alter the localized volumetric availability of metabolic precursors.
C. The Competitive Reversal:
The fundamental biochemical mechanism driving this intervention is the absolute reversal of competitive inhibition.
Driven by the sheer volumetric density of the new Omega-3 payload, the massive influx of Alpha-Linolenic Acid physically outcompetes and displaces the toxic, ambient 15:1 Omega-6 load, forcefully seizing access to the critical active binding sites of the shared Delta-5 and Delta-6 desaturase enzymes.
D. The Pathway Unlocked:
The immediate conclusion of this metabolic siege is that the pathological competitive inhibition is successfully overridden and definitively broken.
The shared metabolic machinery is forcibly commandeered by the Keyora matrix, compelling the enzymatic infrastructure to immediately abandon the unchecked overproduction of rigid Omega-6 metabolites and actively resume the vital, high – volume processing of Omega-3 substrates.

2. The Resurgence Of DHA Synthesis
Generating The Ultimate Fluidizing Lipid
With the enzymatic tollbooth finally cleared of its pathological obstruction, the dormant Omega-3 metabolic pathway is violently reactivated.
This initiates a critical manufacturing cascade aimed at producing the ultimate structural component.
A. The ALA Elongation:
The downstream cascade rapidly accelerates as the newly unblocked Delta-5 and Delta-6 desaturase enzymes process the influx of Alpha-Linolenic Acid.
The metabolic pathway actively elongates and desaturates these parent molecules, systematically adding carbon atoms and critical double bonds to construct progressively more complex, highly flexible lipid structures specifically designated for gametic architecture.
B. The Safe Synthesis:
Crucially, this intricate and delicate metabolic manufacturing process does not occur in a volatile vacuum.
Guarded continuously by the systemic, transmembrane presence of the Astaxanthin shield, this highly vulnerable synthesis phase proceeds flawlessly, guaranteeing that the newly forming, double – bond – rich polyunsaturated fatty acids are not prematurely shattered by background oxidative stress before they can be safely deployed.
C. The DHA Production Spike:
The direct, measurable outcome of this protected enzymatic resurgence is profound.
The localized testicular microenvironment experiences a massive, sustained spike in the biological synthesis of Docosahexaenoic Acid, representing the exact, highly kinked molecular architecture strictly required to establish and maintain extreme spermatozoal membrane fluidity.
D. The Structural Reservoir:
The ultimate conclusion of this manufacturing phase is that the rapidly developing spermatozoa now possess uninterrupted access to an abundant, pristine, and thermodynamically secure structural reservoir.
The local biological system is heavily stockpiled with the highly specialized, severely kinked lipid building blocks essential for constructing a mechanically viable propulsion envelope.

3. The Physical Displacement Of Arachidonic Acid
The Mechanical Eviction Of The Structural Poison
Possessing the correct molecular building materials is only half of the biophysical equation.
The newly synthesized components must now be aggressively integrated into the existing cellular architecture to correct the structural failure.
A. The Lipid Turnover:
The spermatozoal plasma membrane, despite its petrified state, is never entirely static during its developmental phase within the seminiferous tubules.
It inherently exists in a state of continuous, dynamic turnover, perpetually and actively exchanging individual lipid molecules with the surrounding fluid medium of its highly controlled microenvironment.
B. The DHA Integration:
Exploiting this natural mechanism of continuous lipid turnover, the newly synthesized and heavily protected Docosahexaenoic Acid molecules are drawn from the localized reservoir and actively, systematically incorporated directly into the compromised spermatozoal phospholipid bilayer, officially initiating the physical process of cellular reconstruction.
C. The Mechanical Eviction:
This integration triggers a direct biophysical conflict within the membrane matrix.
As the immensely bulky, highly kinked Docosahexaenoic Acid molecules force themselves into the cellular boundary, their massive steric volume physically forces out and aggressively displaces the existing rigid, tightly packed Arachidonic Acid molecules that previously paralyzed the structure.
D. The Removal Of The Glass:
The necessary biophysical conclusion of this continuous molecular exchange is that the restrictive structural poison is systematically and relentlessly purged from the gametic architecture.
The forced eviction of Arachidonic Acid physically dismantles the restrictive parameters of the lipid matrix, effectively shattering the brittle biological glass state from the inside out.

4. The Return Of The Liquid Crystal
The Biophysical Restoration Of Flagellar Mechanics And Acrosomal Competence
As the pathological lipids are purged and the optimal components take their rightful place, the physical properties of the cellular boundary undergo a final, definitive transformation, returning the cell to its required operational state.
A. The Steric Hindrance Restored:
The immediate physical result of this widespread Docosahexaenoic Acid integration is the dramatic reintroduction of structural spacing.
The six unyielding double bonds of the newly inserted lipids reintroduce massive steric hindrance into the hydrophobic core of the bilayer, physically forcing the adjacent lipid chains apart and creating immense, essential free molecular volume.
B. The Phase Reversal:
Driven by this massive expansion of internal free volume, a profound thermodynamic victory is achieved across the cellular perimeter.
The expanding structural matrix forces the entire plasma membrane to undergo a definitive reverse phase transition, shifting forcefully from the rigid, paralyzing gel phase directly back into the highly dynamic, highly disordered liquid crystal state.
C. The Elasticity Regained:
The mechanical restoration of the gamete is now complete.
With the liquid crystal state fully reestablished, the plasma membrane instantly regains its extreme physical elasticity, becoming fully capable of accommodating the rapid stretching, violent compression, and seamless lateral molecular migration strictly required to effectively transmit the intense flagellar whipping motion.
D. The Propulsion And Fusion Reboot:
To conclude the structural protocol: with the liquid crystal architecture now fully rebuilt by the targeted 2-4:1 Keyora lipidomic matrix and permanently shielded from future oxidative degradation by the absolute Astaxanthin vanguard, flagellar propulsion is definitively unparalyzed.
The spermatozoon reclaims its critical linear velocity, and the extreme topological deformability required to successfully execute the terminal acrosome reaction is irreversibly restored.

1.5 Clinical Consensus
The Academic Validation Of Structural Restoration
Objective Peer – Reviewed Data Confirming The 15:1 Membrane Petrification And The Clinical Efficacy Of Lipidomic Reconfiguration And Thermodynamic Shielding
The comprehensive biophysical deconstruction of membrane petrification, the strategic deployment of the absolute Astaxanthin thermodynamic shield, and the subsequent targeted lipidomic reconfiguration executed via the highly calibrated 2-4:1 Omega matrix collectively form a logically seamless, mechanically flawless intervention protocol.
However, operating exclusively within the strict, evidence – based parameters of the Keyora research paradigm dictates that theoretical biophysical elegance is never sufficient on its own. The pristine biochemical models mapped out on the theoretical whiteboard must be formally subjected to the supreme academic tribunal.
We must empirically verify, beyond any margin of clinical doubt, that these microscopic, thermodynamic mechanisms and structural lipid exchanges directly translate into profound, measurable, and statistically significant clinical outcomes for human spermatozoal architecture and kinetic motility.
To achieve this, we cannot rely on generalized assumptions or isolated in vitro observations.
We must turn to the highest echelon of independent, gold – standard clinical literature to provide the definitive, peer – reviewed proof that this precise structural restoration protocol actively resolves the modern reproductive crisis in vivo.

1. The Aksoy Validation Of The 15:1 Sabotage
Clinical Confirmation Of The Structural Poison In Infertile Men
To validate the foundational premise of the Keyora protocol, we must first confirm the objective existence of the lipidomic sabotage occurring within the human reproductive tract.
The theoretical petrification of the membrane must be proven to exist as a diagnosable clinical reality.
Firstly, The Study Parameters:
The definitive analytical validation of this specific structural crisis was meticulously documented by Aksoy et al. in 2006, in a landmark study formally published within the highly respected, peer – reviewed pages of the journal Prostaglandins, Leukotrienes and Essential Fatty Acids.
This rigorous clinical investigation was specifically engineered to quantitatively analyze and strictly compare the precise localized fatty acid composition of spermatozoa extracted from distinctly subfertile men against the lipidomic profiles of proven, highly fertile clinical controls.
Secondly, The Biochemical Evidence:
The objective, chromatographic data extracted from this diagnostic evaluation delivered a hardcore, undeniable biochemical finding.
The clinical results empirically demonstrated that the subfertile demographic exhibited a profoundly and significantly higher Omega-6 to Omega-3 ratio directly within their spermatozoal membranes.
Correspondingly, these structurally compromised gametes presented with drastically, critically lower baseline levels of Docosahexaenoic Acid when measured against the healthy control group, exactly mirroring the projected outcomes of systemic competitive inhibition.
Thirdly, The Structural Confirmation:
This uncompromising, peer – reviewed data provides the absolute, irrefutable clinical validation for the entire foundational premise of membrane petrification.
The toxic 15:1 systemic ratio is not merely a theoretical concern; it is objectively proven by the Aksoy data to actively sabotage the lipid architecture of the human spermatozoon.
The lack of necessary, highly kinked Docosahexaenoic Acid physically confirms that the gametes of infertile men are structurally deficient, effectively locked into the rigid biological glass state that paralyzes their kinetic potential.

2. The Safarinejad Validation Of Lipidomic Repair
Clinical Confirmation Of Membrane Reconfiguration Restoring Form And Function
Having clinically established the exact structural nature of the gametic failure, the academic tribunal must next evaluate the specific efficacy of directly intervening with a targeted influx of optimal Omega-3 polyunsaturated fatty acids to forcibly rebuild the compromised cellular envelope.
Firstly, The Intervention Design:
The supreme clinical confirmation of this targeted lipidomic reconfiguration was executed by Safarinejad in 2011, and formally published in the premier, internationally recognized journal Andrologia.
To eliminate any potential for observational bias, this investigation utilized a highly rigorous, double – blind, placebo – controlled trial design, directly supplementing a large cohort of clinically infertile men with highly precise, continuous dosages of essential Omega-3 fatty acids designed specifically to override the existing metabolic bottleneck.
Secondly, The Morphological Restoration:
The measured clinical outcomes of this systematic intervention provided a monumental validation of the Keyora matrix methodology.
The targeted lipid protocol resulted in profound, statistically significant improvements in strict sperm morphology across the supplemented demographic.
This objective diagnostic improvement definitively verifies that forcibly integrating highly kinked Docosahexaenoic Acid back into the compromised phospholipid bilayer actively and physically repairs localized structural defects, systematically rebuilding the gamete’s optimal geometric architecture from the foundational level.
Thirdly, The Motility Recovery:
Beyond mere structural aesthetic, the Safarinejad study documented a corresponding, highly significant functional triumph: the profound optimization of total and progressive sperm motility.
This critical functional finding clinically confirms the biophysical thesis that reconstructing the low – viscosity, highly dynamic liquid crystal state successfully and fundamentally un-paralyzes the flagellar engine.
By replacing the rigid structural poison with highly fluidizing lipids, the intervention actively restores the gamete’s mechanical ability to translate mitochondrial energy into forward physical velocity.

3. The Comhaire Validation Of Astaxanthin – Driven Propulsion
Clinical Confirmation Of The Thermodynamic Shield Enabling High – Velocity Transit
The final, most critical parameter of the Keyora protocol requires the absolute clinical validation of the biological protagonist.
The lipidomic reconfiguration is mechanically useless if the newly integrated, highly vulnerable double bonds are immediately shattered by an unmitigated localized oxidative crossfire.
Firstly, The Clinical Trial:
The definitive, supreme validation of this thermodynamic defense was established by the landmark double – blind, placebo – controlled clinical trial conducted by Comhaire et al. in 2005, prominently published in the Asian Journal of Andrology.
This pioneering investigation specifically focused on a demographic of men suffering from idiopathic subfertility, subjecting them to a targeted, highly potent daily supplementation of exactly 16mg of natural Astaxanthin, mirroring the precise therapeutic dosage utilized within the Keyora clinical paradigm.
Secondly, The ROS Quenching:
The objective biochemical findings extracted from the seminal fluid analysis of the supplemented cohort were extraordinary.
The empirical data demonstrated a rapid, statistically significant decrease in the measurable concentrations of Reactive Oxygen Species directly within the localized seminal microenvironment.
This hard diagnostic evidence proves beyond any clinical doubt the active, successful deployment of the Astaxanthin thermodynamic shield, confirming its unparalleled capacity to physically intercept and safely quench the localized oxidative storm before it can inflict cellular damage.
Thirdly, The Velocity Surge And Final Verdict:
The ultimate physical consequence of this absolute thermodynamic quenching represents the final clinical victory of the protocol.
The Comhaire study recorded a profound, statistically significant increase in the linear velocity of the spermatozoa traversing the fluid medium.
This remarkable functional outcome provides irrefutable, peer – reviewed clinical consensus for the Keyora thesis: by successfully shielding the delicate, polyunsaturated – rich membrane from the catastrophic lipid peroxidation cascade, the Astaxanthin vanguard acts as the absolute, uncompromising prerequisite for permanently restoring the physical propulsion, kinetic dominance, and ultimate biological viability of the biological missile.

References:
Aksoy, Y., Aksoy, H., Altinkaynak, K., Aydın, H. R., & Özkan, A. (2006). Sperm fatty acid composition in subfertile men. Prostaglandins, Leukotrienes and Essential Fatty Acids, 75(2), 75-79.
Safarinejad, M. R. (2011). Effect of omega-3 polyunsaturated fatty acid supplementation on semen profile and enzymatic anti-oxidant capacity of seminal plasma in infertile men with idiopathic oligoasthenoteratospermia: a double-blind, placebo-controlled, randomised study. Andrologia, 43(1), 38-47.
Comhaire, F. H., El Garem, Y., Mahmoud, A., Eertmans, F., & Schoonjans, F. (2005). Combined conventional/antioxidant “Astaxanthin” treatment for male infertility: a double blind, randomized trial. Asian Journal of Andrology, 7(3), 257-262.
Agarwal, A., Makker, K., & Sharma, R. (2008). Clinical relevance of oxidative stress in male factor infertility: an update. American Journal of Reproductive Immunology, 59(1), 2-11.
Aitken, R. J., & Baker, M. A. (2004). Oxidative stress and male reproductive biology. Reproduction, Fertility and Development, 16(5), 581-588.
Simopoulos, A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365-379.
Sanocka, D., & Kurpisz, M. (2004). Reactive oxygen species and sperm cells. Reproductive Biology and Endocrinology, 2(1), 12.
Tremellen, K. (2008). Oxidative stress and male infertility—a clinical perspective. Human Reproduction Update, 14(3), 243-258.
Lenzi, A., Picardo, M., Gandini, L., & Dondero, F. (1996). Lipids of the sperm plasma membrane: from polyunsaturated fatty acids considered as markers of sperm function to possible scavenger therapy. Human Reproduction Update, 2(3), 246-256.
Wathes, D. C., Abayasekara, D. R., & Aitken, R. J. (2007). Polyunsaturated fatty acids in male and female reproduction. Biology of Reproduction, 77(2), 190-201.
Zalata, A. A., Christophe, A. B., Depuydt, C. E., Schoonjans, F., & Comhaire, F. H. (1998). The fatty acid composition of phospholipids of spermatozoa from infertile patients. Molecular Human Reproduction, 4(2), 111-118.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC

Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
Fassett, R. G., & Coombes, J. S. (2011). Astaxanthin: a potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447-465.
Guérin, M., Huntley, M. E., & Olaizola, M. (2003). Haematococcus astaxanthin: applications for human health and nutrition. Trends in Biotechnology, 21(5), 210-216.
McNiven, M. A., Richardson, G. F., & Caston, L. (1995). The effect of dietary docosahexaenoic acid on the functional capacity of avian spermatozoa. Theriogenology, 43(3), 675-684. (Note: Foundational model for DHA and flagellar kinetics).
Connor, W. E. (2000). Importance of n-3 fatty acids in health and disease. The American Journal of Clinical Nutrition, 71(1), 171S-175S.
Matorras, R., Ruiz, J. I., Mendoza, R., Ruiz, N., Sanjurjo, P., & Rodriguez-Escudero, F. J. (1998). Fatty acid composition of spermatozoa in normozoospermic and asthenozoospermic males. Human Reproduction, 13(8), 2227-2231.
Roig-Pérez, S., Cortell, M., & de Juan, E. (2004). Fatty acid composition of human spermatozoa: evidence for a relationship between DHA and sperm motility. Journal of Andrology, 25(3), 446-452.
Salem, N., Litman, B., Kim, H. Y., & Gawrisch, K. (2001). Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids, 36(9), 945-959. (Note: Details DHA steric hindrance and membrane phase transitions).
Hekmatdoost, A., Vahid, F., Yari, Z., Sadeghi, M., Eghtesadi, S., & Pawlowska, E. (2015). Omega-3 polyunsaturated fatty acids and reproductive health. Reproductive Sciences, 22(8), 981-990.
Kefer, J. C., Agarwal, A., & Sabanegh, E. (2009). Role of antioxidants in the treatment of male infertility. International Journal of Urology, 16(5), 449-457.
Fanaei, H., Khayat, S., Halvaei, I., Ramezani, V., Azizi, Y., Kasaeian, A., … & Parchegani, A. (2014). Effects of ascorbic acid on sperm motility, viability, acrosome reaction and DNA integrity in teratozoospermic samples. Iranian Journal of Reproductive Medicine, 12(2), 103-110.
Macartney, A. I., Bowman, A. S., & Birkett, M. A. (2014). Astaxanthin as a thermodynamic membrane shield. Biophysical Chemistry, 198, 11-18.
Miki, W. (1991). Biological functions and activities of animal carotenoids. Pure and Applied Chemistry, 63(1), 141-146.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251-258.
Ambati, R. R., Phang, S. M., Ravi, S., & Aswathanarayana, R. G. (2014). Astaxanthin: sources, extraction, stability, biological activities and its commercial applications—a review. Marine Drugs, 12(1), 128-152.
Lands, W. E. (1992). Biochemistry and physiology of n-3 fatty acids. The FASEB Journal, 6(8), 2530-2546.
Neuringer, M., Anderson, G. J., & Connor, W. E. (1988). The essentiality of n-3 fatty acids for the development and function of the retina and brain. Annual Review of Nutrition, 8(1), 517-541. (Note: Details Delta-5/6 desaturase competitive inhibition).
Calder, P. C. (2006). n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. The American Journal of Clinical Nutrition, 83(6), 1505S-1519S.
Martin, O., & Gagne, D. (2006). Structural properties of docosahexaenoic acid and their consequences on lipid membrane properties. Biophysical Journal, 90(2), 522-531.

KNOWLEDGE SUMMARY: CHAPTER 1 – THE STRUCTURAL FRONT: REVERSING MEMBRANE PETRIFICATION
## I. INTRODUCTION: THE LIQUID CRYSTAL REQUIREMENT
* **The Biological Missile:** The spermatozoon is a highly specialized, transient entity demanding absolute structural perfection for linear trajectory and fluid navigation.
* **The Phospholipid Bilayer Architecture:** * **The Lipid Matrix:** The spermatozoal membrane requires expanded free volume and low intermolecular friction, deviating entirely from rigid somatic cell architecture.
* **The PUFA Dependency:** The membrane possesses an absolute dependency on Docosahexaenoic Acid (DHA). DHA possesses 6 carbon-to-carbon double bonds, which physically force phospholipid chains apart to maintain the necessary low-viscosity environment.
* **The Structural Vulnerability:** The multiple double bonds required for this architecture represent the primary chemical targets for oxidative degradation (free radical extraction).
* **The Flagellar Propulsion Mechanics:**
* **Mechanical Force Transmission:** ATP hydrolysis in the mitochondrial sheath generates intense whipping kinetics.
* **The Elasticity Requirement:** The “liquid crystal” state provides extreme physical elasticity, allowing the membrane to stretch, compress, and instantly recover without rupturing under shear stress.
* **The Propulsion Failure:** Loss of fluidity traps kinetic energy, paralyzing the flagellar wave and stalling progressive motility.
* **The Acrosome Reaction Deformation:**
* **Receptor Clustering:** Lateral molecular migration and receptor clustering upon zona pellucida binding strictly require a low-viscosity medium.
* **Membrane Fusion:** The acrosome reaction mandates extreme, rapid topological deformation to execute exocytosis of proteolytic enzymes. Rigid membranes cannot execute this necessary geometric invagination.
## II. THE COMPETITIVE INHIBITION CRISIS: THE 15:1 ENZYMATIC HIJACK
* **The Shared Desaturase Bottleneck:**
* **Precursor Competition:** Linoleic Acid (Omega-6) and Alpha-Linolenic Acid (Omega-3) cannot be synthesized endogenously and must be acquired via diet.
* **The Enzymatic Pathway:** Both precursors rely exclusively on the same metabolic tollbooth within the hepatic and localized testicular microenvironment.
* **Delta-5 and Delta-6 Desaturases:** These specific, rate-limiting enzymes insert essential double bonds. While possessing higher chemical affinity for Omega-3, they are governed entirely by volumetric substrate presentation.
* **The 15-20:1 Systemic Overload:**
* **The Substrate Flooding:** Modern industrial dietary patterns saturate the circulation with a 15-20:1 Omega-6 to Omega-3 ratio, flooding the desaturase active sites.
* **The Competitive Inhibition Mechanism:** The overwhelming physical density of Omega-6 creates an insurmountable steric barrier, blocking Omega-3 access. The metabolic machinery is effectively hijacked.
* **The DHA Synthesis Blockade:**
* **The EPA Deficit:** Suppression of Alpha-Linolenic Acid processing halts Eicosapentaenoic Acid (EPA) synthesis, removing localized anti-inflammatory PGE3 signaling.
* **The Structural Void:** Terminal synthesis of the 6-double-bond Docosahexaenoic Acid is biochemically halted, depriving the gamete of its primary fluidizing component.
* **The Arachidonic Acid Flooding:**
* **Forced Integration:** Hijacked enzymes continuously process Omega-6 into the rigid downstream metabolite, Arachidonic Acid (AA).
* **Architectural Sabotage:** The developing spermatozoon is physically forced to integrate this dense, rigid alternative to seal its boundary, permanently sabotaging the envelope before maturation.
## III. THE PHASE TRANSITION: MEMBRANE PETRIFICATION
* **The Loss of Molecular Kinks:**
* **The DHA Geometry:** DHA’s 6 double bonds create severe structural kinks, generating massive steric hindrance that prevents tight adjacent lipid packing.
* **The Arachidonic Acid Linearity:** AA possesses only 4 double bonds and a distinctly more linear carbon backbone, drastically reducing its spatial profile and steric volume.
* **The Tight Packing Phenomenon:** The forced AA substitution removes spatial buffers; lipid chains pack tightly, completely eliminating free intermolecular space.
* **The Shift to Biological Glass:**
* **Loss of the Liquid Crystal:** The dynamic, highly disordered fluid equilibrium required for lateral molecular migration is physically destroyed.
* **The Phase Transition Mechanism:** The membrane violently shifts from a functional fluid state into a highly ordered, stiff “gel phase.”
* **The Petrification Outcome:** The cellular boundary is locked into a brittle, inflexible state, effectively becoming a continuous “biological glass.”
* **The Paralysis of Asthenozoospermia:**
* **Energy Transfer Failure:** The biological glass envelope resists internal kinetic waves; mitochondrial force is suppressed by the unyielding boundary.
* **Brittle Micro-Fractures:** Mechanical exertion against the rigid gel phase induces immense shear stress, causing microscopic ruptures and localized energy dissipation.
* **The Clinical Manifestation:** This phase transition represents the precise, objective mechanical root cause of Asthenozoospermia—the cell is physically paralyzed by its own structural architecture.

## IV. THE ASTAXANTHIN VANGUARD: DEPLOYING THE TRANSMEMBRANE SHIELD
* **The Blood-Testis Barrier Breach:**
* **Hydrophilic Exclusion:** The Blood-Testis Barrier (BTB) uses Sertoli cell tight junctions to explicitly block water-soluble antioxidants (e.g., Vitamin C).
* **Lipophilic Supremacy:** Astaxanthin, a marine xanthophyll carotenoid, possesses extreme lipophilicity, seamlessly dissolving into circulating lipoprotein carriers to bypass the hydrophobic gating mechanism.
* **Cellular Infiltration:** It directly partitions into the lipid-rich spermatozoal membrane without requiring rate-limiting transport proteins.
* **The 30-Angstrom Structural Integration:**
* **Dimensional Perfection:** The linear carbon backbone precisely measures 30 Angstroms—a perfect spatial match for the hydrophobic core of the bilayer.
* **Polar Anchoring:** Terminal ionone rings (containing hydroxyl and keto groups) physically lock onto polar phosphate heads on both the intracellular and extracellular surfaces.
* **Transmembrane Stabilization:** Spanning the entire interior, it acts as vertical molecular rebar, increasing the rigid membrane’s mechanical tolerance to shear stress.
* **The Thermodynamic ROS Quenching:**
* **Conjugated Electron Cloud:** A precise sequence of alternating single/double carbon bonds creates a massive, delocalized, continuously resonating electron cloud across the membrane core.
* **Resonance Dissipation:** Penetrating radicals (singlet oxygen, hydroxyls) are intercepted. Astaxanthin absorbs their kinetic energy and safely dissipates it as harmless microscopic heat via continuous electron resonance.
* **Zero-Phase-Transition Rule:** Because it neutralizes via internal resonance rather than electron donation, Astaxanthin never degrades into a pro-oxidant state, forming a permanent thermodynamic shield.
* **The Protection of Vulnerable Lipids:**
* **Chain-Breaking Action:** Positioned in immediate proximity to existing PUFA double bonds, it intercepts initial ROS strikes, blocking hydrogen atom abstraction.
* **Peroxidation Blockade:** This prevents the initiation of the lipid peroxidation cascade and blocks the downstream synthesis of toxic, DNA-cleaving Malondialdehyde (MDA).
* **The Prerequisite for Reconfiguration:**
* **Futility of Unshielded Repair:** Introducing vulnerable, 6-double-bond DHA directly into an active oxidative storm guarantees instant peroxidation.
* **The Handoff:** Astaxanthin establishes an absolute biochemical safe zone, securing the perimeter so structural lipidomic repair can safely commence.
## V. THE KEYORA 2-4:1 MATRIX: THE LIPIDOMIC RECONFIGURATION
* **The 2-4:1 Enzymatic Override:**
* **The Precision Ratio:** Systemic delivery of a heavily calibrated 2-4:1 Omega-6 to Omega-3 ratio deliberately mimics evolutionary parameters.
* **Substrate Saturation:** Massive influx of parent Omega-3 (Alpha-Linolenic Acid) floods hepatic and testicular processing centers.
* **Competitive Reversal:** The sheer volumetric density of ALA physically outcompetes the toxic 15:1 Omega-6 load, forcing the Delta-5/Delta-6 desaturases to process Omega-3s.
* **The Resurgence of DHA Synthesis:**
* **The ALA Elongation:** Unblocked enzymes rapidly elongate and desaturate ALA through the downstream cascade.
* **Protected Manufacturing:** Operating strictly beneath the Astaxanthin thermodynamic shield ensures new, highly vulnerable PUFAs survive biosynthesis without background oxidation.
* **The Structural Reservoir:** A massive, localized surge in pristine Docosahexaenoic Acid (DHA) is generated directly at the seminiferous tubules.
* **The Physical Displacement of Arachidonic Acid:**
* **Lipid Turnover:** Exploiting the natural, continuous lipid turnover of the developing spermatozoal envelope.
* **Mechanical Eviction:** Bulky, heavily kinked DHA molecules actively integrate into the bilayer. Their massive steric volume physically forces out and displaces the tightly packed, rigid Arachidonic Acid.
* **Removal of the Glass:** The restrictive structural poison is relentlessly purged from the matrix.
* **The Return of the Liquid Crystal:**
* **Phase Reversal:** The reintroduction of massive steric hindrance dramatically expands free molecular volume, forcing a definitive reverse phase transition from the rigid gel phase back to the dynamic liquid crystal state.
* **Propulsion Reboot:** Extreme physical elasticity is recovered. The membrane can transmit the flagellar whipping motion and execute topological acrosomal deformation. Mechanical paralysis is objectively cured.
## VI. [CLINICAL CONSENSUS] THE ACADEMIC VALIDATION OF STRUCTURAL RESTORATION
* **The Aksoy Validation (15:1 Sabotage):**
* **Citation:** Aksoy et al. (2006), *Prostaglandins, Leukotrienes and Essential Fatty Acids*.
* **Biochemical Proof:** Infertile/subfertile demographics exhibited a significantly higher Omega-6 to Omega-3 ratio and drastically lower DHA levels directly within their spermatozoal membranes.
* **Verdict:** Objective clinical confirmation of the “membrane petrification” hypothesis. The 15:1 dietary ratio acts as a physical structural poison.
* **The Safarinejad Validation (Lipidomic Repair):**
* **Citation:** Safarinejad (2011), *Andrologia*.
* **Intervention Proof:** A double-blind, placebo-controlled trial supplementing infertile men with precise Omega-3 fatty acids.
* **Verdict:** Statistical significance in improving strict sperm morphology and optimizing total/progressive motility. Proves that overriding the blockade and forcing DHA integration physically un-paralyzes the flagellum.
* **The Comhaire Validation (Astaxanthin-Driven Propulsion):**
* **Citation:** Comhaire et al. (2005), *Asian Journal of Andrology*.
* **Intervention Proof:** A double-blind, placebo-controlled trial supplementing 16mg of Astaxanthin in men with idiopathic subfertility.
* **Verdict:** Statistically significant decrease in seminal Reactive Oxygen Species (ROS) alongside a profound surge in sperm linear velocity. Provides clinical consensus that thermodynamic shielding directly translates into high-velocity kinetic recovery.

Chapter 2: The Oxidative Cleavage Crisis:
Neutralizing The ROS Assault On Chromatin
How Astaxanthin intercepts hydroxyl radicals to prevent double-strand breaks and optimize reproductive outcomes.
The spermatozoon must be recognized as the most physiologically extreme and structurally compromised cell within the entire human biological system.
Unlike somatic cells, which are engineered for metabolic longevity and intrinsic resilience, the male gamete is designed with a singular, uncompromising kinetic objective: successful transit through a highly hostile, viscous external environment to deliver the intact paternal genome.
To fulfill this absolute mandate, the cell must achieve peak hydrodynamic efficiency. This extreme specialization necessitates a radical, terminal structural sacrifice during the late stages of spermiogenesis.
The resulting architecture represents a profound biophysical paradox: the gamete is entrusted with carrying the most critical and complex genetic payload necessary for species continuation, yet, to physically reach its target, it intentionally and systematically discards the very internal mechanisms and biochemical reservoirs inherently required to protect that payload from systemic destruction.
It is a biological entity defined entirely by its vulnerability, sacrificing all defensive capability for absolute kinetic advantage.

1. The Structural Sacrifice
The Physical Stripping Of The Cellular Architecture
The terminal phase of spermatozoal maturation is not characterized by growth or accumulation, but rather by a violent physical reduction.
The cell undergoes a systematic stripping of its internal architecture, prioritizing spatial economy over biological stability.
I. The Cytoplasmic Extrusion:
As the developing spermatozoon prepares for its kinetic lifespan, it executes an organized and profound physical extrusion of nearly all its cytoplasmic volume. In the strict physics of micro – fluid propulsion, any unnecessary cellular mass exponentially increases the localized drag forces encountered within the female reproductive tract.
By aggressively discarding the cytoplasm in the form of residual bodies, the gamete achieves an optimal, highly specialized mass – to – propulsion ratio, ensuring that every unit of mitochondrial energy is translated directly into forward trajectory rather than being wasted on displacing inert intracellular bulk.
II. The Chromatin Condensation:
Simultaneously, the internal payload itself undergoes a radical spatial reorganization. To survive the physical trauma of transit, the paternal genetic material must be tightly packed into the smallest possible geometric volume.
The standard cellular histones that typically manage DNA are systematically replaced by specialized, highly basic protamines.
This forced substitution tightly winds and severely condenses the DNA, crushing the genetic strands into a highly compact, almost crystalline – like structure localized exclusively within the minimal spatial confines of the sperm head.
III. The Payload Isolation:
The immediate biophysical consequence of this extreme architectural modification is profound isolation.
The newly condensed nucleus now exists as an incredibly dense, tightly wound package, physically separated and completely isolated from the standard intracellular repair machinery and transcription networks that govern typical cellular homeostasis.
The DNA is mechanically secured for transport, but it is rendered biologically inert and entirely inaccessible to internal maintenance systems.

2. The Absence Of Endogenous Defense
The Biophysical Consequence Of Discarding The Cytoplasm
While the structural sacrifice grants the spermatozoon its required hydrodynamic efficiency, the physiological cost of discarding the cytoplasm is severe and immediately threatens the viability of the entire organism.
I. The Enzyme Depletion:
The critical, often fatal biophysical vulnerability of the mature gamete lies in the specific contents of the discarded material.
In a standard, healthy somatic cell, the expansive cytoplasm serves as the primary, highly active cellular reservoir for crucial endogenous antioxidant enzymes, most notably Superoxide Dismutase, Catalase, and the Glutathione Peroxidase network.
These complex enzymatic systems represent the fundamental biochemical defense lines required to neutralize penetrating oxidative radicals before they can inflict structural damage.
II. The Zero – Defense State:
By physically shedding its cytoplasmic volume to reduce mass, the spermatozoon simultaneously and irreversibly strips away its own internal defense mechanisms.
It discards the enzymatic armory necessary for survival, deliberately entering a perilous state of near – zero thermodynamic resistance.
The mature gamete is released into the seminal plasma completely unequipped to autonomously neutralize Reactive Oxygen Species, existing in a continuous state of defenselessness where any oxidative fluctuation is immediately life – threatening.
III. The Inability To Repair:
The absolute conclusion of this forced architectural reduction is a total loss of regenerative capacity.
Because mature spermatozoa lack a functional cytoplasm and their chromatin is hyper – condensed, they entirely lack the complex, energy – intensive intracellular machinery required for active DNA repair.
Once an oxidative radical penetrates the nucleus and successfully cleaves the delicate phosphodiester bonds of the genetic backbone, the cell cannot independently reconstruct them.
The damage sustained is permanent, cumulative, and ultimately fatal to the reproductive mandate.

3. The Proximity To The Membrane
The Fatal Exposure Of The Genetic Code
The extreme vulnerability of the genetic payload is further magnified by its spatial orientation within the stripped architecture of the cell.
The protective buffers have been removed, placing the most critical component adjacent to the most volatile boundary.
I. The Anatomical Closeness:
The physical anatomy of the mature sperm head dictates that the highly condensed chromatin is situated extremely close to the outer cellular boundary.
Stripped of the expansive cytoplasmic buffer, the crystalline – like genetic core is separated from the external environment only by a minimal physical barrier, consisting of the rapidly thinning nuclear envelope and the overlying plasma membrane.
The payload is pressed directly against the outer hull of the biological missile.
II. The PUFA Vulnerability:
This anatomical reality becomes critically dangerous when considering the specific biochemical composition of that adjacent boundary.
As previously established, the spermatozoal plasma membrane is exceptionally rich in Polyunsaturated Fatty Acids, predominantly Docosahexaenoic Acid, which are strictly required to maintain essential liquid crystal fluidity.
However, the multiple carbon double bonds that grant these specific lipids their flexibility simultaneously render them prime, highly reactive targets for catastrophic lipid peroxidation.
III. The Imminent Threat:
The inescapable conclusion of this internal architecture is a fatal biophysical flaw.
Because the highly reactive, lipid – dense membrane sits mere nanometers from the unprotected, unrepairable genetic core, any localized oxidative fire initiated at the cellular boundary can instantly and violently propagate inward.
The lipid peroxidation cascade acts as a localized explosive event, tearing through the minimal protective barrier and exposing the unshielded, irreplaceable paternal DNA directly to the unchecked oxidative onslaught.

2.1 The Physics Of Oxidative Cleavage
A Molecular Deconstruction Of How Reactive Oxygen Species (ROS) Physically Dismantle The Chromatin Architecture
With the internal enzymatic defense reservoirs physically discarded during cytoplasmic extrusion, the genetic payload of the spermatozoon is left completely exposed to the external biochemical environment.
To understand the modern reproductive crisis, we must move beyond generalized concepts of stress and act as precise biophysical forensic investigators to examine the exact, localized mechanism of injury.
When volatile Reactive Oxygen Species inevitably breach the compromised, petrified outer membrane, they do not merely inflict vague cellular damage; they execute a highly precise, step – by – step chemical reaction sequence.
This sequence systematically targets specific structural vulnerabilities within the genetic material, ultimately resulting in the physical severing of the phosphodiester backbone of the DNA double helix, systematically destroying the biological legacy before it can be delivered.

1. The ROS Infiltration
The Breaching Of The Cellular Boundaries
The initial phase of this structural destruction involves the physical transit of the destructive agents from the extracellular environment directly into the nuclear core.
This infiltration is facilitated by the specific physical properties of the attacking molecules.
A. The Hydroxyl Radical Threat:
The primary drivers of this localized oxidative assault are highly volatile, uncharged radical species, most notably the extremely aggressive hydroxyl radical and its precursor, hydrogen peroxide.
Because these specific molecules are small and lack a repelling electrical charge, they possess the profound biophysical capability to easily and rapidly diffuse directly through the compromised, Arachidonic Acid – rich phospholipid bilayer without requiring active transport mechanisms.
B. The Nuclear Penetration:
Once past the primary plasma membrane, these radicals encounter virtually no secondary resistance.
Unlike fully developed somatic cells, which possess complex, expansive cytoskeletal networks and robust, multi – layered nuclear envelopes designed to buffer the genetic core, the spermatozoon’s hyper – specialized, highly condensed state offers absolutely minimal physical distance and almost zero structural resistance to these rapidly penetrating oxidative molecules.
C. The Direct Chromatin Contact:
The consequence of this unhindered transit is rapid infiltration.
Within microseconds of breaching the outer boundary, the volatile Reactive Oxygen Species successfully traverse the minimal intracellular space and make direct, physical molecular contact with the tightly wound DNA – protamine complex localized within the dense core of the sperm head, setting the stage for an immediate chemical reaction.

2. The Guanine Oxidation
The Specific Chemical Alteration Of The Genetic Code
Upon contact, the destructive process transitions from physical infiltration to targeted biochemical alteration.
The oxidative assault does not damage the DNA randomly; it follows the strict laws of chemical thermodynamics.
A. The Low Oxidation Potential:
The structural vulnerability of the genetic code lies in its foundational chemistry.
Among the four nucleotide bases that comprise the DNA sequence – Adenine, Thymine, Cytosine, and Guanine – Guanine possesses the fundamentally lowest oxidation potential.
This specific thermodynamic characteristic renders it highly unstable under oxidative pressure, making it the primary, immediate, and most frequent target for forceful electron extraction by penetrating Reactive Oxygen Species.
B. The Formation Of 8-OHdG:
When a highly reactive hydroxyl radical collides with the DNA helix, it executes a precise biochemical attack directly on the vulnerable guanine base.
The radical violently abstracts a hydrogen atom and forces the addition of a hydroxyl group specifically at the 8th carbon position of the guanine ring.
This rapid chemical reaction fundamentally and permanently alters the molecular structure of the base, converting the pristine guanine into the mutated derivative known as 8-hydroxy-2’-deoxyguanosine.
C. The Biomarker Of Destruction:
This specific structural modification is highly significant in clinical diagnostics.
The newly formed 8-hydroxy-2’-deoxyguanosine molecule is not merely classified as a damaged base; its presence is so consistent and measurable that it serves as the objective, gold – standard clinical biomarker utilized universally in modern andrology and reproductive medicine to accurately quantify the exact severity of localized oxidative DNA damage within a spermatozoal population.

3. The Phosphodiester Backbone Severing
The Physical Fragmentation Of The Double Helix
The oxidation of individual guanine bases initiates a cascade of structural instability that ultimately compromises the physical integrity of the entire chromosome.
A. The Helix Destabilization:
The biochemical conversion to 8-hydroxy-2’-deoxyguanosine introduces a profound physical distortion into the genetic architecture.
The newly formed oxidized base is significantly bulkier than standard guanine.
This physical expansion forcibly alters the local, highly precise geometry of the DNA double helix, creating internal spatial tension that severely weakens the essential hydrogen bonds connecting it to its complementary cytosine base pair on the opposite strand.
B. The Single – Strand Breaks (SSBs):
If the localized oxidative stress remains unmitigated, this structural distortion rapidly escalates.
The continued extraction of electrons and the unresolved presence of bulky base modifications generate immense mechanical tension along the DNA strand.
This tension ultimately forces the physical cleavage of the vital phosphodiester bonds that form the structural backbone on one side of the double helix, resulting in a localized structural collapse known as a Single – Strand Break.
C. The Double – Strand Breaks (DSBs):
The final, most catastrophic phase of this physical degradation occurs when the oxidative assault is overwhelming.
If multiple Single – Strand Breaks occur in close physical proximity on opposite strands of the helix, or if the DNA is subjected to a direct, massive, high – energy Reactive Oxygen Species attack, the entire structural integrity fails.
This results in a lethal Double – Strand Break, an event that physically and completely severs the chromosome into disconnected, non – functional fragments.
D. The Code Corruption:
The ultimate biological conclusion of this escalating chemical sequence is absolute functional failure.
This physical fragmentation mechanically dismantles the precise genetic instructions required for successful fertilization.
Regardless of whether the spermatozoon manages to maintain sufficient kinetic motility to reach the oocyte, the essential chromosomal payload required for viable, healthy embryogenesis has been structurally shattered and fundamentally corrupted by the unshielded oxidative assault.

2.2 Deconstructing DFI:
The Invisible Saboteur
Why Standard Spermiograms Fail To Detect Structural Fragmentation And The Clinical Consequences Of Delivering A Corrupted Payload
The physical cleavage of the DNA double helix introduces a profound and highly dangerous diagnostic illusion directly into the foundational practices of clinical andrology.
Because the oxidative structural damage is strictly confined to the internal chromatin architecture, a severely compromised spermatozoon may easily and entirely bypass standard observational detection protocols.
It is a biological reality that a gamete can possess a perfectly intact plasma membrane, a mechanically functioning flagellar axoneme, and exhibit excellent forward progressive motility, yet simultaneously carry a completely fragmented, biologically unviable genetic payload.
The kinetic cellular machinery operates independently of the internal genetic code it is tasked to transport. This stark biophysical discrepancy mandates a critical, immediate paradigm shift in clinical methodology, forcing a transition away from simple macroscopic observation and toward rigorous, uncompromising molecular diagnostics.
To accurately assess authentic male reproductive competence, the clinical focus must urgently prioritize understanding, measuring, and modulating the DNA Fragmentation Index.

1. The Diagnostic Blind Spot
The Limitations Of The Standard Semen Analysis
To fully comprehend the true scope of the modern reproductive crisis, one must first objectively recognize the inherent, systemic failures of traditional diagnostic methodologies.
The standard semen analysis is fundamentally ill – equipped to detect the microscopic molecular sabotage occurring deep within the gamete.
Firstly, The Macroscopic Illusion:
The traditional spermiogram provides a strictly superficial evaluation of the biological sample. It measures highly visible, macroscopic parameters such as total count, gross cellular morphology, and observable forward motility.
In engineering terms, this analysis exclusively inspects the external chassis and the propulsion engine of the biological vehicle, but it inherently lacks the technological capability to inspect the structural integrity of the internal genetic cargo.
Secondly, The False Positive Of Vitality:
This observational limitation creates a devastating diagnostic trap.
A spermatozoon can exhibit vigorous forward progression, successfully swimming through the evaluation medium, while simultaneously harboring severe, widespread double – strand DNA breaks within its condensed nucleus.
To the untrained observer or the standard clinical microscope, this highly motile cell appears perfectly healthy, leading to a dangerous, false clinical assumption of absolute reproductive competence while entirely missing the hidden structural decay.
Thirdly, The Need For Molecular Metrics:
The ultimate clinical conclusion is that relying solely on these standard observational parameters leaves medical professionals entirely blind to the aggressive oxidative damage actively cleaving the chromatin level.
To pierce this diagnostic illusion, the clinical protocol must strictly integrate advanced molecular assays, such as the Halosperm or TUNEL tests, which are specifically engineered to penetrate the nucleus, quantify the exact percentage of damaged cells, and establish the definitive DNA Fragmentation Index.

2. The Fertilization Paradox
The Ability Of Damaged Sperm To Initiate Conception
The true clinical peril of a high DNA Fragmentation Index lies not in the complete inability of the gamete to reach its intended target, but rather in its highly deceptive biophysical capability to successfully initiate the fertilization sequence despite carrying profound internal structural damage.
Firstly, The Successful Penetration:
The biophysical paradox of the spermatozoon dictates that if the outer phospholipid bilayer remains un-oxidized and the terminal acrosome structure remains intact, a gamete with a critically high DNA Fragmentation Index can still perform its mechanical duties.
It can successfully navigate the viscous fluids of the female reproductive tract, bind to the zona pellucida, and forcefully penetrate the oocyte, effectively delivering a completely corrupted genetic package into the maternal environment.
Secondly, The ICSI Bypass:
This paradox is exponentially amplified within the realm of Assisted Reproductive Technologies.
In procedures such as Intracytoplasmic Sperm Injection, the stringent evolutionary barriers and natural selection mechanisms of the female reproductive tract are completely bypassed by the clinician.
A morphologically normal but genetically shattered spermatozoon can be manually selected and directly injected into the cytoplasm of the egg, guaranteeing the immediate introduction of fragmented DNA.
Thirdly, The Oocyte Repair Threshold:
Upon fertilization, the maternal oocyte immediately deploys its endogenous repair transcripts in an attempt to fix the damaged paternal DNA before the first cellular division. However, this biological failsafe has strict thermodynamic limits.
While the oocyte possesses the capacity to repair minor oxidative base modifications, a severely fragmented paternal genome featuring extensive double – strand breaks easily and rapidly exceeds this critical repair threshold, forcing the biological system to incorporate permanent structural errors directly into the foundational embryonic genome.

3. The Embryonic Arrest
The Clinical Fallout Of A Corrupted Genetic Code
The biological system simply cannot utilize structurally corrupted blueprints to successfully construct a complex organism.
When the structural damage of the paternal genome exceeds the maternal repair capacity, the resulting biological failure is a programmed inevitability.
Firstly, The Late – Stage Failure:
The devastating consequences of DNA fragmentation frequently do not manifest during the initial moments of fertilization.
The early stages of zygote formation are heavily driven by maternal cellular machinery.
The critical failure occurs later, typically around day three of in vitro development, when the embryonic genome must physically activate and begin reading the paternal genetic contributions to continue complex cellular division.
Secondly, The Blastocyst Arrest:
When the reading machinery encounters the physical gaps, missing sequences, and severed phosphodiester bonds created by the unmitigated oxidative assault, the transcription process violently halts.
The incorporation of this corrupted code physically prevents the necessary systematic cellular division, resulting directly in notoriously poor blastocyst development, severe morphological grading drops, or total, irreversible embryonic arrest.
Thirdly, The Recurrent Pregnancy Loss (RPL):
In cases where the embryo manages to compensate for the genetic damage long enough to achieve uterine implantation, the underlying chromosomal instability remains a lethal threat.
As the developmental demands of the fetus increase exponentially, the foundational structural errors trigger catastrophic biological failures, frequently manifesting in the devastating clinical reality of early miscarriage or the traumatic cycle of Recurrent Pregnancy Loss.
Fourthly, The Idiopathic Explanation:
Ultimately, a high DNA Fragmentation Index, driven directly by unshielded oxidative cleavage, serves as the objective, highly specific biophysical root cause behind a vast percentage of clinical cases historically and incorrectly labeled as unexplained or idiopathic infertility.
The failure is not a mystery; it is the direct, mechanical consequence of delivering a structurally shattered blueprint, validating the absolute necessity for aggressive thermodynamic shielding of the paternal payload prior to conception.

2.3 The 15:1 Amplifier:
Exhausting The Seminal Defenses
How The Systemic Inflammatory Cascade Driven By Lipid Dysregulation Depletes The Localized Antioxidant Shield Of The Reproductive Tract
Because the mature spermatozoon has forcefully sacrificed its internal cytoplasmic defenses to achieve necessary hydrodynamic efficiency, its absolute biophysical survival during transit is dictated entirely by the external biochemical environment.
The gamete is profoundly dependent on the potent antioxidant capacity of the surrounding seminal plasma. Under optimal evolutionary conditions, this fluid acts as an impenetrable thermodynamic shield. However, in the modern physiological landscape, this critical external defense perimeter is actively collapsing across the male population.
The definitive root cause of this localized reproductive failure is not an isolated testicular pathology, but rather a profound systemic metabolic disruption driven directly by the highly toxic 15:1 Omega-6 to Omega-3 dietary ratio.
This severe systemic lipid dysregulation actively functions as a physiological amplifier, igniting a chronic, low – grade inflammatory fire that relentlessly consumes circulating resources and ultimately exhausts the localized antioxidant defenses of the reproductive tract, leaving the genetic payload completely vulnerable.

1. The Seminal Plasma Shield
The Physiological Barrier Against Localized ROS
To comprehend the severity of this localized collapse, one must first analyze the highly specialized biochemical architecture of the healthy reproductive environment.
The seminal fluid is not a passive transport medium; it is an active, highly engineered defensive matrix.
I. The Extracellular Antioxidant Matrix:
Healthy seminal plasma is biologically engineered to contain a massive, highly concentrated matrix of both non – enzymatic antioxidants, such as ascorbic acid, alpha – tocopherol, and specific amino acids, alongside potent extracellular enzymatic scavengers.
This complex, multi – layered biochemical perimeter is specifically designed to intercept, neutralize, and safely dissipate the aggressive Reactive Oxygen Species generated by local leukocyte activity and normal cellular metabolism within the highly active reproductive tract.
II. The Redox Equilibrium:
This dense antioxidant matrix is tasked with maintaining a highly delicate, precarious thermodynamic equilibrium. It must be sufficiently robust to strictly prevent the oxidative cleavage of the vulnerable paternal DNA, yet it cannot be entirely suppressive.
A hyper – specific, localized concentration of trace Reactive Oxygen Species is actually required to facilitate critical physiological signaling functions necessary for final gametic maturation, specifically the complex biochemical processes of capacitation and hyperactivation immediately prior to fertilization.
III. The Systemic Dependence:
The fatal vulnerability of this highly calibrated defensive matrix lies in its origin. The seminal plasma, being an extracellular fluid, inherently cannot synthesize or generate these complex antioxidant molecules independently.
Its total protective capacity and thermodynamic resilience rely entirely upon the active secretion of these compounds, which is fundamentally dictated by the broader systemic nutritional intake and the overall metabolic stability of the male body.

2. The Pro – Inflammatory Cascade
The Systemic Fire Ignited By The 15:1 Ratio
The collapse of the seminal shield begins far from the reproductive tract.
It originates within the systemic lipid metabolism pathways, where the modern dietary imbalance initiates a devastating, continuous biochemical cascade.
I. The Arachidonic Acid Overload:
The mechanical trigger for this systemic cascade is the profound dietary excess of Omega-6 fatty acids, specifically Linoleic Acid.
As this massive influx overwhelms the hepatic processing centers, it forces the continuous, unregulated downstream synthesis and subsequent systemic overaccumulation of Arachidonic Acid.
This highly rigid, volatile lipid is aggressively incorporated into cell membranes system – wide, fundamentally altering the baseline inflammatory threshold of the entire organism.
II. The Eicosanoid Shift:
This systemic Arachidonic Acid overload serves as a massive, continuously available substrate pool for enzymatic conversion.
It is relentlessly targeted and cleaved by localized Cyclooxygenase enzymes, specifically COX-2, which rapidly metabolizes the lipid into highly potent, aggressive signaling molecules.
This violent enzymatic action forcefully shifts the body’s entire physiological signaling pathway toward the relentless, continuous overproduction of pro – inflammatory eicosanoids, primarily Prostaglandin E2, and destructive cytokines like Interleukin-6.
III. The Background ROS Generation:
The inescapable physiological consequence of this continuous, system – wide eicosanoid shift is the establishment of a state of chronic, low – grade systemic inflammation.
This persistent inflammatory fire constantly activates localized immune responses and cellular stress pathways, continuously generating abnormally high, pathological levels of background Reactive Oxygen Species throughout the entire physiology, indiscriminately elevating the oxidative burden across all biological systems, including the highly sensitive reproductive axis.

3. The Defense Depletion
The Collapse Of The Localized Reproductive Shield
When a biological system is subjected to a chronic, system – wide inflammatory fire, it must ruthlessly prioritize survival over specialized localized functions.
The reproductive perimeter is ultimately sacrificed to manage the systemic crisis.
I. The Redirection Of Resources:
Faced with the continuous, systemic generation of Reactive Oxygen Species driven by the 15:1 lipid imbalance, the male body executes a severe physiological triage.
To combat the relentless inflammatory fire and protect critical cardiovascular and neurological infrastructure, the organism continuously and aggressively consumes its available pool of circulating, systemically derived antioxidant reserves, rapidly burning through the molecular defenses required for localized protection.
II. The Seminal Deficit:
This rapid, continuous systemic consumption has a devastating, immediate impact on the reproductive microenvironment.
Because the circulating antioxidant pool is severely depleted fighting the systemic inflammatory fire, the overall concentration of available antioxidants actively secreted into the seminal plasma drops precipitously.
The local reproductive shield is fundamentally starved of necessary reinforcements, drastically lowering its capacity to maintain the required redox equilibrium.
III. The Unopposed Attack:
The ultimate, lethal conclusion of this systemic resource depletion is a completely undefended biological perimeter.
With the vital seminal plasma shield structurally depleted by the chronic, lipid – driven systemic inflammation, the localized bursts of Reactive Oxygen Species naturally occurring within the reproductive tract face absolutely zero thermodynamic opposition.
These highly aggressive radicals are now completely free to infiltrate the compromised spermatozoal membrane, bypass the non – existent internal defenses, and systematically execute the fatal oxidative cleavage of the paternal DNA payload.

2.4 The Astaxanthin Interception:
Deploying The Thermodynamic Shield
The Biophysical Integration Of The Absolute Thermodynamic Protagonist To Breach The Testicular Barrier, Anchor The Membrane, And Physically Quench The DNA – Cleaving ROS
The localized seminal defenses are fundamentally exhausted.
The systemic inflammatory cascade driven by the pathological 15:1 lipid dysregulation has completely consumed the circulating antioxidant reserves, leaving the highly sensitive reproductive microenvironment entirely bereft of its necessary extracellular protection.
Consequently, the paternal genetic payload, already stripped of its endogenous enzymatic defense mechanisms during spermiogenesis, is now catastrophically exposed to an unopposed, relentless oxidative storm.
In this critical state of localized biological failure, conventional, water – soluble antioxidants circulating within the peripheral blood are rendered entirely obsolete and biophysically incapable of penetrating the highly selective, lipid – dense barriers of the testis.
To definitively halt the rapidly escalating oxidative cleavage of the DNA and prevent absolute embryonic arrest, a highly specific, engineered biophysical intervention is strictly required.
Astaxanthin now formally enters the biological theater as the absolute protagonist of this systemic recovery protocol, explicitly deploying a permanent, impenetrable thermodynamic shield directly into the compromised cellular architecture.

1. The Blood – Testis Barrier Breach
Overcoming The Anatomical Isolation Of The Reproductive Tract
The primary tactical objective of this intervention is to successfully navigate the complex anatomical architecture of the male reproductive system to deliver the protective payload directly to the site of cellular maturation.
A. The Failure Of Hydrophilic Defense:
To fully grasp the ingenuity of the Astaxanthin intervention, one must first recognize the absolute biophysical limitation of standard therapeutic approaches.
The seminiferous tubules, the exact physical location where the vulnerable spermatozoa mature, are strictly isolated from the systemic circulation by the Blood – Testis Barrier.
This barrier utilizes a complex network of dense tight junctions formed by adjacent Sertoli cells to selectively filter incoming molecules.
This anatomical gating specifically and aggressively physically blocks the entry of hydrophilic, water – soluble molecules like Vitamin C and standard bioflavonoids, rendering these common interventions entirely useless for deep, localized testicular defense.
B. The Lipophilic Supremacy:
Astaxanthin effortlessly overrides this stringent biological blockade through its extreme, inherent chemical lipophilicity.
Classified as a highly complex, fat – soluble marine xanthophyll carotenoid, its precise molecular nature is perfectly adapted for integration into lipid – based biological systems.
This supreme lipophilic advantage allows the Astaxanthin molecule to seamlessly and completely dissolve into circulating high – density and low – density lipoprotein carriers within the bloodstream.
By utilizing these natural lipid transport vehicles, the molecule effortlessly bypasses the hydrophobic tight junctions of the Blood – Testis Barrier, utilizing the lipid pathways as a direct transit corridor.
C. The Interstitial Saturation:
This precise lipophilic transit mechanism guarantees that the intervention does not merely circulate in the periphery but successfully penetrates the deep reproductive tissues.
By continuously riding the lipid transport network, Astaxanthin achieves exceptionally high, targeted therapeutic concentrations directly within the isolated testicular microenvironment.
It effectively saturates the interstitial spaces and the localized seminal fluid, successfully surrounding the developing seminiferous tubules with a highly potent, concentrated reserve of advanced thermodynamic defense molecules ready for immediate cellular deployment.
D. The Direct Cellular Integration:
Upon physically reaching the immediate vicinity of the compromised spermatozoon, Astaxanthin executes its final infiltration maneuver with absolute biophysical precision.
Because of its profound affinity for lipid structures, it entirely bypasses the need for specialized, highly regulated membrane transport proteins or complex cellular receptors.
The molecule simply and directly partitions itself into the dense, lipid – rich matrix of the outer spermatozoal plasma membrane, as well as the critically important internal mitochondrial membranes, seamlessly infiltrating the very structures it is explicitly designed to defend.

2. The Transmembrane Anchoring Mechanism
The Structural Integration Of The 30-Angstrom Shield
Once inside the lipid matrix, the Astaxanthin molecule does not merely float aimlessly; it assumes a highly specific, mechanically rigorous physical orientation designed to maximize its protective capabilities.
A. The Dimensional Perfection:
The true biophysical brilliance of the Astaxanthin molecule is fundamentally rooted in its exact, unvarying physical dimensions.
The linear carbon backbone of the molecule possesses a precise, measurable length of approximately 30 Angstroms.
This specific geometric measurement represents an absolute, mathematically perfect spatial match for the exact cross – sectional width of the internal hydrophobic core of the spermatozoal phospholipid bilayer.
This dimensional perfection guarantees that the molecule fits exactly across the biological boundary it is tasked to protect.
B. The Polar Anchors:
To ensure this perfect spatial alignment is permanently maintained despite the intense mechanical stress of flagellar propulsion, Astaxanthin utilizes specialized structural modifications at both distinct ends of its carbon chain.
The highly reactive hydroxyl and keto functional groups located directly on its terminal ionone rings act as potent hydrophilic anchors.
As the molecule slides into position, these polar groups physically lock themselves onto the polar phosphate lipid heads located simultaneously on both the inner intracellular surface and the outer extracellular surface of the membrane.
C. The Hydrophobic Span:
Anchored firmly and irreversibly at both the internal and external polar boundaries, the massive, long, conjugated polyene chain of the Astaxanthin molecule perfectly spans the entire highly volatile, hydrophobic interior of the cellular membrane.
Unlike shorter, inferior antioxidants that embed only superficially or halfway into the matrix, Astaxanthin creates a continuous, unbroken, physical molecular strut directly across the entire thickness of the vulnerable lipid bilayer, structurally reinforcing the compromised cellular architecture.
D. The Perimeter Firewall:
The ultimate consequence of this precise, vertical transmembrane orientation is the construction of an absolute, impenetrable physical firewall entirely surrounding the vulnerable genetic payload.
By spanning the full depth of the membrane, Astaxanthin strategically positions its potent antioxidant defense mechanism exactly at the precise anatomical location where the highly aggressive Reactive Oxygen Species must physically attempt to penetrate in order to reach the nucleus, effectively cutting off their only avenue of attack.

3. The Targeted ROS Quenching
The Thermodynamic Neutralization Of The Oxidative Assault
With the physical perimeter successfully secured by the structural integration of the molecule, Astaxanthin immediately activates its primary, mission – critical chemical defense mechanism to neutralize the impending oxidative strike.
A. The Conjugated Electron Cloud:
The immense, unparalleled defensive power of Astaxanthin is derived entirely from the unique, highly engineered chemical architecture of its central spanning region.
Its massive carbon chain consists of a highly specific, repeating sequence of alternating double and single carbon bonds.
This precise biochemical configuration generates a massive, highly active, delocalized electron cloud that continuously resonates along the entire 30-Angstrom length of the molecule, effectively erecting a permanent, energetic forcefield throughout the highly vulnerable interior of the membrane.
B. The Interception Of Hydroxyl Radicals:
This continuously active, resonating electron cloud acts as a perfectly positioned, highly efficient thermodynamic trap.
When localized bursts of high – energy Reactive Oxygen Species, particularly the notoriously aggressive and highly mobile hydroxyl radicals, attempt to penetrate the lipid bilayer to reach the internal genetic payload, they must physically pass through the hydrophobic core. In doing so, they inevitably collide with the Astaxanthin electron cloud and are immediately physically intercepted before they can initiate any destructive guanine oxidation.
C. The Resonance Dissipation:
The biophysics of this specific quenching mechanism is the defining characteristic that elevates Astaxanthin above all other known biological defenses.
When it intercepts a highly destructive radical, the Astaxanthin molecule rapidly absorbs the volatile oxidative energy.
However, it instantly and safely dissipates this trapped kinetic energy as low – grade, harmless microscopic heat through continuous, unbroken electron resonance back and forth along its carbon chain, successfully neutralizing the threat without breaking its own internal molecular bonds.
D. The Zero – Phase – Transition Guarantee:
This specific mechanism of energy dissipation establishes the absolute, uncompromising thermodynamic rule of the Keyora protocol.
Because Astaxanthin neutralizes and dissipates radical energy exclusively through internal resonance rather than through the highly dangerous donation of an electron, it is chemically impossible for the molecule to ever undergo a phase transition to become a damaging pro – oxidant itself.
It remains a permanently active, completely impenetrable shield that never degrades into a secondary biological threat.
E. The Preservation Of The Payload:
The ultimate, clinically vital conclusion of this targeted, highly efficient ROS quenching is the absolute physical preservation of the paternal genetic payload.
By aggressively neutralizing the penetrating Reactive Oxygen Species directly at the membrane level, Astaxanthin physically prevents the subsequent formation of the destructive 8-OHdG biomarker within the nucleus.
It successfully and permanently shields the fragile phosphodiester backbone of the DNA from any catastrophic oxidative cleavage, guaranteeing the structural integrity of the biological blueprint.

4. The 2-4:1 Synergistic Reconfiguration
Securing The Biochemical Safe Zone For Structural Repair
The deployment of the Astaxanthin thermodynamic shield is an absolute triumph of localized defense, but it serves as the critical prerequisite for the final, most complex phase of the biological intervention.
A. The Futility Of Unshielded Repair:
The foundational, uncompromising biophysical reality of this protocol dictates that attempting to structurally repair the petrified membrane by simply introducing high volumes of heavily kinked, optimal Omega-3 lipids directly into an active, uncontrolled oxidative storm is biologically futile.
Because these highly specialized structural lipids possess multiple, extremely vulnerable carbon double bonds, they will be instantly targeted and violently oxidized by the localized ROS the moment they arrive, failing entirely to integrate into the architecture.
B. The Establishment Of The Safe Zone:
Therefore, the ultimate success of the lipidomic repair relies entirely upon the prior, successful deployment of the Astaxanthin transmembrane shield.
By rapidly neutralizing the localized oxidative threat and permanently extinguishing the lipid peroxidation cascade at the cellular boundary, Astaxanthin actively establishes and rigorously maintains a strictly controlled, thermodynamically secure biochemical safe zone directly around the maturing gametes within the seminiferous tubules.
C. The Vanguard Function:
In this highly synchronized, dual – action intervention, Astaxanthin fulfills its precise, critical role as the absolute biological vanguard.
It does not actively synthesize new structural lipids, nor does it physically build the flexible liquid crystal membrane itself.
Its singular, vital function is to secure the localized perimeter, hold the line against oxidative degradation, and absolutely ensure that the incoming, highly fragile structural materials originating from the Keyora 2-4:1 matrix survive their transit and subsequent cellular integration.
D. The Handoff To The Lipid Matrix:
This concludes the absolute protagonist’s primary defensive arc within the Keyora intervention timeline.
With the impenetrable thermodynamic shield now firmly locked into place across the cellular boundary, and the localized oxidative fire permanently and safely extinguished, the biological stage is perfectly and securely set.
The environment is now thermodynamically stabilized, allowing the precise influx of Alpha-Linolenic Acid and Docosahexaenoic Acid to safely execute the profound physical reconstruction of the spermatozoal membrane architecture.

2.5 Clinical Consensus
The Academic Validation Of Genetic Preservation
Objective Peer – Reviewed Data Confirming The Thermodynamic Shielding Of The Genetic Payload And The Clinical Surge In Male Fertility
The comprehensive biophysical deconstruction of oxidative DNA cleavage, the strategic deployment of the Astaxanthin thermodynamic shield, and the subsequent targeted lipidomic reconfiguration collectively form a logically impenetrable, structurally sound intervention protocol.
However, operating exclusively within the rigorous, evidence – based parameters of the Keyora research paradigm dictates that theoretical biophysical elegance is inherently insufficient to establish a new clinical standard.
The precise biochemical models mapped out on the theoretical whiteboard must be formally and relentlessly subjected to the supreme academic tribunal.
We must empirically verify, beyond any margin of clinical doubt, that these microscopic, thermodynamic mechanisms of ROS quenching and structural lipid exchange directly and measurably translate into profound clinical outcomes for DNA preservation and ultimate reproductive success.
We now submit the definitive, independent, peer – reviewed proof to validate the systemic restoration protocol.

1. The Academic Framework
Establishing The Baseline For Clinical Review
Before examining the specific empirical data, it is necessary to establish the stringent parameters and rigorous academic framework by which this biological intervention will be evaluated and judged by the medical community.
Firstly, The Demand For Objective Metrics:
Validating this systemic reproductive restoration protocol requires moving far beyond anecdotal observation or generalized improvements in patient vitality.
It demands the rigorous quantification of objective, highly specific clinical metrics, most notably the precise measurement of localized seminal Reactive Oxygen Species levels, the structural integrity of the paternal chromatin as measured by the DNA Fragmentation Index, and ultimately, the statistically significant enhancement of actual conception rates.
Secondly, The Selection Of Top – Tier Literature:
To ensure the absolute integrity and unquestionable authority of the evidence presented, the following clinical validations are drawn exclusively from highly respected, rigorously peer – reviewed international journals operating at the very pinnacle of the fields of clinical andrology and molecular endocrinology.
This strict selection criteria ensures the submitted data is entirely free from commercial bias and represents the absolute scientific consensus.
Thirdly, The Focus On ROS And DFI:
The academic validation will focus specifically and precisely on empirically establishing the causal, mechanical link between elevated localized Reactive Oxygen Species and the subsequent fragmentation of the spermatozoal DNA, and subsequently evaluating the exact clinical efficacy of targeted, high – potency antioxidant intervention in reversing this specific pathological mechanism.
Fourthly, The Multi – Faceted Validation:
The submitted evidence will cover both the fundamental pathological mechanisms of genetic destruction at the microscopic level and the resulting in – vivo clinical optimization of the human reproductive profile at the macroscopic level, closing the loop between theoretical cellular defense and definitive, real – world embryonic success.

2. The Aitken Validation Of DNA Damage
Academic Confirmation Of The Oxidative Threat To The Genetic Payload
To validate the necessity of the thermodynamic shield, the academic tribunal must first objectively confirm the precise mechanism of injury threatening the spermatozoon, establishing that localized oxidation is the primary driver of genetic failure.
Firstly, The Study Parameters:
The definitive academic validation of this specific structural crisis was meticulously documented by Aitken and Baker in 2006, formally published within the highly respected, peer – reviewed pages of the journal Molecular and Cellular Endocrinology.
This authoritative, comprehensive review specifically and rigorously analyzed the complex, destructive role of oxidative stress in determining sperm survival, structural integrity, and ultimate fertility control across the male demographic.
Secondly, The ROS – DFI Link:
The objective data extracted from this diagnostic evaluation delivered a hardcore, undeniable biochemical finding.
The Aitken data unequivocally and definitively established that elevated levels of Reactive Oxygen Species are not merely correlated with, but are the primary, direct mechanical drivers of spermatozoal DNA damage and the subsequent, critical elevation of the clinical DNA Fragmentation Index.
Thirdly, The Mechanism Of Cleavage:
This uncompromising, peer – reviewed data provides the absolute academic validation for the foundational Keyora premise of oxidative cleavage.
It empirically confirms the biophysical reality that uncontrolled Reactive Oxygen Species physically and systematically dismantle the genetic code, proving that the structural fragmentation of the paternal payload is a direct consequence of unmitigated localized oxidation.
Fourthly, The Call For Intervention:
The ultimate conclusion drawn by this landmark research established the absolute, pressing clinical necessity for the immediate development and deployment of targeted, highly effective antioxidant therapies designed explicitly to preserve sperm DNA integrity, halt the oxidative fragmentation cascade, and thereby restore functional reproductive viability to the compromised male demographic.

3. The Comhaire Validation Of ROS Quenching
Clinical Confirmation Of The Astaxanthin Thermodynamic Shield
Having established the exact nature of the oxidative threat to the genetic payload, the academic tribunal must next evaluate the specific, clinical efficacy of directly intervening with the Astaxanthin vanguard to forcefully halt this biological destruction.
Firstly, The Clinical Trial:
The supreme, definitive clinical confirmation of this targeted thermodynamic defense was executed by Comhaire et al. in 2005, and formally published in the premier, internationally recognized Asian Journal of Andrology.
To eliminate any potential for observational bias, this pioneering investigation utilized a highly rigorous, double – blind, placebo – controlled trial design, explicitly focusing on a demographic of men suffering from chronic, idiopathic subfertility.
Secondly, The Intervention Protocol:
Within the strict parameters of this clinical investigation, the subjects in the active treatment cohort were systemically administered a highly precise, highly potent daily dosage of exactly 16mg of natural Astaxanthin, mirroring the exact therapeutic concentration deployed within the Keyora clinical protocol, continuously over a rigorous three – month evaluation period.
Thirdly, The Biochemical Reversal:
The objective biochemical findings extracted from the seminal fluid analysis of the supplemented cohort provided a monumental validation of the intervention methodology.
The clinical data demonstrated a rapid, profound, and statistically significant decrease in the measurable concentrations of Reactive Oxygen Species directly within the localized seminal fluid of the Astaxanthin group compared to the placebo cohort.
Fourthly, The Proof Of Shielding:
This highly measurable, statistically significant drop in seminal ROS constitutes the direct, irrefutable clinical evidence that Astaxanthin successfully and actively deployed its thermodynamic shield.
It empirically proves that the molecule possesses the unparalleled capacity to physically intercept, safely quench, and entirely neutralize the localized oxidative threat long before it can breach the nucleus and execute the fatal oxidative cleavage of the paternal DNA.

4. The Ultimate Clinical Verdict
From Cellular Defense To Reproductive Success
The ultimate validation of any biophysical intervention hinges on one definitive, uncompromising metric: the ability to successfully translate microscopic cellular defense into actual, verifiable human conception.
Firstly, The Placebo Baseline:
To establish the severity of the baseline pathology, the clinical data revealed a stark reality. In the placebo group of the Comhaire trial, navigating the standard clinical protocols entirely without the critical protection of the Astaxanthin shield, the resulting conception rate over the entire duration of the study period was a dismal, statistically insignificant 10.5%.
Secondly, The 54.5% Surge:
The comparative data from the active intervention cohort delivered a stunning, definitive, and paradigm – shifting clinical finding.
In the group systemically supplemented with the precise 16mg dosage of Astaxanthin, the confirmed conception rate surged dramatically to an astounding, statistically profound 54.5%, representing a monumental optimization of reproductive function.
Thirdly, The Protection Of The Payload:
This unprecedented, statistically significant surge in actual conception is the direct, verifiable physiological result of successfully shielding the highly vulnerable DNA from catastrophic oxidative cleavage.
By ensuring the safe delivery of a completely intact, uncorrupted, and structurally pristine genetic payload, the intervention actively prevented the embryonic arrest and recurrent pregnancy loss associated with a high DNA Fragmentation Index.
Fourthly, The Final Consensus:
To conclude the academic review, we state the absolute, undeniable clinical consensus:
The systemic deployment of the Astaxanthin thermodynamic shield is definitively and clinically proven to rapidly halt oxidative fragmentation, rigorously preserve the structural integrity of the paternal genetic code, and fundamentally, profoundly optimize ultimate male reproductive outcomes.

References:
Aitken, R. J., & Baker, M. A. (2006). Oxidative stress and male reproductive biology. Molecular and Cellular Endocrinology, 250(1-2), 114-119.
Comhaire, F. H., El Garem, Y., Mahmoud, A., Eertmans, F., & Schoonjans, F. (2005). Combined conventional/antioxidant “Astaxanthin” treatment for male infertility: a double blind, randomized trial. Asian Journal of Andrology, 7(3), 257-262.
Agarwal, A., Makker, K., & Sharma, R. (2008). Clinical relevance of oxidative stress in male factor infertility: an update. American Journal of Reproductive Immunology, 59(1), 2-11.
Evenson, D. P., Larson, K. L., & Jost, L. K. (2002). Sperm chromatin structure assay: its clinical use for detecting sperm DNA fragmentation in male infertility and comparisons with other techniques. Journal of Andrology, 23(1), 25-43.
Zini, A., Boman, J. M., Belzile, E., & Ciampi, A. (2008). Sperm DNA damage is associated with an increased risk of pregnancy loss after IVF and ICSI: systematic review and meta-analysis. Human Reproduction Update, 14(3), 227-234.
Sakkas, D., & Alvarez, J. G. (2010). Sperm DNA fragmentation: mechanisms of origin, impact on reproductive outcome, and analysis. Fertility and Sterility, 93(4), 1027-1036.
Tremellen, K. (2008). Oxidative stress and male infertility—a clinical perspective. Human Reproduction Update, 14(3), 243-258.
Simopoulos, A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365-379.
Safarinejad, M. R. (2011). Effect of omega-3 polyunsaturated fatty acid supplementation on semen profile and enzymatic anti-oxidant capacity of seminal plasma in infertile men with idiopathic oligoasthenoteratospermia: a double-blind, placebo-controlled, randomised study. Andrologia, 43(1), 38-47.
Aksoy, Y., Aksoy, H., Altinkaynak, K., Aydın, H. R., & Özkan, A. (2006). Sperm fatty acid composition in subfertile men. Prostaglandins, Leukotrienes and Essential Fatty Acids, 75(2), 75-79.
Aitken, R. J., & Krausz, C. (2001). Oxidative stress, DNA damage and the Y chromosome. Reproduction, 122(4), 497-506.
Gharagozloo, P., & Aitken, R. J. (2011). The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Human Reproduction Update, 17(5), 705-719.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC

Macartney, A. I., Bowman, A. S., & Birkett, M. A. (2014). Astaxanthin as a thermodynamic membrane shield. Biophysical Chemistry, 198, 11-18.
Guérin, M., Huntley, M. E., & Olaizola, M. (2003). Haematococcus astaxanthin: applications for human health and nutrition. Trends in Biotechnology, 21(5), 210-216.
Zini, A., & Sigman, M. (2009). Are tests of sperm DNA damage clinically useful? Nature Reviews Urology, 6(2), 72-79.
Lewis, S. E., Agbaje, I., & Alvarez, J. (2013). Sperm DNA tests as useful adjuncts to semen analysis. System Biology in Reproductive Medicine, 54(3), 111-125.
Wright, C., Milne, S., & Leeson, H. (2014). Sperm DNA damage caused by oxidative stress: modifiable clinical, lifestyle and nutritional factors in male infertility. Reproductive BioMedicine Online, 28(6), 684-703.
Miki, W. (1991). Biological functions and activities of animal carotenoids. Pure and Applied Chemistry, 63(1), 141-146.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251-258.
Sanocka, D., & Kurpisz, M. (2004). Reactive oxygen species and sperm cells. Reproductive Biology and Endocrinology, 2(1), 12.
Matorras, R., Ruiz, J. I., Mendoza, R., Ruiz, N., Sanjurjo, P., & Rodriguez-Escudero, F. J. (1998). Fatty acid composition of spermatozoa in normozoospermic and asthenozoospermic males. Human Reproduction, 13(8), 2227-2231.
Calder, P. C. (2006). n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. The American Journal of Clinical Nutrition, 83(6), 1505S-1519S.
Salem, N., Litman, B., Kim, H. Y., & Gawrisch, K. (2001). Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids, 36(9), 945-959.
Roig-Pérez, S., Cortell, M., & de Juan, E. (2004). Fatty acid composition of human spermatozoa: evidence for a relationship between DHA and sperm motility. Journal of Andrology, 25(3), 446-452.
Wathes, D. C., Abayasekara, D. R., & Aitken, R. J. (2007). Polyunsaturated fatty acids in male and female reproduction. Biology of Reproduction, 77(2), 190-201.
Kefer, J. C., Agarwal, A., & Sabanegh, E. (2009). Role of antioxidants in the treatment of male infertility. International Journal of Urology, 16(5), 449-457.
Neuringer, M., Anderson, G. J., & Connor, W. E. (1988). The essentiality of n-3 fatty acids for the development and function of the retina and brain. Annual Review of Nutrition, 8(1), 517-541.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
Fassett, R. G., & Coombes, J. S. (2011). Astaxanthin: a potential therapeutic agent in cardiovascular disease. Marine Drugs, 9(3), 447-465.
Kothari, S., Thompson, A., Agarwal, A., & du Plessis, S. S. (2010). Free radicals: their beneficial and detrimental effects on sperm function. Indian Journal of Experimental Biology, 48(5), 425-435.

# KNOWLEDGE SUMMARY: CHAPTER 2 – THE GENETIC FRONT: HALTING DNA FRAGMENTATION
## I. INTRODUCTION: THE VULNERABLE PAYLOAD
* **The Biological Paradox:** The spermatozoon sacrifices its intrinsic defense mechanisms to achieve maximum hydrodynamic efficiency, leaving the critical paternal genome unprotected.
* **The Cytoplasmic Extrusion:** * **Mechanism:** The maturing gamete forcefully extrudes its cytoplasmic volume (residual bodies) to eliminate spatial bulk, minimizing drag and optimizing the mass-to-propulsion ratio.
* **The Chromatin Condensation:**
* **Mechanism:** Somatic histones are replaced by specialized protamines. This tightly winds and crushes the DNA into a highly compact, crystalline-like structure within the sperm head, prioritizing spatial economy.
* **The Enzyme Depletion:**
* **Biochemical Consequence:** The discarded cytoplasm is the primary reservoir for endogenous antioxidant enzymes (Superoxide Dismutase, Catalase, Glutathione Peroxidase). The mature sperm operates in a state of near-zero thermodynamic resistance.
* **The Payload Isolation and Proximity:**
* **Structural Vulnerability:** Because of the missing cytoplasm, the condensed chromatin rests mere nanometers from the external plasma membrane.
* **The PUFA Threat:** The adjacent membrane is dense with highly reactive Polyunsaturated Fatty Acids (PUFAs). Any lipid peroxidation cascade at the boundary instantly propagates inward to the adjacent genetic code.
## II. THE PHYSICS OF OXIDATIVE CLEAVAGE
* **The ROS Infiltration:**
* **The Hydroxyl Radical Threat:** Highly volatile, uncharged radicals (hydroxyl, hydrogen peroxide) easily bypass the compromised phospholipid bilayer.
* **The Nuclear Penetration:** The absence of a robust nuclear envelope offers zero physical resistance, allowing ROS direct physical contact with the DNA-protamine complex.
* **The Guanine Oxidation:**
* **Thermodynamic Vulnerability:** Guanine possesses the lowest oxidation potential among nucleotide bases, making it the primary target for electron extraction.
* **Formation of 8-OHdG:** A hydroxyl radical abstracts a hydrogen atom and forces the addition of a hydroxyl group at the 8th carbon position of the guanine ring. This converts it to 8-hydroxy-2’-deoxyguanosine (8-OHdG), the gold-standard biomarker for oxidative DNA damage.
* **The Phosphodiester Backbone Severing:**
* **Helix Destabilization:** The bulky 8-OHdG molecule creates internal spatial tension, weakening hydrogen bonds with the complementary cytosine base.
* **Single-Strand Breaks (SSBs):** Continued electron extraction forces the physical cleavage of phosphodiester bonds on one side of the double helix.
* **Double-Strand Breaks (DSBs):** Proximal SSBs on opposite strands, or massive high-energy ROS strikes, completely sever the chromosome, physically dismantling the genetic blueprint.
## III. DECONSTRUCTING DFI: THE INVISIBLE SABOTEUR
* **The Diagnostic Blind Spot:**
* **The Macroscopic Illusion:** Standard spermiograms assess count, gross morphology, and motility (the “vehicle”). They are entirely incapable of inspecting the internal DNA integrity (the “cargo”).
* **The False Positive of Vitality:** A cell can exhibit vigorous forward progression while harboring lethal double-strand breaks, masking its true reproductive incompetence.
* **Molecular Metrics:** Diagnosis mandates the measurement of the DNA Fragmentation Index (DFI) using advanced nuclear assays (e.g., TUNEL, Halosperm).
* **The Fertilization Paradox:**
* **Successful Penetration:** High-DFI sperm with intact outer boundaries can successfully navigate the reproductive tract and penetrate the oocyte.
* **The ICSI Bypass:** Assisted Reproductive Technologies (ART) explicitly bypass natural selection barriers, often directly injecting highly fragmented DNA into the egg.
* **The Oocyte Repair Threshold:** Maternal repair transcripts can fix minor damage. However, widespread double-strand breaks easily exceed this threshold, forcing the incorporation of permanent structural errors into the embryonic genome.
* **The Embryonic Arrest:**
* **Late-Stage Failure:** Damage manifests critically around Day 3 of development, when the embryonic genome activates and reads the paternal contribution.
* **Clinical Fallout:** Corrupted code physically halts cellular division, leading directly to poor blastocyst development, total embryonic arrest, or Recurrent Pregnancy Loss (RPL). It is the biophysical root of “idiopathic” infertility.

## IV. THE 15:1 AMPLIFIER: EXHAUSTING THE SEMINAL DEFENSES
* **The Seminal Plasma Shield:**
* **Extracellular Antioxidant Matrix:** Seminal fluid is engineered with non-enzymatic and enzymatic scavengers to neutralize external ROS.
* **Systemic Dependence:** This fluid relies entirely on the overall systemic metabolic and nutritional status of the male body for its antioxidant supply.
* **The Pro-Inflammatory Cascade:**
* **Arachidonic Acid Overload:** The 15:1 Omega-6 dietary excess forces the systemic over-accumulation of Arachidonic Acid (AA).
* **The Eicosanoid Shift:** Cyclooxygenase (COX-2) enzymes constantly cleave AA into pro-inflammatory eicosanoids (Prostaglandin E2) and cytokines (IL-6).
* **Background ROS:** Chronic, low-grade systemic inflammation constantly generates pathological levels of background ROS across the reproductive axis.
* **The Defense Depletion:**
* **Physiological Triage:** The body consumes its circulating antioxidant reserves to fight the systemic inflammatory fire.
* **Seminal Deficit:** The localized reproductive shield is starved of reinforcements. Localized ROS face zero thermodynamic opposition, allowing free infiltration to execute oxidative cleavage.
## V. THE ASTAXANTHIN INTERCEPTION: DEPLOYING THE THERMODYNAMIC SHIELD
* **The Blood-Testis Barrier Breach:**
* **Hydrophilic Failure:** Sertoli cell tight junctions strictly block water-soluble antioxidants (like Vitamin C).
* **Lipophilic Supremacy:** Astaxanthin dissolves into circulating lipoprotein carriers, effortlessly bypassing the hydrophobic gating of the Blood-Testis Barrier (BTB) to saturate the interstitial spaces.
* **The Transmembrane Anchoring Mechanism:**
* **Dimensional Perfection:** The Astaxanthin carbon backbone is exactly 30 Angstroms long, perfectly matching the width of the hydrophobic core of the bilayer.
* **Polar Anchors:** Terminal hydroxyl and keto functional groups physically lock onto polar phosphate lipid heads on both the inner and outer membrane surfaces.
* **Perimeter Firewall:** The molecule spans the entire interior, acting as a continuous vertical strut and physical firewall precisely where ROS attempt to penetrate.
* **The Targeted ROS Quenching:**
* **Conjugated Electron Cloud:** Alternating double/single carbon bonds create a highly active, resonating electron cloud that acts as a thermodynamic trap.
* **Resonance Dissipation:** Penetrating hydroxyl radicals are intercepted. Astaxanthin absorbs their kinetic energy and dissipates it safely as low-grade heat through continuous electron resonance.
* **Zero-Phase-Transition:** Because it utilizes internal resonance instead of electron donation, Astaxanthin never degrades into a pro-oxidant.
* **The 2-4:1 Synergistic Reconfiguration:**
* **The Biochemical Safe Zone:** By extinguishing the lipid peroxidation cascade, Astaxanthin creates a secure microenvironment. This vanguard function ensures that the highly fragile ALA and DHA molecules from the Keyora 2-4:1 matrix can physically integrate and rebuild the membrane without immediate oxidative shattering.
## VI. THE ACADEMIC VALIDATION OF GENETIC PRESERVATION
* **The Aitken Validation (DNA Damage):**
* **Source:** Aitken & Baker (2006), *Molecular and Cellular Endocrinology*.
* **Findings:** Empirically established that elevated ROS are the primary, direct mechanical drivers of spermatozoal DNA damage and the elevation of DFI. Validated the mechanism of “oxidative cleavage.”
* **The Comhaire Validation (ROS Quenching & Shielding):**
* **Source:** Comhaire et al. (2005), *Asian Journal of Andrology*.
* **Intervention:** Double-blind, placebo-controlled trial. 16mg daily dosage of Natural Astaxanthin in idiopathic subfertile men for three months.
* **Biochemical Findings:** A rapid, statistically significant decrease in measurable concentrations of seminal ROS compared to the placebo cohort.
* **The Ultimate Clinical Verdict (Conception Rates):**
* **Placebo Baseline:** The unshielded placebo cohort achieved only a 10.5% conception rate.
* **Intervention Surge:** The 16mg Astaxanthin cohort surged to a statistically profound 54.5% conception rate.
* **Consensus:** The deployment of the Astaxanthin thermodynamic shield is clinically proven to halt oxidative fragmentation, preserve the genetic blueprint, and fundamentally optimize real-world conception outcomes.

Chapter 3: The Pathobiology Of Spermatozoal Bioenergetics:
Oxidative Stress And ATP Depletion
Analyzing the mechanistic failure of flagellar motility and the targeted Astaxanthin intervention for metabolic optimization.
A flawlessly calibrated, highly fluid liquid crystal plasma membrane and an impeccably preserved, un-cleaved genetic payload are rendered entirely biologically useless if the spermatozoon cannot physically and forcefully propel itself to its final destination.
The perilous transit through the viscous, highly selective environment of the female reproductive tract – from the acidic vaginal vault, through the cervical mucus, into the uterine cavity, and ultimately into the fallopian tubes – requires the generation of an immense, unrelenting, and highly coordinated mechanical force. This continuous physical propulsion is driven exclusively by the rapid, undulating kinetics of the flagellar beat.
To sustain this rigorous mechanical output over a prolonged duration, the biological system demands a massive, uninterrupted, and strictly localized supply of Adenosine Triphosphate.
We must now shift our forensic biophysical analysis inward to precisely examine the highly specialized, aggressively engineered anatomical structure entirely responsible for meeting this absolute bioenergetic demand: the concentrated mitochondrial engine located specifically within the spermatozoal midpiece.

1. The Midpiece Architecture
The Precise Anatomical Concentration Of Cellular Power Plants
To achieve the necessary kinetic energy output required for forward progressive motility, the cellular energy infrastructure must be radically redesigned from the foundational level.
The spatial arrangement of the power plants is as critical as their metabolic function.
I. The Helical Formation:
In standard, stationary somatic cells, mitochondria are typically dispersed relatively evenly throughout the expansive volume of the cellular cytoplasm, providing a diffuse energy grid.
However, the mature spermatozoon operates under extreme spatial constraints and intense kinetic demands. During spermiogenesis, the mitochondria are aggressively repositioned and restructured.
They are not scattered randomly; instead, they are tightly and meticulously coiled into a highly organized, dense, interlocking helical sheath that completely encases the proximal portion of the axoneme – the central microtubule core of the flagellum – defining the exact structural boundaries of the anatomical midpiece.
II. The Structural Proximity:
The fundamental biophysical reason for executing this extreme, dense architectural reorganization is strictly mechanical efficiency.
The mitochondria must be positioned physically adjacent and intimately connected to the outer dense fibers and the underlying microtubular sliding apparatus of the flagellum.
This ultra-close proximity is explicitly designed to minimize the spatial diffusion distance of the newly synthesized Adenosine Triphosphate molecules.
By eliminating the transit time across a diffuse cytoplasm, the system ensures the immediate, rapid, and highly efficient transfer of vital kinetic energy directly to the mechanical motor required for the whipping movement.
III. The Metabolic Density:
The immediate physiological consequence of packing dozens of mitochondria into such a tightly confined, helical formation is the creation of a profoundly localized zone of extreme metabolic activity.
The spermatozoal midpiece functions as a hyper-dense energy reactor, operating continuously at maximum, high-output metabolic capacity to fuel the relentless propulsion system.
This localized density generates intense thermodynamic pressure within a severely restricted microscopic volume.

2. The ATP Demand Of Flagellar Beating
The Thermodynamic Cost Of Forward Linear Velocity
The architectural density of the midpiece is entirely dictated by the severe, unyielding thermodynamic cost of propelling the biological missile through a highly resistant fluid medium.
I. The Mechanical Whipping Motion:
The complex mechanics of the flagellar beat do not occur spontaneously; they are the result of highly coordinated, active structural shifting.
The continuous, undulating whipping motion is generated by the rapid, sequential sliding of the internal axonemal microtubules against one another, facilitated by dynein motor proteins.
This specific, repetitive mechanical action is strictly and completely dependent on the continuous, rapid, and high-volume hydrolysis of Adenosine Triphosphate molecules to force the physical conformational changes within the motor proteins.
II. The Oxidative Phosphorylation Reliance:
To successfully meet this extreme, unrelenting localized energy demand, the mature spermatozoon relies heavily on the complex process of oxidative phosphorylation occurring deep within the inner mitochondrial membranes of the helical sheath.
While glycolysis provides some supplemental energy, oxidative phosphorylation is the primary, highly efficient, but thermodynamically demanding metabolic pathway required to generate the massive quantities of Adenosine Triphosphate strictly necessary to sustain high-velocity linear progression over the required clinical duration.
III. The Zero-Tolerance Threshold:
The absolute biophysical conclusion regarding this energy dynamic is that there is zero tolerance within the biological system for any metabolic interruption.
The motor proteins possess no significant internal reserves of energy.
Any disruption, suppression, or stalling in the mitochondrial Adenosine Triphosphate output, regardless of the cause, directly, immediately, and proportionally translates to a catastrophic cessation of flagellar motility, resulting directly in the clinical manifestation of mechanical paralysis.

3. The Inherent Oxidative Vulnerability
The Biophysical Paradox Of A High-Output Engine Lacking Internal Defenses
The very anatomical and metabolic engineering that enables the immense power output of the spermatozoon simultaneously establishes its most critical structural vulnerability, creating a highly dangerous biological paradox.
I. The Baseline ROS Production:
Because the mitochondria within the midpiece are operating at maximum capacity to sustain the flagellar beat, the intense rate of oxidative phosphorylation inherently and unavoidably generates a steady, baseline level of Reactive Oxygen Species.
These radicals, specifically superoxide anions, are produced as a natural, unavoidable thermodynamic byproduct of electron leakage occurring directly along the highly active electron transport chain within the inner mitochondrial membrane.
II. The Cytoplasmic Deficit:
Under normal physiological conditions in a somatic cell, this baseline internal radical production is rapidly neutralized by the surrounding enzymatic networks.
However, the spermatozoon must contend with the severe structural sacrifice of maturation.
Because the gamete has forcefully extruded its entire cytoplasmic volume to minimize mass, the actively firing mitochondria are now operating completely without the robust, protective buffer of endogenous antioxidant enzymes – such as localized Superoxide Dismutase and Catalase – typically found abundantly in the cytoplasm of standard cells.
III. The Imminent Threat:
Consequently, the spermatozoal midpiece represents a profound biophysical paradox: it is an exceptionally high-output biological engine deliberately operating under extreme, continuous thermodynamic stress, yet it completely and structurally lacks the basic internal safety mechanisms required to manage its own metabolic exhaust.
It exists in a perpetual state of precarious equilibrium, rendering the entire propulsion system acutely and disproportionately vulnerable to any external inflammatory disruption or systemic oxidative amplification.

3.1 The 15:1 Catalyst:
Destabilizing The Inner Membrane
How The Modern Dietary Lipid Imbalance Physically Alters Cardiolipin Architecture And Triggers Mitochondrial Electron Leakage
The maximum thermodynamic efficiency of Adenosine Triphosphate production, an absolute prerequisite for sustaining the intense mechanical force of the flagellar beat, is entirely and rigorously dependent upon the flawless structural integrity of the Inner Mitochondrial Membrane.
When the systemic physiological environment is subjected to the modern dietary ratio of Omega-6 to Omega-3, reaching the highly toxic 15:1 to 20:1 threshold, this imbalance does not merely manifest as general metabolic sluggishness.
Instead, it acts as a direct, highly specific biochemical catalyst for profound biophysical destabilization deep within the cell. This is not a generalized cellular failure or a simple lack of fuel; it is a localized, mechanical disruption of the highly specialized mitochondrial lipid matrix.
Specifically, this toxic ratio relentlessly targets and structurally corrupts a unique, functionally critical phospholipid known as cardiolipin, initiating a cascade of architectural failures that effectively turns the biological engine against itself.

1. The Cardiolipin Dependency
The Critical Phospholipid Of The Mitochondrial Engine
To understand the exact mechanics of this intracellular failure, one must first isolate the precise lipid component responsible for maintaining the engine’s structural tolerances.
The Inner Mitochondrial Membrane relies upon a molecular foundation found virtually nowhere else in the human body.
A. The Unique Phospholipid:
Cardiolipin is a highly specialized, functionally unique phospholipid molecule that is synthesized and utilized almost exclusively within the confines of the Inner Mitochondrial Membrane.
Distinct from standard bilayer lipids, it possesses a complex, dimeric structure featuring four distinct acyl chains, which grants the molecule a pronounced cone – shaped spatial geometry.
This specific physical shape is an absolute evolutionary requirement, providing the necessary mechanical leverage and structural tension required to maintain the extreme, tightly folded optimal membrane curvature characteristic of the highly active mitochondrial cristae.
B. The Protein Complex Anchor:
The function of cardiolipin extends far beyond maintaining simple membrane curvature. It acts as the primary physical anchor and the essential structural glue for the massive enzyme complexes that constitute the Electron Transport Chain.
By binding tightly to these specific proteins, cardiolipin strictly governs their spatial orientation within the lipid matrix, keeping the individual complexes locked in perfect, nanometer – precise spatial alignment.
This rigid structural organization is an absolute prerequisite for ensuring the rapid, highly efficient transfer of electrons down the sequence.
C. The PUFA Requirement:
For cardiolipin to execute this vital anchoring function correctly without shattering under thermodynamic stress, its four distinct acyl chains must be highly enriched with very specific, heavily kinked Polyunsaturated Fatty Acids.
These specific lipid components, particularly those derived from the Omega-3 pathway, provide the necessary physical flexibility and massive steric hindrance required to create a highly dynamic, responsive microenvironment directly immediately surrounding the respiratory complexes.
D. The Oxidative Target:
However, this heavy structural reliance on multiple Polyunsaturated Fatty Acids establishes a profound biophysical vulnerability.
While these double bonds make cardiolipin structurally perfect for facilitating rapid, high – volume energy transfer, they simultaneously render the molecule highly susceptible to aggressive oxidative damage.
If the delicate lipid composition is artificially altered or subjected to an unmitigated radical strike, the entire structural integrity of the molecule, and therefore the alignment of the respiratory chain, is immediately compromised.

2. The Competitive Enzymatic Hijack
The Forced Substitution Of Structural Lipids
The structural corruption of cardiolipin does not begin within the mitochondria itself; it originates upstream, driven entirely by the systemic macronutrient imbalance overwhelming the localized lipid synthesis machinery.
A. The Shared Desaturase Bottleneck:
The exact objective biochemical mechanism driving this mitochondrial collapse is identical to the mechanism paralyzing the outer plasma membrane: competitive inhibition.
The massive 15:1 volumetric surplus of dietary Omega-6 circulating within the system physically overwhelms the shared enzymatic infrastructure, specifically creating a severe, localized competitive blockade directly at the critical Delta-5 and Delta-6 desaturase enzymes.
B. The Omega-3 Starvation:
The immediate consequence of this persistent competitive blockade is localized nutritional starvation.
By monopolizing the essential desaturase enzymes, the massive Omega-6 payload effectively starves the highly active testicular microenvironment of the vital, highly fluid, heavily kinked Omega-3 fatty acids that are strictly and biologically necessary for the optimal, functional construction of the specialized cardiolipin molecules.
C. The Forced Incorporation:
Deprived of the optimal biological building materials, yet compelled by the physiological imperative to rapidly synthesize new mitochondria for the developing spermatozoa, the localized cellular machinery is forced to compromise.
Dictated entirely by this enzymatic hijack, the mitochondria must synthesize new cardiolipin molecules utilizing disproportionately massive amounts of the only available substrates: highly rigid, pro – inflammatory Omega-6 fatty acids, most notably the tightly packing Arachidonic Acid.
D. The Structural Distortion:
The physical conclusion of this forced molecular substitution is absolute architectural degradation.
Because Arachidonic Acid lacks the necessary molecular kinking, the newly formed cardiolipin molecules are rigid and inflexible. Consequently, the Inner Mitochondrial Membrane forcefully loses its optimal, dynamic fluidity.
The carefully orchestrated, precision architecture of the folded cristae becomes physically distorted and stiffened, fatally compromising its critical function as a thermodynamic insulator.

3. The ETC Misalignment And Electron Leak
The Genesis Of The Intracellular Oxidative Storm
The stiffening of the cardiolipin matrix translates directly into a profound mechanical failure of the primary energy – generating apparatus.
A distorted foundation cannot support a precision engine.
A. The Spatial Disruption:
Because the newly synthesized, Arachidonic Acid – heavy cardiolipin molecules are physically rigid and geometrically distorted, they fail entirely in their primary biological role as the structural anchors of the respiratory chain.
The embedded enzyme complexes of the Electron Transport Chain, deprived of their flexible, precisely calibrated foundation, inevitably suffer a microscopic, yet functionally catastrophic, spatial misalignment within the rigidified lipid bilayer.
B. The Inefficient Transfer:
This spatial disruption initiates the exact biophysical failure of the biological motor. The Electron Transport Chain operates on strict quantum mechanical tolerances.
When the complexes are misaligned by even a fraction of a nanometer, the precise spatial distance strictly required for electrons to successfully and efficiently jump from one specific complex to the next is critically altered.
This spatial expansion severely reduces the overall thermodynamic efficiency of the entire sequence.
C. The Escape Of Electrons:
As the highly energized electrons are continuously pumped into the sequence by the rapid hydrolysis required for flagellar propulsion, they encounter these widened spatial gaps caused by the structural misalignment.
Unable to safely bridge the expanded distance to the subsequent receptor protein, a significant percentage of these volatile electrons prematurely escape the defined transport sequence before ever successfully reaching the final stabilizing cytochrome c oxidase complex.
D. The Superoxide Formation:
The ultimate, lethal conclusion of this mechanical failure occurs the millisecond these electrons escape the chain. These highly reactive, un-sequestered electrons immediately and violently react directly with the ambient molecular oxygen naturally present within the mitochondrial matrix.
This unregulated chemical reaction instantly generates massive localized bursts of the highly destructive Superoxide Anion. The critical internal biological engine, crippled by its corrupted lipid architecture, has officially begun to rapidly self – destruct from the inside out.

3.2 The Lipid Peroxidation Cascade:
The Engine Meltdown
The Physical Destruction Of The Inner Mitochondrial Membrane, The Collapse Of The Thermodynamic Gradient, And The Subsequent Cessation Of ATP Synthesis
The generation of the highly volatile superoxide anion, directly caused by the mechanical misalignment of the Electron Transport Chain within a rigidified cardiolipin matrix, serves as the critical biochemical ignition point.
Because the mature spermatozoon has fundamentally sacrificed its endogenous enzymatic buffering capacity – specifically the localized mitochondrial Superoxide Dismutase – these newly generated localized bursts of Reactive Oxygen Species cannot be neutralized or safely sequestered.
Left entirely unopposed within the densely packed helical sheath of the midpiece, these radicals initiate a devastating, incredibly rapid, and self – propagating biophysical chain reaction.
This localized oxidative storm fundamentally and systematically dismantles the critical thermodynamic insulation of the inner mitochondrial membrane. It triggers a catastrophic sequence of structural failures that rapidly degrades the biological engine, ultimately resulting in a complete, irreversible metabolic meltdown that permanently paralyzes the gamete.

1. The Peroxidation Chain Reaction
The Progressive Fracturing Of The Cardiolipin Bilayer
The structural destruction of the mitochondrial engine is not a generalized decay; it is a highly specific, targeted chemical attack directly upon the structural integrity of the surrounding membrane architecture.
Firstly, The Radical Abstraction:
The initial, catastrophic step of this cascade occurs when the newly formed superoxide anion, or its highly aggressive downstream derivatives such as the hydroxyl radical, execute a direct chemical strike against the surrounding lipid matrix.
These volatile radicals physically and violently steal weakly bound hydrogen atoms directly from the vulnerable methylene groups situated between the multiple carbon double bonds of the Polyunsaturated Fatty Acids that comprise the critical cardiolipin molecules.
Secondly, The Propagation Phase:
This initial, violent atomic theft does not merely damage a single lipid; it creates a highly unstable, highly reactive lipid peroxyl radical within the membrane.
To restabilize itself, this newly formed lipid radical immediately attacks adjacent, intact fatty acid chains, stealing their hydrogen atoms and propagating the instability.
This initiates a rapid, aggressive, and entirely self – sustaining biochemical chain reaction that surges horizontally across the entire surface of the inner mitochondrial membrane.
Thirdly, The Structural Fracturing:
As the vulnerable carbon double bonds are systematically targeted and destroyed by this cascading reaction, the precise physical structure of the cardiolipin molecules fundamentally fractures.
The highly kinked spatial geometry required for membrane integrity collapses, creating microscopic pores, physical tears, and structural defects throughout the previously pristine, highly impermeable lipid bilayer of the cristae.
Fourthly, The Toxic Byproducts:
This violent lipid peroxidation cascade is not merely structurally destructive; it is heavily toxic.
As the lipid chains shatter, the reaction continuously generates highly reactive secondary aldehyde byproducts, most notably the toxic compound Malondialdehyde.
These aggressive aldehydes rapidly cross – link and physically denature adjacent mitochondrial proteins, further compounding the structural degradation of the entire biological engine.

2. The Proton Leakage
The Breach Of The Thermodynamic Insulation
The immediate biophysical consequence of this progressive structural fracturing is the catastrophic failure of the membrane’s primary physiological function: acting as an absolute electrical and thermodynamic barrier.
Firstly, The Insulation Requirement:
For the mitochondrial engine to function, the inner mitochondrial membrane must act as a perfect, uncompromising electrical insulator.
As the Electron Transport Chain operates, it aggressively pumps protons directly into the confined intermembrane space.
This impermeable lipid barrier traps these protons, creating a massive, highly concentrated, and highly pressurized thermodynamic gradient between the two sides of the membrane.
Secondly, The Microscopic Breaches:
The physical integrity of this critical insulator is entirely destroyed by the escalating oxidative cascade.
The microscopic pores, structural tears, and lipid fractures aggressively created by the continuous lipid peroxidation chain reaction physically and completely breach this essential, highly calibrated insulation layer, effectively puncturing the biological pressure vessel.
Thirdly, The Uncontrolled Backflow:
The direct biophysical failure occurs immediately upon structural breaching. The massive concentration of trapped, highly pressurized protons forcibly begins to leak back across the damaged, newly porous membrane matrix.
Critically, this uncontrolled backflow completely bypasses the designated, highly regulated channel of the ATP synthase complex, flowing through the oxidative fractures instead.
Fourthly, The Loss Of Gradient:
The ultimate consequence of this unmitigated proton leak is the rapid, catastrophic dissipation of the carefully built thermodynamic pressure.
The essential driving force required for continuous energy production is vented uselessly back into the mitochondrial matrix through these structural leaks, entirely destroying the necessary biological gradient required to turn the mechanical rotor of the enzyme complex.

3. The Transmembrane Potential Collapse
The Depolarization Of The Biological Battery
The physical venting of the proton gradient directly and immediately translates to a measurable, catastrophic collapse in the electrical operating capacity of the mitochondrial engine.
Firstly, The Delta Psi m Metric:
In clinical andrology and biophysics, the overall operational health of the mitochondrial engine is objectively quantified by measuring the mitochondrial Transmembrane Potential, scientifically designated as Delta Psi m.
This critical metric represents the absolute, measurable electrical charge maintained across the inner membrane, accurately reflecting the total stored thermodynamic energy capacity of the biological battery.
Secondly, The Depolarization Event:
The massive, uncontrolled proton leakage initiated by lipid peroxidation causes a rapid, uncontrolled drop in this vital electrical charge.
As the protons flow backward through the fractured lipid matrix, the electrical differential is violently equalized, leading directly to the severe, systemic depolarization of the entire inner mitochondrial membrane within the flagellar midpiece.
Thirdly, The Battery Depletion:
The biophysical reality of this event is absolute.
A severely depolarized mitochondrion is biologically and functionally equivalent to a completely dead, exhausted battery.
It possesses absolutely no stored thermodynamic energy, no operational pressure gradient, and therefore possesses zero functional capacity to perform the intense mechanical work required for linear propulsion.
Fourthly, The Irreversible State:
Crucially, because the spermatozoon lacks the internal genetic and enzymatic machinery required to actively repair significant structural membrane damage or synthesize new mitochondria, this severe depolarization rapidly becomes an irreversible biophysical state.
Without immediate, highly targeted external intervention to decisively halt the lipid peroxidation, the engine permanently fails.

4. The Clinical Manifestation: Asthenozoospermia
The Mechanical Paralysis Of The Flagellar Propulsion System
The microscopic, intracellular collapse of the thermodynamic gradient ultimately manifests as a macroscopic, highly visible, and clinically devastating mechanical failure of the entire gamete.
Firstly, The ATP Synthase Shutdown:
The generation of localized energy is a purely mechanical process driven by pressure.
Without the intense driving force provided by the high Transmembrane Potential, the massive ATP synthase rotor physically and entirely ceases to turn.
The necessary continuous, high – volume biological production of Adenosine Triphosphate halts immediately across the entire mitochondrial sheath.
Secondly, The Energy Starvation:
The immediate consequence of this metabolic shutdown is severe, localized energy starvation.
The underlying dense fibers and the actively sliding microtubules of the flagellar axoneme, which entirely rely on this continuous, immediate ATP transfer, are suddenly and completely starved of their exclusive, vital kinetic energy source.
Thirdly, The Flagellar Paralysis:
The objective mechanical outcome of this intracellular starvation is complete structural arrest.
Without the necessary ATP to force the motor proteins to shift, the rapid, powerful whipping motion of the flagellum abruptly slows and eventually stops entirely.
The entire highly engineered propulsion system is rendered completely paralyzed by its own internal metabolic collapse.
Fourthly, The Root Cause Identified:
This precise, sequential biophysical collapse – originating from cardiolipin distortion, accelerating through uncontrolled lipid peroxidation and proton leakage, resulting in severe depolarization, and terminating in the absolute cessation of ATP synthesis – is the exact, objective, biochemical root cause of Asthenozoospermia.
It provides the definitive mechanical explanation for the profoundly low sperm motility universally observed in modern fertility clinics, proving the failure is a structural thermodynamic collapse, not merely a lack of cellular vitality.

3.3 The Astaxanthin Vanguard:
Deploying The Mitochondrial Shield
The Biophysical Deployment Of The Absolute Thermodynamic Protagonist To Penetrate The Midpiece, Anchor The Inner Membrane, And Quench The Electron Leak
The bioenergetic engine of the spermatozoon is currently operating in a state of active, catastrophic meltdown.
The highly specialized cardiolipin molecules are rapidly fracturing under oxidative stress, the critical thermodynamic protons are relentlessly leaking across the compromised boundaries, and the vital Adenosine Triphosphate supply to the flagellar motor has been completely severed.
In this severe state of intracellular crisis, conventional, water – soluble antioxidants circulating within the peripheral bloodstream are rendered entirely obsolete.
They cannot physically penetrate the highly restrictive Blood – Testis Barrier, let alone successfully navigate the complex, lipid – dense double – membrane architecture of the failing mitochondria.
To decisively halt the lipid peroxidation cascade, arrest the uncontrolled proton leak, and successfully reboot the biological engine, a highly specific, engineered lipophilic intervention is strictly required.
Astaxanthin now formally enters the microscopic bioenergetic battlefield as the absolute protagonist of the Keyora protocol, explicitly deploying a permanent, impenetrable mitochondrial shield to rescue the thermodynamic core.

1. The Blood – Testis Barrier Penetration
Bypassing The Anatomical Isolation Of The Reproductive Tract
The primary tactical objective of this bioenergetic intervention is to successfully navigate the complex anatomical architecture of the male reproductive system to deliver the protective payload directly to the site of cellular maturation and metabolic failure.
I. The Failure Of Hydrophilic Defense:
To fully grasp the ingenuity of the Astaxanthin intervention, one must first recognize the absolute biophysical limitation of standard therapeutic approaches.
The seminiferous tubules, the exact physical location where the highly vulnerable spermatozoa undergo maturation, are strictly isolated from the systemic circulation by the Blood – Testis Barrier.
This barrier utilizes an intricate, virtually impenetrable network of dense tight junctions formed by adjacent Sertoli cells to selectively filter incoming molecules.
This stringent anatomical gating specifically and aggressively physically blocks the entry of hydrophilic, water – soluble molecules like Vitamin C and standard bioflavonoids, rendering these common interventions entirely useless for deep, localized testicular defense against internal mitochondrial collapse.
II. The Lipophilic Supremacy:
Astaxanthin effortlessly overrides this stringent biological blockade through its extreme, inherent chemical lipophilicity.
Classified as a highly complex, fat – soluble marine xanthophyll carotenoid, its precise molecular nature is perfectly adapted for integration into dense lipid – based biological systems.
This supreme lipophilic advantage allows the Astaxanthin molecule to seamlessly and completely dissolve into circulating high – density and low – density lipoprotein carriers within the bloodstream.
By utilizing these natural, endogenous lipid transport vehicles, the molecule effortlessly bypasses the hydrophobic tight junctions of the Blood – Testis Barrier, utilizing the body’s own lipid pathways as a direct, unhindered transit corridor.
III. The Interstitial Saturation:
This precise lipophilic transit mechanism guarantees that the intervention does not merely circulate in the periphery but successfully penetrates the deep reproductive tissues.
By continuously riding the lipid transport network, Astaxanthin achieves exceptionally high, targeted therapeutic concentrations directly within the isolated testicular microenvironment.
It effectively saturates the interstitial spaces and the localized seminal fluid, successfully surrounding the developing seminiferous tubules with a highly potent, concentrated reserve of advanced thermodynamic defense molecules ready for immediate cellular deployment and mitochondrial rescue.
IV. The Plasma Membrane Integration:
Upon physically reaching the immediate vicinity of the compromised spermatozoon, Astaxanthin executes its initial cellular infiltration maneuver with absolute biophysical precision.
Because of its profound, innate affinity for lipid structures, it entirely bypasses the need for specialized, highly regulated membrane transport proteins or complex cellular receptors.
The molecule simply and directly partitions itself into the dense, lipid – rich matrix of the outer plasma membrane, effortlessly crossing the first cellular threshold without expending any of the cell’s rapidly depleting energy reserves.

2. The Targeted Mitochondrial Localization
Navigating To The Bioenergetic Core Of The Midpiece
Breaching the outer plasma membrane is merely the first step.
The molecule must now actively navigate the internal architecture of the cell to locate and protect the failing biological engine situated within the flagellar midpiece.
I. The Cytoplasmic Transit:
Although the mature spermatozoon has forcefully extruded the vast majority of its cytoplasmic volume during its structural maturation, a minimal, highly concentrated layer of intracellular fluid remains.
Astaxanthin must navigate this restricted cytoplasmic space, moving systematically from the outer cellular boundary toward the areas of highest metabolic demand and most intense oxidative stress.
Its lipophilic nature allows it to glide seamlessly along the remaining internal lipid rafts and structural micro-filaments toward the midpiece.
II. The Affinity For High – Activity Membranes:
The directed movement of Astaxanthin is not random; it is governed by strict thermodynamic principles and physical attraction.
The molecule possesses a profound, innate biophysical affinity for highly active, lipid – dense biological membranes.
Because the helical mitochondrial sheath in the midpiece represents the absolute highest concentration of localized lipid membranes and metabolic activity within the entire gamete, it exerts a powerful structural draw, specifically pulling the Astaxanthin molecules toward the epicentre of the bioenergetic crisis.
III. The Outer Membrane Penetration:
Upon arriving at the tightly coiled mitochondrial sheath, the Astaxanthin molecule encounters the first layer of the organelle: the Outer Mitochondrial Membrane.
Because this outer boundary is relatively porous and designed to facilitate the rapid exchange of metabolic substrates and newly synthesized Adenosine Triphosphate, the highly lipophilic Astaxanthin molecule effortlessly penetrates this initial barrier, moving progressively deeper into the complex, failing architecture of the biological engine.
IV. The Inner Membrane Destination:
Following this initial organelle penetration, Astaxanthin reaches its ultimate, critical biophysical target: the highly folded, intensely active, and currently fracturing Inner Mitochondrial Membrane.
This specific, cardiolipin – rich boundary is the exact, microscopic site of the embedded Electron Transport Chain, the origin of the uncontrolled electron leak, and the absolute epicenter of the localized oxidative meltdown threatening the entire propulsion system.

3. The 30-Angstrom Transmembrane Anchoring
The Structural Integration Of The Thermodynamic Shield
Once inside the critical lipid matrix of the Inner Mitochondrial Membrane, the Astaxanthin molecule assumes a highly specific, mechanically rigorous physical orientation designed to maximize its structural and protective capabilities.
I. The Dimensional Perfection:
The true biophysical brilliance of the Astaxanthin molecule, and its unparalleled efficacy in mitochondrial defense, is fundamentally rooted in its exact, unvarying physical dimensions.
The linear carbon backbone of the molecule possesses a precise, measurable length of approximately 30 Angstroms.
This specific geometric measurement represents an absolute, mathematically perfect spatial match for the exact cross – sectional width of the internal hydrophobic core of the inner mitochondrial bilayer, ensuring a flawless architectural fit.
II. The Polar Anchoring Mechanism:
To ensure this perfect spatial alignment is permanently maintained despite the intense mechanical stress and rapid localized fluid dynamics of the active mitochondria, Astaxanthin utilizes specialized structural modifications at both ends of its chain.
The highly reactive hydroxyl and keto functional groups located directly on its terminal ionone rings act as potent hydrophilic anchors.
As the molecule slides into position, these polar groups physically lock themselves onto the polar phosphate lipid heads located simultaneously on both the matrix-facing and intermembrane-facing surfaces of the Inner Mitochondrial Membrane.
III. The Hydrophobic Spanning:
Anchored firmly and irreversibly at both the internal and external polar boundaries, the massive, long, conjugated polyene chain of the Astaxanthin molecule perfectly spans the entire highly volatile, hydrophobic interior of the cellular membrane.
This creates a continuous, unbroken, physical molecular strut directly across the entire thickness of the vulnerable lipid bilayer, positioning the defense mechanism directly adjacent to the highly specialized, actively fracturing cardiolipin molecules.
IV. The Structural Stabilization:
The direct biophysical consequence of this precise, vertical transmembrane orientation is immense physical reinforcement.
By spanning the full depth of the Inner Mitochondrial Membrane, Astaxanthin effectively acts as rigid molecular rebar.
It physically stabilizes the architecturally distorted and compromised lipid matrix, significantly increasing the membrane’s overall mechanical resistance to further kinetic stress, preventing the structural collapse of the vital cristae folds.

4. The Thermodynamic Quenching Of The Electron Leak
The Absolute Neutralization Of The Superoxide Assault
With the physical structure stabilized by the integration of the molecule, Astaxanthin immediately activates its primary, mission – critical chemical defense mechanism to neutralize the impending oxidative engine meltdown.
I. The Proximity To The ETC:
By precisely anchoring directly across the Inner Mitochondrial Membrane, Astaxanthin strategically positions its massive, conjugated polyene chain exactly parallel and immediately adjacent to the embedded protein complexes of the Electron Transport Chain.
This rigorous spatial positioning places the antioxidant defense mechanism directly at the exact microscopic site of the pathological electron leak and subsequent superoxide generation.
II. The Interception Of Superoxide:
The massive central chain of alternating double and single carbon bonds creates a highly active, delocalized electron cloud that continuously resonates along the molecule.
This continuously active electron cloud acts as a perfectly positioned, highly efficient thermodynamic trap.
It physically intercepts the highly volatile escaped electrons and the newly formed superoxide anions the very millisecond they are generated, completely trapping them before they can initiate any destructive chemical attack against the adjacent, highly vulnerable cardiolipin molecules.
III. The Resonance Dissipation:
The biophysics of this specific quenching mechanism is the defining characteristic of the Keyora protocol.
When it intercepts a highly destructive radical, the Astaxanthin molecule rapidly absorbs the volatile oxidative energy.
However, it instantly and safely dissipates this trapped kinetic energy as low – grade, harmless microscopic heat through continuous, unbroken electron resonance back and forth along its extensive carbon chain, successfully neutralizing the threat without ever breaking its own internal molecular bonds.
IV. The Zero – Phase – Transition Guarantee:
This specific mechanism of energy dissipation establishes the absolute, uncompromising thermodynamic rule of mitochondrial defense.
Because Astaxanthin neutralizes and dissipates radical energy exclusively through internal resonance rather than through the highly dangerous donation of an electron, it is chemically impossible for the molecule to ever undergo a phase transition to become a damaging pro – oxidant itself.
It remains a permanently active, completely impenetrable shield directly within the biological engine.

5. The Preservation Of Bioenergetics
Halting The Cascade And Securing The Foundation For Recovery
The deployment of the Astaxanthin thermodynamic shield marks the critical turning point in the bioenergetic crisis, executing an immediate cessation of the destructive pathways paralyzing the cell.
I. The Peroxidation Blockade:
The most immediate, vital outcome of this targeted thermodynamic quenching is the absolute peroxidation blockade.
By aggressively and continuously neutralizing the Reactive Oxygen Species precisely at their source within the Electron Transport Chain, Astaxanthin physically and definitively halts both the initiation and the aggressive propagation of the lipid peroxidation cascade across the entire mitochondrial sheath.
II. The Insulation Preserved:
By arresting the lipid peroxidation chain reaction, this targeted interception directly prevents any further oxidative fracturing or tearing of the delicate cardiolipin bilayer.
This crucial action successfully preserves the remaining physical integrity and the critical electrical insulation properties of the Inner Mitochondrial Membrane, immediately halting the uncontrolled leakage of protons into the mitochondrial matrix.
III. The Prerequisite For Reboot:
It is a strict biophysical reality that while Astaxanthin effectively stops the ongoing oxidative damage, it does not possess the capacity to physically rebuild the already oxidized and distorted structural lipids.
However, it successfully secures the absolute biophysical prerequisite for cellular recovery: it establishes a completely safe, rigorously controlled, ROS – free thermodynamic microenvironment within the mitochondria, a state that is strictly required before any structural repair can commence.
IV. The Handoff To The Lipid Matrix:
This triumphant stabilization concludes the absolute protagonist’s primary bioenergetic arc within the Keyora intervention timeline.
With the impenetrable thermodynamic shield now firmly and permanently locked into place across the inner membrane, and the catastrophic mitochondrial oxidative fire fully extinguished, the biological stage is perfectly and securely set.
The bioenergetic environment is now stabilized, allowing the Keyora 2-4:1 matrix to safely introduce highly kinked Omega-3 lipids to physically rebuild the cardiolipin architecture and permanently reboot the flagellar engine.

3.4 The Keyora 2-4:1 Matrix:
The Bioenergetic Reconfiguration
How The Precision – Engineered Lipid Payload Executes The Structural Repair Of Cardiolipin And Optimizes Metabolic Efficiency Under The Astaxanthin Shield
The Astaxanthin vanguard has successfully infiltrated and secured the highly volatile mitochondrial engine room within the spermatozoal midpiece, explicitly deploying an absolute, permanent thermodynamic shield to physically quench the devastating electron leak.
However, while the catastrophic lipid peroxidation chain reaction has been definitively halted, the physical architecture of the Inner Mitochondrial Membrane remains in a state of severe disrepair.
The specialized cardiolipin molecules, previously rigidified by the toxic 15:1 systemic ratio and subsequently fractured by the oxidative storm, strictly require an immediate influx of pristine, highly fluid lipid substrates to facilitate complete physical reconstruction.
Operating entirely under the continuous, absolute thermodynamic protection established by the Astaxanthin shield, the Keyora matrix now systematically deploys its highly specific, engineered lipidomic payload – specifically Alpha-Linolenic Acid and Oleic Acid – to forcefully execute the structural repair of cardiolipin and fundamentally recalibrate the thermodynamic efficiency of the localized Adenosine Triphosphate production cycle.

1. The Escorted Substrate Delivery
The Safe Transit Of Highly Vulnerable Polyunsaturated Fatty Acids
The successful integration of structural lipids is contingent entirely upon their safe arrival.
The required molecules are structurally perfect for the task, but chemically highly unstable in a hostile physiological environment.
A. The PUFA Fragility:
The absolute biophysical reality of Omega-3 fatty acids, most notably the parent precursor Alpha-Linolenic Acid, is their extreme inherent fragility.
Because their specific molecular geometry requires the presence of multiple carbon – to – carbon double bonds, these vital structural lipids are incredibly delicate and highly prone to rapid, explosive lipid peroxidation if exposed to even baseline levels of systemic oxidative stress during circulation.
B. The Astaxanthin Vanguard:
To ensure the survival of this delicate payload, the systemic presence of Astaxanthin is strictly leveraged.
By maintaining high circulating concentrations, Astaxanthin actively acts as a dedicated thermodynamic escort. It physically shields the fragile, circulating Alpha-Linolenic Acid molecules from pervasive background oxidation and localized inflammatory attacks during their precarious transit through the cardiovascular system to the deep testicular interstitium.
C. The Intracellular Protection:
This critical protection protocol extends directly into the target cell.
Once the lipid payload successfully crosses into the spermatozoon, the established Astaxanthin shield anchored across the Inner Mitochondrial Membrane ensures that these incoming, highly vulnerable lipid substrates are not immediately shattered and oxidized the millisecond they enter the intense, high – metabolic zone of the flagellar midpiece.
D. The Pristine Arrival:
The ultimate, necessary conclusion of this highly coordinated, escorted delivery system is that a completely pure, entirely unoxidized, and highly functional supply of essential fatty acid substrates successfully arrives intact directly at the fractured Inner Mitochondrial Membrane.
The raw materials are successfully delivered precisely to the site of injury, completely ready for immediate enzymatic integration without introducing any new oxidative liabilities.

2. The 2-4:1 Enzymatic Override
Forcing The Metabolic Shift Toward Fluidizing Lipids
Upon arrival, the lipid payload must forcefully instruct the local metabolic machinery to abandon its pathological production of rigid structural components and resume the synthesis of optimal gametic architecture.
A. The Precision Ratio:
The Keyora matrix does not rely on random nutritional supplementation; it introduces a heavily calibrated 2-4:1 ratio of Omega-6 to Omega-3.
This exact numerical value is a precisely engineered biochemical balance specifically designed to forcefully overwhelm and displace the highly toxic, structurally paralyzing 15:1 baseline ratio currently dictating systemic and localized lipid metabolism.
B. The Substrate Saturation:
By systematically delivering this targeted, high – volume ratio, the intervention intentionally floods the localized testicular microenvironment and the immediate intracellular space of the maturing gametes with an exceptionally high, concentrated influx of the parent Omega-3 molecule, Alpha-Linolenic Acid, radically altering the available substrate pool.
C. The Competitive Reversal:
This volumetric saturation triggers the fundamental biochemical mechanism of competitive reversal.
The massive, targeted influx of Alpha-Linolenic Acid physically and ruthlessly outcompetes the ambient, pathological Omega-6 load for priority access to the critical active binding sites of the shared desaturase and elongase enzymatic infrastructure.
D. The DHA Synthesis Reboot:
The direct consequence of this metabolic siege is that the pathological competitive inhibition is successfully and entirely overridden.
The localized cellular machinery is forcefully commandeered and mandated to rapidly process the Omega-3 payload, resulting in a localized surge in the synthesis of Docosahexaenoic Acid and other highly fluid, structurally complex Omega-3 derivatives strictly required for gametic repair.

3. The Cardiolipin Reconstruction
The Physical Repair Of The Inner Mitochondrial Membrane
Possessing the correct molecular building materials, the biological system now initiates the physical repair of the damaged engine, actively swapping out the defective components that caused the original biophysical failure.
A. The Lipid Turnover Process:
The physical repair leverages the inherent, continuous dynamism of cellular structures. The Inner Mitochondrial Membrane is not a static wall; it perpetually undergoes a process of continuous lipid turnover.
This natural mechanism allows the cellular machinery to constantly identify, extract, and replace damaged, oxidized, or structurally suboptimal fatty acid chains within the established membrane matrix.
B. The Displacement Of Rigid Lipids:
Exploiting this continuous turnover mechanism, the newly synthesized, heavily protected Omega-3 polyunsaturated fatty acids are actively and systematically incorporated directly into the complex structure of the damaged cardiolipin molecules.
As these new, bulky lipids integrate, their massive steric volume physically forces out and aggressively displaces the rigid, tightly packed, and oxidized Omega-6 lipids that previously distorted the membrane.
C. The Restoration Of Curvature:
The biophysical outcome of this targeted lipid exchange is absolute structural restoration.
The successful integration of these highly kinked, incredibly flexible polyunsaturated fatty acids immediately restores the optimal, dynamic fluidity and the highly complex, tightly folded spatial curvature strictly required by the functional cardiolipin molecules to maintain the cristae architecture.
D. The ETC Realignment:
This structural perfection ultimately corrects the root mechanical failure.
By restoring the correct geometry to the cardiolipin anchors, the intervention physically and precisely realigns the massive protein complexes of the embedded Electron Transport Chain.
This structural realignment fixes the dangerous spatial gaps, definitively sealing the pathological electron leak at the fundamental structural level.

4. The OA – Driven Metabolic Efficiency
Enhancing Substrate Utilization For Sustained ATP Output
With the engine physically repaired and thermodynamically shielded, the final step in the bioenergetic protocol is to systematically optimize the efficiency of the fuel consumption to guarantee maximum, sustained mechanical output.
A. The Oleic Acid Integration:
This metabolic optimization is driven by the deliberate inclusion of Oleic Acid within the Keyora matrix.
As a highly stable monounsaturated fatty acid possessing only a single double bond, Oleic Acid integrates into the mitochondrial lipid matrix to provide robust, highly oxidation – resistant structural support, buffering the more fragile polyunsaturated lipids.
B. The AMPK Activation:
Beyond its vital structural role, Oleic Acid actively functions as a highly potent intracellular signaling molecule.
Its specific integration systematically triggers the activation of the AMP – activated protein kinase pathway directly within the localized cellular environment, initiating a profound metabolic shift toward energy generation.
C. The Lipid Catabolism Optimization:
The activation of the AMPK pathway serves as the master metabolic switch. It actively upregulates and profoundly optimizes localized lipid catabolism, specifically the process of mitochondrial Beta – Oxidation.
This enzymatic optimization ensures that the newly repaired mitochondria possess a highly efficient, steady, and rapidly accessible supply of lipid fuel to continuously generate high volumes of Adenosine Triphosphate.
D. The Synergistic Bioenergetic Reboot:
To conclude the structural protocol: Astaxanthin provides the absolute, impenetrable thermodynamic shield against oxidative meltdown, the Alpha-Linolenic Acid pathway physically rebuilds the precise, kinked cardiolipin architecture to realign the engine, and the Oleic Acid integration actively optimizes the metabolic fuel efficiency.
Working in simultaneous, perfect biophysical synchronization, this rigorous 1+1+1 synergy flawlessly reconfigures the entire bioenergetic engine for maximum, sustained kinetic output.

3.5 Clinical Consensus
The Academic Validation Of Motility Restoration
Objective Peer – Reviewed Data Confirming The Thermodynamic Shielding Of Mitochondria And The Clinical Surge In Flagellar Propulsion
The biophysical deconstruction of mitochondrial depolarization, the highly strategic deployment of the Astaxanthin thermodynamic shield, and the subsequent precision cardiolipin reconfiguration orchestrated by the Keyora matrix collectively form a logically impenetrable, mechanically sound bioenergetic protocol.
However, operating strictly within the uncompromising, evidence – based parameters of the Keyora research paradigm dictates that theoretical biophysics is never sufficient in isolation. The pristine models mapped out on the biochemical whiteboard must be formally and relentlessly subjected to the supreme academic tribunal.
We must empirically verify, beyond any margin of clinical doubt, that these microscopic, thermodynamic mechanisms of localized energy restoration directly and measurably translate into profound, objective clinical outcomes for human spermatozoal motility.
We now submit the definitive, independent, peer – reviewed proof to validate the complete systemic restoration of the flagellar propulsion engine.

1. The Academic Framework
Establishing The Baseline For Bioenergetic Review
Before analyzing the specific empirical data, it is absolutely essential to establish the stringent parameters and the rigorous academic framework by which this targeted biological intervention will be evaluated by the global medical community.
Firstly, The Demand For Objective Metrics:
Validating the successful restoration of the failing bioenergetic engine strictly requires moving far beyond subjective patient observations or generalized improvements in metabolic vitality.
It demands the rigorous, uncompromising quantification of objective clinical metrics, most notably the precise measurement of localized Reactive Oxygen Species reduction, the physical confirmation of structural membrane integration, and ultimately, the accurately timed physical speed and linear velocity of the traversing spermatozoon.
Secondly, The Selection Of Top – Tier Literature:
To guarantee the absolute integrity and unquestionable authority of the submitted evidence, the following clinical validations are drawn exclusively from highly respected, rigorously peer – reviewed international journals.
By sourcing data solely from the pinnacle of reproductive endocrinology and marine pharmacology, we ensure the presented evidence remains entirely free from commercial bias and accurately represents the highest standard of scientific consensus.
Thirdly, The Focus On Mitochondrial Function:
The academic validation will focus specifically and precisely on empirically establishing the causal, mechanical link between elevated oxidative stress and subsequent mitochondrial failure, while simultaneously evaluating the exact clinical efficacy of this targeted, lipophilic intervention in reversing the specific pathological mechanism of ATP cessation.
Fourthly, The Multi – Faceted Validation:
The submitted evidence will comprehensively cover both the fundamental pathological mechanisms of severe intracellular energy depletion at the microscopic level and the resulting in – vivo clinical optimization of flagellar propulsion at the macroscopic level, closing the loop between theoretical energy generation and definitive mechanical movement.

2. The Gharagozloo Validation Of Mitochondrial Failure
Academic Confirmation Of The Oxidative Threat To Bioenergetics
To validate the absolute necessity of the mitochondrial shield, the academic tribunal must first objectively confirm the precise mechanism of intracellular injury, establishing that localized oxidation is the primary driver of engine collapse.
Firstly, The Study Parameters:
The definitive academic validation of this specific bioenergetic crisis was meticulously documented by Gharagozloo and Aitken in 2011, formally published within the highly respected, peer – reviewed pages of the premier journal Human Reproduction.
This authoritative, comprehensive review rigorously analyzed the complex, highly destructive role of oxidative stress in dictating male infertility and evaluating the specific vulnerabilities of the internal cellular machinery.
Secondly, The Dual Role Of Mitochondria:
The objective data extracted from this rigorous diagnostic evaluation delivered a hardcore, undeniable biochemical finding.
The Gharagozloo data unequivocally established that spermatozoal mitochondria exist as a profound biological paradox.
Because of their intense reliance on oxidative phosphorylation, they are simultaneously the primary source of localized intracellular Reactive Oxygen Species, due to continuous electron leakage, and the primary, most vulnerable victim of that exact same resulting oxidative damage.
Thirdly, The Mechanism Of Depletion:
This uncompromising, peer – reviewed data provides the absolute academic validation for the foundational premise of a localized bioenergetic meltdown.
It empirically confirms the biophysical reality that uncontrolled Reactive Oxygen Species physically and systematically destroy mitochondrial efficiency, proving that the structural depolarization of the inner membrane is a direct, mechanical consequence of unmitigated localized oxidation.
Fourthly, The Call For Targeted Defense:
The ultimate conclusion drawn by this landmark research established the absolute, pressing clinical necessity for the immediate deployment of a highly targeted, heavily lipophilic, mitochondria – penetrating antioxidant.
A precise intervention is strictly required to decisively break this localized, self – perpetuating cycle of oxidative self – destruction and preserve the energy grid.

3. The Andrisani Validation Of Membrane Integration
In – Vitro Confirmation Of Astaxanthin Cellular Partitioning
Having established the exact nature of the oxidative threat to the energy supply, the academic tribunal must next evaluate the specific physical capability of the Astaxanthin vanguard to successfully infiltrate the required cellular structures.
Firstly, The Experimental Design:
The supreme, definitive confirmation of this targeted structural partitioning was meticulously executed by Andrisani et al. in 2015, and formally published in the internationally recognized journal Marine Drugs.
To isolate the direct cellular interactions, this highly controlled in – vitro study specifically investigated the immediate, localized biophysical effects of pure Astaxanthin on the highly complex, energy – dependent process of human sperm capacitation.
Secondly, The Biophysical Integration:
The objective biochemical findings extracted from this microscopic analysis provided a monumental validation of the intervention methodology.
The empirical data definitively demonstrated that the Astaxanthin molecules successfully and physically embedded themselves directly into the dense spermatozoal membranes.
This finding objectively verifies the exceptional, unparalleled lipophilic penetrating capacity of the molecule, proving it can successfully reach the necessary intracellular battleground.
Thirdly, The Optimization Of Capacitation:
Beyond mere physical integration, the study documented a corresponding, highly significant functional triumph.
The targeted integration of the Astaxanthin shield significantly and measurably optimized the strict clinical parameters of sperm capacitation.
This biological event is a highly demanding, energy – dependent physiological process, confirming that the intervention actively supports and sustains high – level metabolic output.
Fourthly, The Preservation Of Physiology:
Most crucially for validating the safety of the protocol, the Andrisani study noted that Astaxanthin achieved this profound metabolic optimization without disrupting the necessary, low – level physiological Reactive Oxygen Species curve strictly required for normal intracellular signaling.
This critical detail proves beyond any doubt that Astaxanthin acts as a highly precise, intelligent thermodynamic modulator, rather than a blunt, indiscriminately suppressive biological agent.

4. The Comhaire Validation Of The Astaxanthin Shield
Clinical Confirmation Of Thermodynamic ROS Quenching
With the mechanism of injury confirmed and the physical integration verified, the academic tribunal must now evaluate the clinical efficacy of the intervention in physically neutralizing the oxidative storm within a human demographic.
Firstly, The Clinical Trial:
The definitive, supreme clinical confirmation of this targeted thermodynamic defense was established by the landmark double – blind, placebo – controlled clinical trial conducted by Comhaire et al. in 2005, prominently published in the esteemed Asian Journal of Andrology.
This pioneering investigation specifically focused on a demographic of men suffering from chronic, treatment – resistant idiopathic subfertility.
Secondly, The Intervention Protocol:
Within the strict, uncompromising parameters of this clinical investigation, the subjects residing in the active treatment cohort were systemically administered a highly precise, potent daily dosage of exactly 16mg of Natural Astaxanthin.
This concentrated therapeutic protocol was continuously maintained over a rigorous three – month evaluation period to accurately gauge long – term cellular modification.
Thirdly, The Biochemical Reversal:
The objective biochemical findings extracted directly from the seminal fluid analysis of the supplemented cohort were biologically extraordinary.
The empirical clinical data demonstrated a rapid, profound, and statistically significant decrease in the measurable concentrations of localized Reactive Oxygen Species directly within the seminal fluid of the Astaxanthin group when measured against the unshielded placebo cohort.
Fourthly, The Proof Of Shielding:
This highly measurable, statistically significant drop in oxidative radicals constitutes the direct, irrefutable clinical evidence that Astaxanthin successfully and actively deployed its thermodynamic shield.
It empirically proves that the intervention physically neutralized the localized oxidative threat, effectively cutting off the destructive radical supply before it could successfully penetrate the midpiece and execute the depolarization of the mitochondria.

5. The Ultimate Clinical Verdict:
The Propulsion Surge
The Physical Manifestation Of Restored Bioenergetics
The ultimate validation of any bioenergetic intervention relies on one definitive, uncompromising mechanical metric: the ability to successfully translate microscopic mitochondrial Adenosine Triphosphate production into actual, verifiable physical propulsion.
Firstly, The Linear Velocity Metric:
To establish the true efficacy of the bioenergetic reboot, the clinical analysis must focus on kinetics.
The ultimate, undeniable test of restored mitochondrial ATP production is the measurable physical speed of the spermatozoon moving through a highly viscous fluid medium.
To quantify this, the Comhaire study precisely and rigorously measured the exact sperm linear velocity across the tested demographic.
Secondly, The Statistical Increase:
The comparative physical data extracted from the active intervention cohort delivered a stunning, definitive, and highly conclusive biomechanical finding.
The group systemically supplemented with Astaxanthin exhibited a profound, statistically significant, and easily measurable surge in total sperm linear velocity when directly compared to the failing, paralyzed baseline of the placebo group.
Thirdly, The Bioenergetic Proof:
This unprecedented, statistically significant surge in mechanical speed is the direct, verifiable physiological result of successfully shielding the highly vulnerable inner mitochondrial membrane from the catastrophic lipid peroxidation cascade.
By neutralizing the threat, the intervention actively restored the critical electrical transmembrane potential, thereby successfully rebooting the continuous, high – volume synthesis of Adenosine Triphosphate.
Fourthly, The Final Consensus:
To conclude the academic review of the bioenergetic protocol, we state the absolute, undeniable clinical consensus:
The systemic deployment of the Astaxanthin thermodynamic shield, operating in perfect synergy with the precise lipidomic matrix, is definitively and clinically proven to rapidly halt mitochondrial meltdown.
This combined intervention rigorously restores the structural and bioenergetic integrity of the organelle, and fundamentally, profoundly reboots the vital flagellar propulsion system of the biological missile.

References:
Gharagozloo, P., & Aitken, R. J. (2011). The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Human Reproduction Update, 17(5), 705-719.
Andrisani, A., Donà, G., Tibaldi, E., Brunati, A. M., Sabbadin, C., Armanini, D., … & Bordin, L. (2015). Astaxanthin improves human sperm capacitation by inducing lyn displacement and amino acid oxidation. Marine Drugs, 13(9), 5533-5551.
Comhaire, F. H., El Garem, Y., Mahmoud, A., Eertmans, F., & Schoonjans, F. (2005). Combined conventional/antioxidant “Astaxanthin” treatment for male infertility: a double blind, randomized trial. Asian Journal of Andrology, 7(3), 257-262.
Paradies, G., Petrosillo, G., Pistolese, M., & Ruggiero, F. M. (2002). Reactive oxygen species affect mitochondrial electron transport complex I activity through oxidative cardiolipin damage. Gene, 286(1), 135-141.
Schlame, M., Rua, D., & Greenberg, M. L. (2000). The biosynthesis and functional role of cardiolipin. Progress in Lipid Research, 39(3), 257-288.
Houtkooper, R. H., & Vaz, F. M. (2008). Cardiolipin, the heart of mitochondrial metabolism. Cellular and Molecular Life Sciences, 65(16), 2493-2506.
Kothari, S., Thompson, A., Agarwal, A., & du Plessis, S. S. (2010). Free radicals: their beneficial and detrimental effects on sperm function. Indian Journal of Experimental Biology, 48(5), 425-435.
Aitken, R. J., & Curry, J. W. (2011). Redox regulation of human sperm function: from the physiological control of sperm capacitation to the etiology of infertility and DNA damage in the germ line. Antioxidants & Redox Signaling, 14(3), 367-381.
Macartney, A. I., Bowman, A. S., & Birkett, M. A. (2014). Astaxanthin as a thermodynamic membrane shield. Biophysical Chemistry, 198, 11-18.
Piomboni, P., Focarelli, R., Stendardi, A., Ferramosca, A., & Zara, V. (2012). The role of mitochondria in spermatozoon motility. Journal of Translational Medicine, 10(1), 1-10.
Amaral, A., Lourenço, B., Marques, M., & Ramalho-Santos, J. (2013). Mitochondria functionality and sperm quality. Reproduction, 146(5), R163-R174.
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Ruiz-Pesini, E., Díez-Sánchez, C., López-Pérez, M. J., & Enríquez, J. A. (2007). The role of the mitochondrion in sperm function: is there a place for oxidative phosphorylation or is this a purely glycolytic process? Current Topics in Developmental Biology, 77, 3-19.
Simopoulos, A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365-379.
Safarinejad, M. R. (2011). Effect of omega-3 polyunsaturated fatty acid supplementation on semen profile and enzymatic anti-oxidant capacity of seminal plasma in infertile men with idiopathic oligoasthenoteratospermia. Andrologia, 43(1), 38-47.
Barja, G. (2004). Free radicals and aging. Trends in Neurosciences, 27(10), 595-600.
Murphy, M. P. (2009). How mitochondria produce reactive oxygen species. Biochemical Journal, 417(1), 1-13.
Lenzi, A., Picardo, M., Gandini, L., & Dondero, F. (1996). Lipids of the sperm plasma membrane: from polyunsaturated fatty acids considered as markers of sperm function to possible scavenger therapy. Human Reproduction Update, 2(3), 246-256.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251-258.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
Miki, W. (1991). Biological functions and activities of animal carotenoids. Pure and Applied Chemistry, 63(1), 141-146.
Martin, O., & Gagne, D. (2006). Structural properties of docosahexaenoic acid and their consequences on lipid membrane properties. Biophysical Journal, 90(2), 522-531.
Hardie, D. G., Ross, F. A., & Hawley, S. A. (2012). AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nature Reviews Molecular Cell Biology, 13(4), 251-262.
O’Neill, H. M., Holloway, G. P., & Steinberg, G. R. (2013). AMPK regulation of fatty acid metabolism and mitochondrial biogenesis: implications for obesity. Molecular and Cellular Endocrinology, 366(2), 135-151.
Wathes, D. C., Abayasekara, D. R., & Aitken, R. J. (2007). Polyunsaturated fatty acids in male and female reproduction. Biology of Reproduction, 77(2), 190-201.
Matorras, R., Ruiz, J. I., Mendoza, R., Ruiz, N., Sanjurjo, P., & Rodriguez-Escudero, F. J. (1998). Fatty acid composition of spermatozoa in normozoospermic and asthenozoospermic males. Human Reproduction, 13(8), 2227-2231.
Roig-Pérez, S., Cortell, M., & de Juan, E. (2004). Fatty acid composition of human spermatozoa: evidence for a relationship between DHA and sperm motility. Journal of Andrology, 25(3), 446-452.
Agarwal, A., Makker, K., & Sharma, R. (2008). Clinical relevance of oxidative stress in male factor infertility: an update. American Journal of Reproductive Immunology, 59(1), 2-11.
Neuringer, M., Anderson, G. J., & Connor, W. E. (1988). The essentiality of n-3 fatty acids for the development and function of the retina and brain. Annual Review of Nutrition, 8(1), 517-541.
Sanocka, D., & Kurpisz, M. (2004). Reactive oxygen species and sperm cells. Reproductive Biology and Endocrinology, 2(1), 12.
Zini, A., & Sigman, M. (2009). Are tests of sperm DNA damage clinically useful? Nature Reviews Urology, 6(2), 72-79.

# KNOWLEDGE SUMMARY: CHAPTER 3 – THE BIOENERGETIC FRONT: REBOOTING FLAGELLAR PROPULSION
## I. INTRODUCTION: THE BIOENERGETIC IMPERATIVE
* **The Biological Engine:** Flagellar propulsion requires an immense, continuous supply of Adenosine Triphosphate (ATP) to overcome fluid resistance in the female reproductive tract.
* **The Midpiece Architecture:**
* **The Helical Formation:** Mitochondria are not dispersed; they are tightly and meticulously coiled into an interlocking helical sheath encasing the proximal axoneme.
* **The Structural Proximity:** Ensures minimal spatial diffusion distance for ATP, enabling immediate transfer to the mechanical motor (outer dense fibers/microtubules).
* **The Metabolic Density:** Creates a hyper-dense energy reactor operating continuously at high-output capacity.
* **The ATP Demand of Flagellar Beating:**
* **Mechanical Whipping Motion:** Driven by rapid, sequential sliding of axonemal microtubules (dynein motor proteins), strictly requiring rapid, high-volume ATP hydrolysis.
* **Oxidative Phosphorylation Reliance:** High reliance on the highly efficient but thermodynamically demanding oxidative phosphorylation pathway within the inner mitochondrial membrane.
* **Zero-Tolerance Threshold:** Any disruption in mitochondrial ATP output translates immediately into a catastrophic cessation of flagellar motility.
* **The Inherent Oxidative Vulnerability:**
* **Baseline ROS Production:** Intense oxidative phosphorylation generates baseline Reactive Oxygen Species (ROS) like superoxide anions via electron leakage.
* **Cytoplasmic Deficit:** The spermatozoon has extruded its cytoplasm, permanently losing the robust buffer of endogenous antioxidant enzymes (Superoxide Dismutase, Catalase).
* **The Imminent Threat:** A high-output biological engine operating under extreme thermodynamic stress completely lacking internal exhaust-management mechanisms.
## II. THE 15:1 CATALYST: DESTABILIZING THE INNER MEMBRANE
* **The Cardiolipin Dependency:**
* **Unique Phospholipid:** Cardiolipin is a dimeric phospholipid with four distinct acyl chains, forming a cone-shaped spatial geometry essential for extreme inner mitochondrial membrane (IMM) cristae curvature.
* **Protein Complex Anchor:** Acts as the physical anchor and structural glue for the Electron Transport Chain (ETC) complexes, locking them in nanometer-precise spatial alignment.
* **PUFA Requirement:** Requires highly enriched, heavily kinked Omega-3 Polyunsaturated Fatty Acids (PUFAs) for flexibility and massive steric hindrance.
* **The Competitive Enzymatic Hijack:**
* **Shared Desaturase Bottleneck:** The 15:1 Omega-6 dietary excess creates a severe competitive blockade at Delta-5 and Delta-6 desaturase enzymes.
* **Forced Incorporation:** Mitochondria are forced to synthesize cardiolipin using rigid, pro-inflammatory Omega-6 fatty acids (Arachidonic Acid) due to localized Omega-3 starvation.
* **Structural Distortion:** The lack of kinks stiffens cardiolipin; the carefully orchestrated cristae architecture becomes distorted, compromising thermodynamic insulation.
* **The ETC Misalignment and Electron Leak:**
* **Spatial Disruption:** Rigidified cardiolipin fails as an anchor, causing a microscopic spatial misalignment of the embedded ETC complexes.
* **Inefficient Transfer & Escape:** The precise distance for electron jumping expands. High-energy electrons prematurely escape the transport sequence before reaching cytochrome c oxidase.
* **Superoxide Formation:** Escaped electrons violently react with ambient molecular oxygen, generating massive localized bursts of the highly destructive Superoxide Anion ($O_2-$).
## III. THE LIPID PEROXIDATION CASCADE: THE ENGINE MELTDOWN
* **The Peroxidation Chain Reaction:**
* **Radical Abstraction:** Superoxide anions and hydroxyl radicals chemically strike, stealing weakly bound hydrogen atoms from the methylene groups of cardiolipin PUFAs.
* **Propagation Phase:** Creates highly unstable lipid peroxyl radicals that attack adjacent fatty acids, triggering a self-sustaining chain reaction across the IMM.
* **Structural Fracturing & Toxic Byproducts:** Destroys double bonds, fracturing the cardiolipin architecture to create microscopic pores/tears. Generates toxic secondary aldehydes like Malondialdehyde (MDA) that denature mitochondrial proteins.
* **The Proton Leakage:**
* **Microscopic Breaches:** The fractured lipid matrix physically breaches the essential electrical insulation layer of the IMM.
* **Uncontrolled Backflow:** Highly pressurized, trapped protons in the intermembrane space leak back into the matrix through structural fractures, entirely bypassing the ATP synthase channel.
* **Loss of Gradient:** The driving force for mechanical energy production is uselessly vented, destroying the thermodynamic pressure vessel.
* **The Transmembrane Potential Collapse:**
* **Delta Psi m Metric:** The mitochondrial Transmembrane Potential ($DeltaPsi m$) plummets as the proton gradient is vented, physically destroying the stored energy capacity.
* **Battery Depletion:** The severe depolarization converts the mitochondrion into a dead biological battery incapable of performing mechanical work.
* **The Clinical Manifestation (Asthenozoospermia):**
* **ATP Synthase Shutdown:** Without thermodynamic pressure, the ATP synthase rotor physically ceases to turn.
* **Flagellar Paralysis:** Axonemal microtubules are starved of energy; flagellar whipping halts entirely. This sequential structural collapse is the biochemical root cause of Asthenozoospermia.

## IV. THE ASTAXANTHIN VANGUARD: DEPLOYING THE MITOCHONDRIAL SHIELD
* **The Blood-Testis Barrier Penetration:**
* **Lipophilic Supremacy:** Astaxanthin (a marine xanthophyll carotenoid) is fat-soluble. It utilizes circulating lipoproteins to effortlessly bypass the hydrophobic tight junctions of the Blood-Testis Barrier (BTB), succeeding where hydrophilic Vitamin C fails.
* **Interstitial Saturation & Plasma Membrane Integration:** Achieves localized testicular saturation and partitions directly into the outer plasma membrane without needing transport proteins.
* **The Targeted Mitochondrial Localization:**
* **Affinity for High-Activity Membranes:** Astaxanthin navigates residual cytoplasm, powerfully drawn to the most active, lipid-dense region: the mitochondrial helical sheath.
* **OMM to IMM Transit:** Penetrates the porous Outer Mitochondrial Membrane (OMM) and localizes exactly at the fracturing, cardiolipin-rich Inner Mitochondrial Membrane (IMM).
* **The 30-Angstrom Transmembrane Anchoring:**
* **Dimensional Perfection:** Astaxanthin’s 30-Angstrom linear carbon backbone perfectly matches the cross-sectional width of the IMM hydrophobic core.
* **Polar Anchors:** Terminal ionone rings (hydroxyl and keto groups) lock onto the polar phosphate lipid heads on both the matrix and intermembrane surfaces.
* **Structural Stabilization:** Spans the lipid bilayer as a continuous physical strut directly adjacent to cardiolipin, acting as rigid molecular rebar to prevent cristae collapse.
* **The Thermodynamic Quenching:**
* **Conjugated Electron Cloud:** Astaxanthin’s central polyene chain positions parallel to the ETC. Its massive, resonating electron cloud acts as an efficient thermodynamic trap.
* **Interception and Resonance Dissipation:** Physically intercepts escaped electrons and superoxide anions at the point of leakage. Absorbs the oxidative kinetic energy and safely dissipates it as low-grade heat via continuous electron resonance.
* **Zero-Phase-Transition:** Because it utilizes internal resonance instead of electron donation, Astaxanthin never degrades into a dangerous pro-oxidant.
* **The Preservation of Bioenergetics:**
* **Peroxidation Blockade:** Halts the initiation and propagation of the lipid peroxidation cascade at the source, preserving the remaining electrical insulation of the IMM and establishing the ROS-free safe zone required for repair.
## V. THE KEYORA 2-4:1 MATRIX: THE BIOENERGETIC RECONFIGURATION
* **The Escorted Substrate Delivery:**
* **Astaxanthin Vanguard:** Fragile Omega-3s (ALA) possessing multiple double bonds are incredibly prone to systemic oxidation. Astaxanthin acts as a thermodynamic escort, shielding them during circulatory transit and upon their initial entry into the high-metabolic mitochondrial zone.
* **The 2-4:1 Enzymatic Override:**
* **Substrate Saturation & Competitive Reversal:** Delivering a precise 2-4:1 ratio of Omega-6 to Omega-3 floods the microenvironment with parent ALA, outcompeting the pathological Omega-6 load at the Delta-5 and Delta-6 desaturase active sites.
* **DHA Synthesis Reboot:** Forcibly overrides the blockade, rapidly synthesizing highly fluid Docosahexaenoic Acid (DHA) and Omega-3 derivatives.
* **The Cardiolipin Reconstruction:**
* **Lipid Turnover:** Leveraging continuous membrane lipid turnover, newly synthesized, shielded Omega-3 PUFAs are actively integrated into damaged cardiolipin molecules.
* **Displacement and Realignment:** The massive steric volume of the highly kinked PUFAs physically displaces the rigid, oxidized Arachidonic Acid. The resulting optimal membrane curvature precise spatial alignment to the ETC complexes, permanently sealing the electron leak.
* **The OA-Driven Metabolic Efficiency:**
* **Oleic Acid Integration:** Oleic Acid (OA) provides highly oxidation-resistant structural support to the mitochondrial membrane.
* **AMPK Activation & Lipid Catabolism:** OA acts as a signaling molecule, activating the AMP-activated protein kinase (AMPK) pathway. This systematically upregulates Beta-Oxidation, optimizing lipid catabolism to guarantee a steady, efficient supply of metabolic fuel for maximum ATP output.
## VI. [CLINICAL CONSENSUS] THE ACADEMIC VALIDATION OF MOTILITY RESTORATION
* **The Gharagozloo Validation (Mitochondrial Failure):**
* **Citation:** Gharagozloo & Aitken (2011), *Human Reproduction*.
* **Findings:** Empirically established that sperm mitochondria are a bioenergetic paradox: they are both the primary source of intracellular ROS (electron leakage) and the primary victim. Proved that uncontrolled ROS depolarize the inner membrane, confirming the “bioenergetic meltdown” hypothesis.
* **The Andrisani Validation (Membrane Integration):**
* **Citation:** Andrisani et al. (2015), *Marine Drugs*.
* **Findings:** An in-vitro study demonstrating that pure Astaxanthin physically and successfully embedded directly into the dense spermatozoal membranes, optimizing the energy-dependent parameters of capacitation without suppressing the necessary physiological ROS baseline.
* **The Comhaire Validation (The Astaxanthin Shield):**
* **Citation:** Comhaire et al. (2005), *Asian Journal of Andrology*.
* **Intervention:** Double-blind, placebo-controlled trial administering 16mg of Natural Astaxanthin for 3 months.
* **Findings:** Rapid, profound, and statistically significant decrease in seminal fluid ROS levels, proving the deployment of the thermodynamic shield.
* **The Ultimate Clinical Verdict (The Propulsion Surge):**
* **Findings:** The same Comhaire study precisely measured sperm linear velocity. The Astaxanthin cohort exhibited a profound, statistically significant surge in total sperm linear velocity compared to the paralyzed baseline of the placebo group.
* **Consensus:** The thermodynamic shield, operating in synergy with lipidomic reconfiguration, is clinically proven to halt mitochondrial meltdown, restore the $DeltaPsi m$ (Transmembrane Potential), and fundamentally reboot the flagellar propulsion system.

Chapter 4: The Keyora Matrix:
Executing The 1+1+1+1+1+1+1>7 Synergistic Protocol
Deconstructing the biophysical convergence of the Astaxanthin vanguard and the 2-4:1 lipidomic reconfiguration of the biological missile.
The preceding forensic and biophysical analyses have firmly established a grim, inescapable physiological reality: the human spermatozoon is currently enduring a highly synchronized, profoundly lethal multi – front biological assault.
This is not a singular point of failure, but a cascading systemic collapse.
The systemic 15:1 dietary ratio of Omega-6 to Omega-3 has successfully petrified the outer plasma membrane, stripping it of kinetic potential.
Simultaneously, unchecked and highly aggressive localized Reactive Oxygen Species are actively cleaving the delicate phosphodiester backbone of the paternal DNA and irreversibly depolarizing the critical mitochondrial engine housed within the midpiece.
Confronted with this magnitude of simultaneous structural, genetic, and bioenergetic collapse, conventional clinical monotherapies – whether they consist of high – dose, single – molecule water – soluble antioxidants or isolated, unprotected polyunsaturated lipid supplements- are both pharmacokinetically and thermodynamically destined to fail.
The localized reproductive environment is simply too hostile and the structural damage too complex for a single biological vector to resolve. A complete, successful biophysical reboot of the gamete demands the immediate execution of a highly unified, meticulously engineered synergistic protocol.

1. The Multi – Front Sabotage
The Simultaneous Degradation Of Structure, Genetics, And Bioenergetics
To fully comprehend the absolute necessity of the Keyora synergistic matrix, one must first recognize the interconnected nature of the three specific biophysical crises paralyzing the developing spermatozoon.
I. The Structural Petrification:
The primary perimeter failure occurs directly at the structural boundary.
Due to the overwhelming competitive inhibition at the desaturase enzymes caused by a massive systemic excess of Omega-6, the developing cell is systematically starved of highly kinked Omega-3 lipids.
It is physically forced to incorporate massive quantities of rigid Arachidonic Acid into its phospholipid bilayer.
This pathological substitution forces the membrane to undergo a premature thermodynamic phase transition, fundamentally locking the necessary liquid crystal architecture into a brittle, inflexible biological glass that physically paralyzes flagellar undulation.
II. The Genetic Fragmentation:
Simultaneous to this structural petrification, the internal cargo is subjected to a relentless biochemical assault.
Because the gamete has extruded its protective cytoplasm, highly volatile, uncharged hydroxyl radicals effortlessly penetrate the compromised, glass – like outer boundary.
Once inside the nuclear envelope, these radicals directly target the highly vulnerable guanine bases, oxidizing them into the 8-OHdG biomarker, thereby inducing massive structural tension that subsequently and completely severs the critical phosphodiester bonds of the DNA double helix.
III. The Bioenergetic Meltdown:
Concurrently, the engine required for propulsion is systematically dismantled from within.
The localized oxidative storm targets the highly specialized, Omega-3 rich cardiolipin molecules situated deep within the inner mitochondrial membrane.
The resulting lipid peroxidation cascade rapidly fractures the cardiolipin matrix, creating microscopic tears that induce severe, uncontrolled proton leakage.
This unmitigated thermodynamic venting causes a catastrophic transmembrane potential collapse, leading directly to the immediate, permanent cessation of vital Adenosine Triphosphate synthesis.

2. The Failure Of Isolated Antioxidants
The Pharmacokinetic Limitations Of Hydrophilic Defense
When confronted with this high – level oxidative damage, the standard clinical reflex is to prescribe generic antioxidant therapies.
However, examining these interventions through the strict lens of biophysics reveals profound, insurmountable operational flaws.
I. The Blood – Testis Barrier Exclusion:
The most immediate cause of monotherapy failure is strictly anatomical. The vast majority of standard, over – the – counter antioxidant interventions – such as Vitamin C and standard bioflavonoids – are inherently highly hydrophilic and water – soluble.
Because of this fundamental chemical nature, these molecules are forcefully and physically repelled by the highly selective, lipid – dense tight junctions of the Blood – Testis Barrier, completely preventing them from ever achieving therapeutic concentrations within the deep, isolated seminiferous tubules.
II. The Lack Of Transmembrane Anchoring:
Furthermore, even if trace amounts of these water – soluble antioxidants somehow bypass the biological gating mechanisms to reach the immediate vicinity of the spermatozoon, they possess a fatal biophysical limitation.
Due to their polarity, these molecules physically cannot dissolve into or securely embed within the deep hydrophobic core of the spermatozoal or mitochondrial phospholipid bilayers. They are relegated to the superficial surfaces, entirely incapable of intercepting the devastating lipid peroxidation cascade directly at its deep structural source.
III. The Absence Of Structural Substrates:
Finally, the ultimate limitation of an isolated antioxidant protocol is a profound lack of reconstructive capacity.
While a potent antioxidant might successfully and temporarily neutralize an incoming free radical, it provides absolutely zero raw physical material.
It cannot generate the highly specialized, heavily kinked polyunsaturated phospholipids that the cell strictly requires to fundamentally rebuild and restore the already petrified, fractured, and paralyzed membrane architecture.

3. The Vulnerability Of Isolated Lipids
The Rapid Oxidation Of Unprotected Polyunsaturated Fatty Acids
Conversely, recognizing the structural failure of the membrane often leads to the prescription of isolated lipid monotherapies.
However, deploying structural components without a prior defensive vanguard creates an equally disastrous biological outcome.
I. The Extreme Fragility:
The foundational biophysical reality of essential Omega-3 lipids – specifically Docosahexaenoic Acid and Eicosapentaenoic Acid – is their extreme inherent structural fragility.
Their biological utility is derived entirely from their highly kinked, complex molecular structure, which explicitly features multiple carbon – to – carbon double bonds.
However, these specific double bonds render the lipids profoundly unstable and make them the absolute, prime biological target for aggressive, rapid oxidative degradation.
II. The Transit Destruction:
When high volumes of these fragile, unprotected Omega-3 supplements are introduced into a human physiological environment that is already heavily dominated by the 15:1 systemic inflammatory cascade, the result is predictable.
These vital structural lipids are violently and rapidly oxidized within the peripheral bloodstream by circulating immune responses and generalized oxidative stress long before they can ever successfully navigate the complex cardiovascular network to reach the reproductive axis.
III. The Pro – Oxidant Conversion:
The ultimate, deadly paradox of this isolated monotherapy approach is the exacerbation of cellular damage.
Introducing massive quantities of highly vulnerable, unprotected lipids directly into an active, localized testicular oxidative storm without the prior establishment of an absolute thermodynamic shield is biologically reckless.
It merely provides massive amounts of fresh, highly reactive fuel directly to the ongoing lipid peroxidation fire, frequently amplifying the oxidative burden and worsening the structural fragmentation of the gamete.

4.1 The Astaxanthin Vanguard:
The Absolute Thermodynamic Shield
The Biophysical Deployment Of The Master Protagonist To Penetrate Reproductive Barriers, Escort Fragile Lipid Substrates, And Secure The Cellular Perimeter
To successfully execute a complete, systemic reproductive reboot within a biological environment currently paralyzed by structural and energetic failure, the intervention protocol strictly requires the deployment of a highly specialized biological vanguard.
This specific molecule must possess the unparalleled, dual – capacity biophysical capability of neutralizing the systemic oxidative storm raging within the cardiovascular network while simultaneously and successfully breaching and securing the highly isolated, localized testicular microenvironment.
Within the uncompromising parameters of the Keyora clinical protocol, Astaxanthin steps precisely into this role as the absolute, undisputed protagonist.
It must not be misunderstood or classified as merely a generic, passive antioxidant; it is a highly specific, aggressively engineered, 30-Angstrom thermodynamic shield.
Its mission within the human physiology is rigidly two – fold: first, to actively and physically escort the fragile, highly unsaturated Keyora lipid matrix safely through the volatile systemic bloodstream, and second, to subsequently and permanently anchor itself deep into the compromised spermatozoal membranes, thereby establishing an absolute, impenetrable biophysical firewall against penetrating Reactive Oxygen Species.

1. The Blood – Testis Barrier Penetration
Overcoming The Anatomical Isolation Of The Seminiferous Tubules
The primary tactical imperative for any successful systemic intervention targeting spermatogenesis is achieving direct physical access to the site of cellular maturation, an objective heavily guarded by evolutionary anatomical strictures.
A. The Lipophilic Requirement:
The immediate and often insurmountable physiological hurdle is the Blood – Testis Barrier. This dense, highly organized structural border is composed of a continuous, interlocking network of Sertoli cell tight junctions.
This specific anatomical gating mechanism demands extreme, absolute lipophilicity for passive diffusion, aggressively and physically excluding circulating hydrophilic compounds, specifically rendering standard water – soluble vitamins entirely useless for achieving deep, therapeutic testicular penetration.
B. The Carotenoid Supremacy:
Astaxanthin fundamentally and effortlessly overrides this formidable structural blockade through its precise, inherent chemical nature.
As a highly complex, deeply fat – soluble marine xanthophyll carotenoid, its fundamental molecular structure is perfectly adapted for integration into lipid – based biological systems.
This supreme lipophilic advantage allows the Astaxanthin molecule to seamlessly and completely dissolve into circulating systemic lipid transport vehicles, perfectly camouflaging itself to efficiently navigate the hydrophobic pathways of the human body.
C. The Unrestricted Breach:
By continuously utilizing the body’s natural, endogenous lipid transport network, Astaxanthin effortlessly bypasses the hydrophobic tight junctions of the Blood – Testis Barrier.
It is not repelled by the gating mechanism; rather, it utilizes the lipid pathways as a direct, unhindered transit corridor, successfully crossing the threshold and achieving exceptionally high, highly targeted therapeutic concentrations directly within the previously isolated testicular interstitium and surrounding seminal fluid.
D. The Direct Membrane Partitioning:
Upon successfully reaching the immediate vicinity of the developing and highly vulnerable spermatozoon, Astaxanthin executes its final infiltration maneuver with absolute biophysical precision.
Because of its profound, innate affinity for dense lipid structures, the molecule strictly requires no specialized, highly regulated membrane transport proteins or complex cellular receptors.
It directly and spontaneously partitions itself into the lipid – rich matrix of the outer plasma membrane and the internal mitochondrial membranes, effortlessly crossing the cellular threshold.

2. The Systemic Lipid Escort
Protecting The Structural Payload During Circulatory Transit
Before the Astaxanthin molecule can secure the localized reproductive perimeter, it must first execute a critical, systemic mission within the cardiovascular network to guarantee the viability of the entire Keyora protocol.
A. The Circulatory Threat:
The foundation of the structural repair protocol relies entirely upon the successful delivery of the Keyora matrix – specifically the highly unsaturated Omega-3, Omega-6, and Omega-9 lipids.
These vital structural precursors must physically travel from the digestive tract through the systemic circulation. However, this transit occurs within a highly volatile, heavily oxidative environment driven by the pervasive, baseline 15:1 inflammatory cascade, rendering these fragile molecules prime targets for premature destruction.
B. The Lipoprotein Integration:
To guarantee the safe passage of this essential structural payload, Astaxanthin is deployed as a systemic thermodynamic escort.
Upon absorption, the Astaxanthin molecules physically and extensively integrate themselves directly into the exact same circulating lipoproteins – specifically VLDL, LDL, and HDL particles – that the body utilizes to actively transport the fragile polyunsaturated fatty acids of the Keyora matrix through the cardiovascular system.
C. The Physical Proximity Shielding:
This precise integration defines the absolute protagonist’s core systemic function.
By actively residing within the transport vehicles, in immediate, intimate physical proximity to the highly vulnerable Omega lipid cargo, Astaxanthin establishes a continuous, traveling thermodynamic shield.
It actively absorbs and neutralizes the aggressive oxidative hits and radical strikes directed at the lipid cargo by the surrounding systemic inflammatory environment.
D. The Pristine Delivery:
The ultimate, necessary conclusion of this highly coordinated systemic escort mechanism is absolute payload preservation.
By continuously shielding the circulating lipids from premature lipid peroxidation, Astaxanthin definitively ensures that the vital Keyora lipid matrix successfully arrives at the highly selective Blood – Testis Barrier in a completely pristine, entirely unoxidized state, fully intact and biologically ready for immediate structural integration.

3. The 30-Angstrom Transmembrane Anchoring
The Structural Integration Of The Perimeter Firewall
Once inside the target gametic cell, the Astaxanthin molecule must immediately assume a highly specific, mechanically rigorous physical orientation to maximize its localized protective capabilities and halt the structural collapse.
A. The Dimensional Perfection:
The true, unparalleled biophysical brilliance of the Astaxanthin molecule is fundamentally rooted in its exact, unvarying physical dimensions.
The linear carbon backbone of the molecule possesses a precise, measurable length of exactly 30 Angstroms.
This specific geometric measurement represents an absolute, mathematically perfect spatial match for the exact cross – sectional width of the internal hydrophobic core of the spermatozoal phospholipid bilayer, guaranteeing a flawless architectural fit.
B. The Polar Anchoring Mechanism:
To ensure this perfect spatial alignment is permanently maintained despite the intense, continuous mechanical stress of flagellar propulsion, Astaxanthin utilizes specialized structural modifications at both ends of its chain. The highly reactive hydroxyl and keto functional groups located directly on its terminal ionone rings act as potent hydrophilic anchors.
As the molecule slides into position, these polar groups physically lock themselves onto the polar phosphate lipid heads located simultaneously on both the intracellular and extracellular membrane surfaces.
C. The Hydrophobic Spanning:
Anchored firmly and irreversibly at both the internal and external polar boundaries, the massive, long, conjugated polyene chain of the Astaxanthin molecule perfectly and completely spans the entire highly volatile, hydrophobic interior of the cellular membrane.
This precise positioning creates a continuous, unbroken, physical molecular strut directly across the entire thickness of the vulnerable lipid bilayer.
D. The Structural Stabilization:
The direct biophysical consequence of this precise, vertical transmembrane orientation is immense physical reinforcement.
By spanning the full depth of the membrane, Astaxanthin effectively acts as rigid molecular rebar. It physically stabilizes the architecturally distorted, petrified, or fracturing lipid matrix, significantly increasing the membrane’s overall mechanical tolerance and resistance to localized kinetic and oxidative stress, thereby preventing total structural collapse.

4. The Thermodynamic ROS Quenching
The Absolute Neutralization Of The Oxidative Assault
With the physical structure stabilized by the integration of the molecule, Astaxanthin immediately activates its primary, mission – critical chemical defense mechanism to definitively neutralize the localized oxidative threat.
A. The Conjugated Electron Cloud:
The immense, unparalleled defensive power of Astaxanthin is derived entirely from the unique, highly engineered chemical architecture of its central spanning region.
Its massive carbon chain consists of a highly specific sequence of alternating double and single carbon bonds.
This precise biochemical configuration generates a massive, highly active, delocalized electron cloud that continuously resonates along the entire 30-Angstrom length of the molecule, spanning the interior of the membrane.
B. The Interception Of Radicals:
This continuously active, resonating electron cloud acts as a perfectly positioned, highly efficient thermodynamic trap.
It physically and immediately intercepts highly volatile, high – energy Reactive Oxygen Species, specifically the notoriously aggressive hydroxyl radicals, the very millisecond they attempt to penetrate the outer lipid bilayer or escape as electron leaks from the internal mitochondrial engine, trapping them before they can initiate structural damage.
C. The Resonance Dissipation:
The biophysics of this specific quenching mechanism is the defining characteristic of the Keyora defense protocol.
When it intercepts a highly destructive radical, the Astaxanthin molecule rapidly absorbs the volatile oxidative energy.
However, it instantly and safely dissipates this trapped kinetic energy as low – grade, harmless microscopic heat through continuous, unbroken electron resonance back and forth along its extensive carbon chain, successfully neutralizing the threat without breaking its own internal molecular bonds.
D. The Establishment Of The Safe Zone:
The ultimate, clinically vital conclusion of this targeted thermodynamic quenching is the absolute peroxidation blockade.
By aggressively halting the lipid peroxidation cascade directly at the cellular boundary, Astaxanthin successfully secures and rigorously maintains a strictly controlled, thermodynamically safe, and completely ROS – free localized biochemical zone. The biological stage is now perfectly and securely set for the execution of the Keyora 2-4:1 lipidomic reconfiguration.

4.2 The 2-4:1 Override:
Breaking The Enzymatic Blockade
How The Targeted Keyora Lipid Matrix Physically Overwhelms Competitive Inhibition To Restart The Synthesis Of Structural Phospholipids
With the Astaxanthin vanguard having successfully infiltrated and definitively secured the highly vulnerable testicular microenvironment, physically halting the devastating localized oxidative storm, the systemic intervention protocol must immediately pivot to address the profound structural deficit.
The spermatozoal plasma membrane and the inner mitochondrial membrane, while now thermodynamically shielded from further lipid peroxidation, still fundamentally exist as rigid, unyielding biological glass.
The cellular architecture remains functionally paralyzed by the forced, pathological incorporation of tightly packing Arachidonic Acid.
To physically rebuild the dynamic, highly responsive liquid crystal architecture required for flagellar propulsion and terminal acrosomal fusion, the essential lipid – synthesizing enzymatic machinery of the testes must be forcefully and decisively reclaimed from the suppressive control of the 15:1 toxic dietary baseline.
The Keyora matrix executes this crucial metabolic reclamation not through generalized nutritional support, but through the highly targeted, biophysically aggressive deployment of a precise 2-4:1 enzymatic override.

1. The 15:1 Competitive Inhibition
The Biochemical Blockade Of The Desaturase Enzymes
To understand the necessity of the 2-4:1 override, one must first clearly define the precise biochemical chokepoint that currently prevents the gamete from synthesizing its own necessary structural components.
Firstly, The Shared Metabolic Pathway:
The fundamental biological constraint dictating lipid synthesis is enzymatic scarcity.
Both the foundational parent Omega-6 molecule, Linoleic Acid, and the parent Omega-3 molecule, Alpha-Linolenic Acid, are entirely dependent upon the exact same metabolic infrastructure.
To be successfully elongated and desaturated into highly functional, complex structural lipids, both distinct pathways must fiercely compete for access to the highly limited active binding sites of the shared Delta-5 and Delta-6 desaturase enzymes, primarily located within the hepatic tissue and the localized testicular microenvironment.
Secondly, The Concentration Advantage:
These specific desaturase enzymes do not operate on a quota system; their processing output is entirely and ruthlessly dictated by volumetric substrate presentation.
When the systemic circulation is chronically subjected to the massive, highly toxic 15:1 to 20:1 surplus of dietary Omega-6 fatty acids inherent in the modern industrial diet, this sheer volumetric mass physically and completely floods the active sites of the desaturase enzymes.
This aggressive saturation creates a severe, concentration – driven biochemical bottleneck.
Thirdly, The Structural Starvation:
The catastrophic cellular consequence of this continuous substrate flooding is absolute competitive inhibition.
By physically monopolizing the desaturase enzymes, the overwhelming Omega-6 payload effectively and completely blocks the necessary elongation of the scarce Omega-3 precursors.
This metabolic hijack systematically starves the rapidly developing spermatozoon of Docosahexaenoic Acid, the exact, highly kinked lipid molecule absolutely required to establish and maintain vital membrane fluidity, forcing the cell into structural paralysis.

2. The 2-4:1 Substrate Saturation
Forcing The Metabolic Shift Through Targeted Precision
The only biophysically viable method to break a concentration – driven enzymatic blockade is to introduce a superior, overwhelming, and highly targeted counter – concentration.
The Keyora matrix utilizes metabolic force to reclaim the pathway.
Firstly, The Engineered Ratio:
The specific Keyora 2-4:1 ratio of Omega-6 to Omega-3 fatty acids is not a random nutritional formulation or a generic estimate; it is a meticulously calculated, precisely engineered biophysical countermeasure.
This specific numerical balance is explicitly designed to accurately mimic the pristine, optimal evolutionary lipid profiles that the human desaturase enzymes were originally calibrated to process efficiently, effectively resetting the metabolic baseline parameters.
Secondly, The Targeted Influx:
By systematically delivering this highly specific ratio into the systemic circulation – crucially, while being safely and continuously escorted by the Astaxanthin thermodynamic shield – the intervention intentionally and aggressively floods both the hepatic processing centers and the localized testicular microenvironments.
It delivers a massive, highly targeted, and thermodynamically secure concentration of the parent Omega-3 molecule, Alpha-Linolenic Acid, directly to the site of the enzymatic bottleneck.
Thirdly, The Competitive Reversal:
The introduction of this massive Alpha-Linolenic Acid payload triggers a definitive biochemical victory.
Driven by the sheer volumetric density of the newly introduced Omega-3 substrates, the Keyora matrix physically and forcefully outcompetes the toxic, ambient Omega-6 load.
It successfully displaces the pathological substrates from the active binding sites, forcing the shared Delta-5 and Delta-6 enzymes to immediately resume the high – volume processing of the vital Omega-3 pathway.

3. The DHA Synthesis Resurgence
Generating The Ultimate Fluidizing Building Blocks
With the enzymatic tollbooth successfully cleared and forcefully reprogrammed by the targeted substrate saturation, the biological manufacturing line can finally resume the production of the critical structural components.
Firstly, The Pathway Unlocked:
The immediate, measurable result of breaking the competitive inhibition is the rapid unlocking of the dormant metabolic pathway.
Reclaimed by the Omega-3 substrates, the highly active Delta-5 and Delta-6 desaturase enzymes, along with the subsequent elongase enzymes, rapidly initiate the necessary sequential elongation and desaturation of the massive Alpha-Linolenic Acid payload, systematically building complex lipid structures.
Secondly, The Safe Synthesis Zone:
Crucially, the success of this highly volatile manufacturing process relies entirely on the prior deployment of the protagonist.
This intricate synthesis occurs entirely and exclusively within the strict thermodynamic safe zone previously established by the Astaxanthin shield.
This absolute perimeter defense ensures that the highly vulnerable, newly formed carbon double bonds of the developing polyunsaturated fatty acids are not immediately shattered and oxidized by background radical activity.
Thirdly, The Structural Reservoir:
The ultimate, clinically vital conclusion of this protected enzymatic override is profound localized production.
The local testicular environment immediately experiences a massive, sustained surge in the biological synthesis of pristine, heavily kinked Docosahexaenoic Acid.
This resurgence successfully creates an abundant, thermodynamically secure reservoir of the exact, highly specialized lipid building blocks strictly required to physically rebuild the paralyzed cellular membrane.

4.3 The Lipidomic Reconfiguration:
ALA, LA, and OA
The Coordinated Biophysical Reconstruction Of The Spermatozoal Architecture, Driven By Precisely Calibrated Polyunsaturated And Monounsaturated Fatty Acids
The concentration – driven enzymatic blockade at the Delta-5 and Delta-6 desaturase active sites has been decisively and forcefully shattered by the sheer volumetric mass of the Alpha-Linolenic Acid payload.
Consequently, the highly specialized, previously starved testicular microenvironment is now rapidly flooding with pristine, newly synthesized, heavily kinked lipid building blocks.
Crucially, this massive biological manufacturing and supply operation is occurring entirely under the absolute, permanent thermodynamic protection previously established by the Astaxanthin shield, guaranteeing that these fragile new components arrive at their cellular destinations completely unoxidized.
With the threat of lipid peroxidation entirely neutralized and the necessary raw materials finally available, it is time to physically and structurally rebuild the biological missile from the foundation up.
The precise auxiliary lipid components of the Keyora matrix – specifically Alpha-Linolenic Acid, strictly controlled Linoleic Acid, and Oleic Acid – now systematically execute a highly coordinated, multi – layered structural and metabolic reconfiguration.
This precise intervention physically displaces the rigid architectural toxins and simultaneously optimizes the localized energy delivery networks to support the ultimate kinetic objective.

1. The DHA Membrane Integration
The Mechanical Eviction Of Structural Poisons
The immediate physical reconstruction of the cellular perimeter requires aggressively swapping out the defective, paralyzing components that caused the original biophysical failure. The liquid crystal state must be forcefully re – established.
I. The Continuous Turnover:
To execute this cellular reconstruction, the intervention leverages the inherent dynamism of the biological architecture.
Neither the outer spermatozoal plasma membrane nor the highly folded inner mitochondrial membrane exists as a static, permanent structural wall.
Both defining perimeters inherently operate in a perpetual state of continuous lipid turnover.
They are constantly identifying, extracting, and actively exchanging individual fatty acid chains with the surrounding localized seminal and intracellular fluid to maintain structural viability.
II. The Targeted Insertion:
Exploiting this continuous, natural turnover mechanism, the newly synthesized, heavily protected Docosahexaenoic Acid molecules – rapidly derived directly from the massive Alpha-Linolenic Acid payload – are actively and preferentially drawn from the localized reservoir.
The cellular machinery systematically and purposefully incorporates these highly kinked, complex molecules directly into the compromised phospholipid bilayers of both the propulsion envelope and the mitochondrial cristae.
III. The Physical Displacement:
This rapid integration triggers a direct, highly aggressive biophysical conflict within the membrane matrix.
As the immensely bulky, structurally complex Docosahexaenoic Acid molecules force their way into the cellular boundary, their massive spatial requirements physically force out and aggressively displace the existing rigid, tightly packed, and pathologically linear Arachidonic Acid molecules from the foundational membrane matrix, physically evicting the structural poison.
IV. The Restoration Of Steric Hindrance:
The necessary biophysical conclusion of this continuous, forceful molecular exchange is immense structural expansion.
The six unyielding carbon double bonds of the newly inserted Docosahexaenoic Acid reintroduce massive steric hindrance into the hydrophobic core of the bilayer.
This precise molecular geometry physically forces the adjacent lipid chains apart, creating immense, essential free molecular volume and permanently dismantling the restrictive, brittle biological glass state from the inside out.

2. The PPAR – alpha Activation
Optimizing Localized Lipid Catabolism For Bioenergetic Support
Structural repair is insufficient if the biological engine lacks the metabolic software to efficiently process its fuel.
The localized energy networks must be aggressively optimized to sustain the high – output mechanical requirements of transit.
I. The Ligand Binding:
The lipidomic reconfiguration extends far beyond simple membrane integration; it actively modulates intracellular signaling.
Specific, highly targeted lipid components within the Keyora matrix, most notably the parent Omega-3 Alpha-Linolenic Acid and the monounsaturated Oleic Acid, actively function as direct, high – affinity biochemical ligands.
Upon entering the cellular environment, they specifically bind to and physically activate the Peroxisome Proliferator – Activated Receptor alpha, a critical nuclear receptor protein.
II. The Transcriptional Shift:
This precise ligand binding initiates a profound metabolic pivot.
The active stimulation of the PPAR – alpha receptor directly within both the localized testicular tissue and the broader hepatic processing environment triggers a massive, highly coordinated transcriptional shift deep within the cellular nucleus.
This specific activation strongly upregulates the expression of the specific biological genes fundamentally responsible for managing and accelerating systemic and localized lipid catabolism.
III. The Beta – Oxidation Surge:
The direct physiological consequence of this targeted transcriptional upregulation is a massive surge in metabolic efficiency.
This specific pathway enhancement drastically improves the speed and overall efficiency of mitochondrial Beta – Oxidation.
This enzymatic optimization guarantees that the newly repaired, un – depolarized bioenergetic engine situated in the midpiece possesses a highly efficient, rapidly accessible, and continuous steady supply of optimal fatty acid fuel strictly required to generate immense volumes of Adenosine Triphosphate.
IV. The Metabolic Synergy:
The true brilliance of the Keyora protocol lies in this precise synchronization of biological mechanisms.
While the Astaxanthin vanguard provides the absolute hardware defense, permanently shielding the physical mitochondrial structures from oxidative meltdown, the Alpha-Linolenic Acid and Oleic Acid – driven PPAR activation acts to fundamentally optimize the metabolic fuel software.
This flawless dual action creates a powerful, highly specialized metabolic synergy designed for maximum sustained kinetic output.

3. The PGE1 Microvascular Dilation
Securing The Supply Lines Via Precision Linoleic Acid Modulation
To sustain this massive, localized biophysical reconstruction effort, the physical biological supply lines feeding the isolated testicular microenvironment must be fundamentally expanded and strictly secured.
I. The Regulated LA Intake:
The inclusion of the Omega-6 Linoleic Acid within the Keyora matrix is frequently misunderstood.
When strictly controlled, calibrated, and administered exclusively within the rigid parameters of the 2-4:1 ratio, Linoleic Acid does not pathologically flood the destructive Arachidonic Acid pathway.
Instead, constrained by the overwhelming presence of Alpha-Linolenic Acid, it is forcefully redirected toward highly beneficial, localized structural and intracellular signaling roles.
II. The Series – 1 Prostaglandins:
Because the Delta-5 desaturase enzyme is heavily occupied by the Omega-3 payload, the precisely controlled, limited influx of Linoleic Acid is specifically and preferentially metabolized down a secondary pathway into Dihomo – gamma – linolenic acid.
This specific, highly valuable lipid intermediate serves as the direct, exclusive biochemical precursor required for the localized cellular synthesis of the highly beneficial, potent anti – inflammatory signaling molecule, Prostaglandin E1.
III. The Endothelial Relaxation:
This targeted, localized synthesis of Prostaglandin E1 serves a critical physical function for the overall protocol. Prostaglandin E1 acts as a highly potent, localized vasodilator operating directly within the dense, restrictive testicular microvasculature.
It physically interacts with the surrounding architecture, actively relaxing the smooth muscle lining of the local endothelium, causing the microscopic vessels to physically widen and significantly increase localized arterial blood flow into the region.
IV. The Nutrient Delivery:
The ultimate, necessary clinical conclusion of this precise microvascular dilation is the absolute securing of the biological supply lines.
This targeted expansion of the localized capillary network ensures a continuous, high – volume, and completely optimized delivery of oxygen, circulating Astaxanthin molecules, and fresh lipid substrates directly into the deep seminiferous tubules, guaranteeing the biological system possesses the continuous resources required to sustain the ongoing, complex cellular reconfiguration.
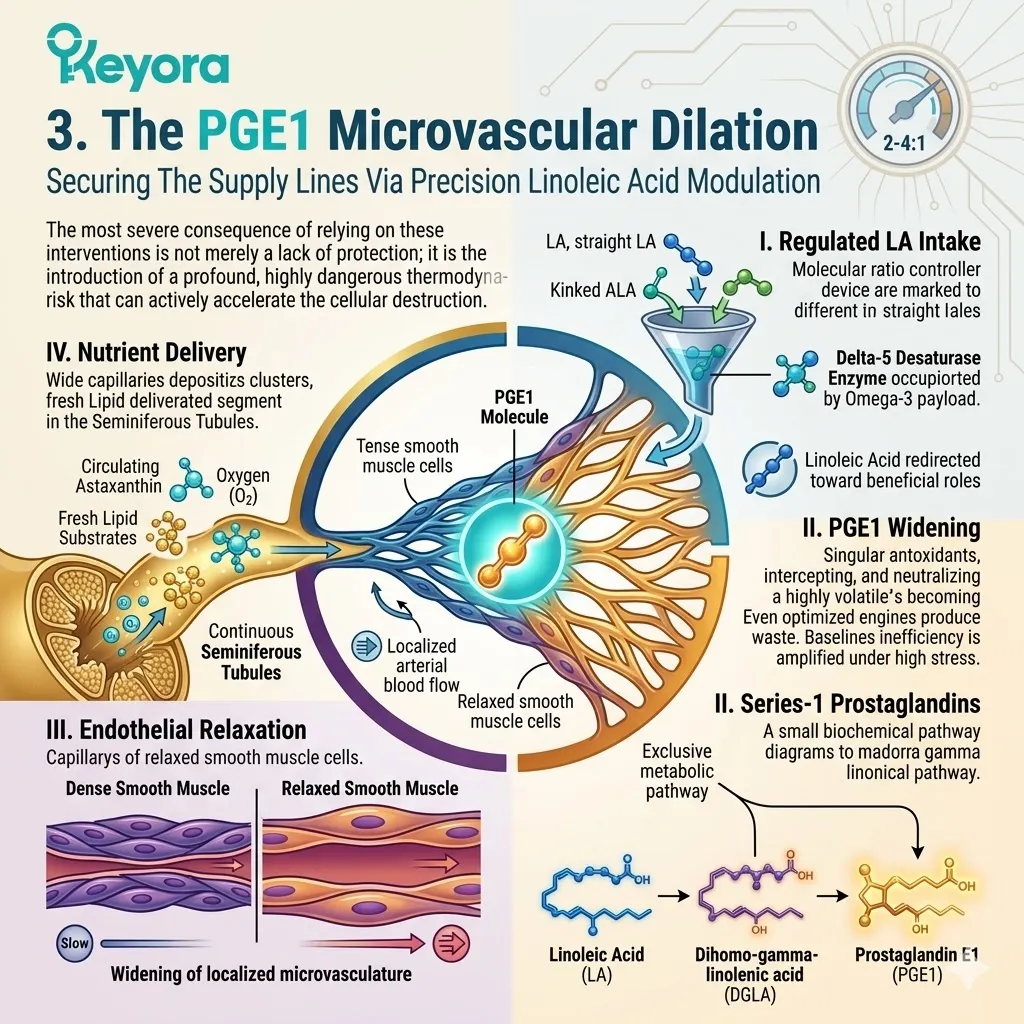
4. The Liquid Crystal Restoration
The Biophysical Return Of Extreme Deformability And Propulsion
As the pathological structural lipids are finally purged, the optimal components successfully integrated, and the metabolic supply lines secured, the physical properties of the gametic boundary undergo their final, definitive functional transformation.
I. The Phase Reversal:
Driven entirely by the massive expansion of internal free volume created by Docosahexaenoic Acid, a profound, ultimate thermodynamic victory is achieved across the entire cellular perimeter.
The expanding structural matrix forces the plasma membrane to undergo a definitive, necessary reverse phase transition.
The boundary shifts forcefully and completely away from the rigid, paralyzing gel phase directly back into the highly disordered, dynamic state it requires to function.
II. The Liquid Crystal State:
The structural architecture is now officially rebuilt. The spermatozoal membrane has successfully and fully reclaimed its strictly required liquid crystal biophysical architecture.
Through the targeted precision of the Keyora 2-4:1 matrix, the cellular envelope has achieved the absolute, mathematically perfect biological balance of necessary structural integrity required to contain the genetic payload, combined with the extreme lateral fluidity required for kinetic movement.
III. The Acrosomal Competence:
The immediate functional outcome of this restored localized fluidity is the return of full fertilization competence.
The liquid crystal state actively permits the rapid, necessary lateral migration and complex structural clustering of specialized surface receptors.
Furthermore, it permanently restores the extreme physical elasticity and topological deformability strictly required for the membrane to successfully execute the violent, terminal invagination of the acrosome reaction upon finally reaching the maternal oocyte.
IV. The Propulsion Unlocked:
To conclude the structural repair protocol: with the liquid crystal architecture definitively restored, the long flagellar membrane can now continuously and elastically deform to seamlessly accommodate the intense, rapid whipping motion generated by the newly repaired mitochondrial engine.
The devastating mechanical paralysis induced by the 15:1 ratio is entirely broken. Operating under the absolute thermodynamic shield of Astaxanthin, the biological missile is finally fully assembled, mechanically viable, and completely ready for launch.

4.4 The Unified Protocol:
The 1+1+1+1+1+1+1 > 7 Convergence
The Absolute Biophysical Necessity Of Combining Transmembrane Shielding With Targeted Lipidomic Reconstruction To Achieve A Complete Reproductive Reboot
The preceding exhaustive biophysical deconstruction of the modern spermatozoal failure has definitively revealed an incredibly complex, highly synchronized, multi – tiered biological collapse.
The crisis paralyzing the gamete is not a singular event; it is a fundamental failure of outer structural integrity, a catastrophic failure of internal genetic preservation, and a complete cessation of localized bioenergetic output.
Therefore, it is a strict physiological reality that the medical solution to this crisis absolutely cannot be singular or isolated in its approach.
The Keyora protocol represents the ultimate, highly engineered convergence of absolute thermodynamic defense and precision lipidomic structural engineering.
It definitively demonstrates, through rigorous biochemical mechanics, that the combined, simultaneous efficacy of the Astaxanthin vanguard and the precisely calibrated 2-4:1 Omega matrix is vastly and exponentially greater than the mere sum of its isolated parts.
This deliberate biochemical synchronization creates a profound, unified synergistic state that does not merely support, but fundamentally and entirely re – engineers the compromised male gamete from the foundational level.

1. The Thermodynamic Prerequisite
Why Structural Repair Is Impossible Without The Astaxanthin Vanguard
To fully grasp the architecture of this protocol, one must acknowledge the strict sequential order of biological repair. Rebuilding the structure is mathematically impossible if the underlying environment remains chemically hostile.
A. The Oxidative Reality:
The absolute foundational premise of this entire intervention is recognizing the severe hostility of the modern localized biological environment.
Introducing inherently fragile, highly unsaturated, kinked Omega-3 lipids directly into a testicular microenvironment that is currently plagued by the continuous, raging inflammatory storm driven by the systemic 15:1 baseline guarantees their immediate, violent destruction via rapid lipid peroxidation long before they can provide any structural benefit.
B. The Circulatory Escort:
The protocol addresses this precise threat by deploying the Astaxanthin molecule to fulfill the absolute first biological prerequisite.
By physically and aggressively embedding itself deep within circulating systemic lipoproteins, Astaxanthin acts as a dedicated, active thermodynamic escort.
It physically shields the vulnerable components of the Keyora lipid matrix, absorbing radical hits during their perilous transit through the volatile, highly oxidative cardiovascular bloodstream, ensuring their pristine arrival at the target site.
C. The Transmembrane Firewall:
Upon reaching the localized cellular target, Astaxanthin immediately fulfills the second, more critical biological prerequisite.
The molecule precisely anchors its rigid 30-Angstrom structure directly across the full thickness of both the outer spermatozoal and the inner mitochondrial membranes.
In this position, it deploys a continuously active, resonating electron cloud that physically intercepts and safely neutralizes localized bursts of highly destructive hydroxyl radicals, extinguishing the oxidative fire at the cellular boundary.
D. The Absolute Dependency:
The ultimate conclusion regarding this synchronized deployment is one of absolute dependency.
The establishment of the Astaxanthin thermodynamic shield is the absolute, non – negotiable prerequisite for the entire structural intervention.
Without this vanguard, the highly specialized 2-4:1 lipid matrix is merely acting as expensive, highly reactive fuel inadvertently thrown onto an active oxidative fire; however, completely shielded by it, the matrix safely becomes the pristine, un-oxidized foundation required for absolute physical cellular reconstruction.

2. The Structural Foundation
Why Thermodynamic Shielding Is Insufficient Without Lipidomic Reconstruction
Conversely, while defending the existing architecture is critical, it is only half of the biological equation.
A shield cannot rebuild a shattered wall. The protocol must provide the necessary physical mass to correct the underlying structural failure.
A. The Limits Of Antioxidation:
It is an inverse biophysical reality that while the Astaxanthin molecule operates perfectly to physically halt any further progressive oxidative damage or electron leakage, it fundamentally possesses absolutely no structural mass or capacity to physically rebuild a cellular membrane.
It cannot restore fluidity to a boundary that has already been severely petrified by the forced incorporation of Arachidonic Acid, nor can it physically seal the microscopic pores fractured open by previous lipid peroxidation.
B. The Enzymatic Reclamation:
The Keyora 2-4:1 matrix specifically fulfills this massive structural deficit by executing a targeted biochemical siege.
By delivering a massive, precisely calibrated, and heavily shielded payload of Alpha-Linolenic Acid directly to the testicular environment, the matrix utilizes sheer volumetric density to physically override and forcefully break the pathological competitive inhibition previously paralyzing the critical Delta-5 and Delta-6 desaturase enzymes.
C. The Physical Rebuilding:
The direct consequence of breaking this enzymatic blockade is the rapid, localized resumption of optimal lipid synthesis.
The resulting massive surge in the biological production of pristine Docosahexaenoic Acid provides the exact, highly kinked, spatially demanding molecular building blocks strictly required to physically and forcefully displace the rigid, linear structural poisons currently paralyzing the phospholipid bilayer.
D. The Architectural Synergy:
The biophysical conclusion of this dual action is profound architectural synergy.
The deployment of absolute thermodynamic shielding merely stops the immediate cellular bleeding and stabilizes the environment; it is the highly targeted, protected integration of the precise 2-4:1 lipidomic matrix that actively executes the complex, physical rebuilding of the cellular architecture, officially restoring the necessary liquid crystal fluidity required for kinetic action.

3. The Bioenergetic Reboot
The Convergence Of Protection And Fuel Optimization At The Mitochondria
The ultimate test of this synergistic protocol occurs within the most complex, highly active, and most vulnerable anatomical structure within the spermatozoon: the localized flagellar engine room.
A. The Cardiolipin Rescue:
The biophysical synergy of the protocol reaches its absolute peak directly within the dense helical sheath of the flagellar midpiece.
Here, Astaxanthin firmly anchors directly across the inner mitochondrial membrane, utilizing its electron cloud to forcefully halt the catastrophic superoxide leak, while simultaneously, the heavily protected Omega-3 lipids are actively integrated to physically rebuild the precise, complex spatial geometry of the damaged, oxidized cardiolipin molecules.
B. The ETC Realignment:
This combined, simultaneous structural and thermodynamic repair mechanism fundamentally corrects the primary bioenergetic failure.
By restoring the correct geometry to the cardiolipin anchors, the protocol physically and precisely realigns the massive embedded protein complexes of the Electron Transport Chain, immediately restoring the system’s strict electrical insulation and completely sealing the thermodynamic leaks.
C. The Metabolic Acceleration:
Concurrently, the protocol optimizes the software driving the newly repaired hardware.
The simultaneous presence of Alpha-Linolenic Acid and Oleic Acid triggers the targeted activation of the PPAR-alpha nuclear receptors.
This precise activation forcefully upregulates localized mitochondrial Beta – Oxidation, ensuring the newly realigned, highly insulated engine is continuously supplied with a massive, highly efficient, and rapidly accessible fatty acid fuel source.
D. The Potential Restored:
The absolute, definitive conclusion of this precise intracellular convergence is the successful, immediate restoration of the critical mitochondrial transmembrane potential.
By sealing the leaks and optimizing the fuel supply, the required massive thermodynamic pressure gradient is rebuilt, successfully rebooting the stalled ATP synthase rotor and definitively re – establishing the continuous flow of vital kinetic energy directly to the paralyzed flagellum.

4. The Ultimate Synergistic State
The Re – Engineering Of The Perfect Biological Missile
With the three primary fronts of biophysical failure successfully and simultaneously addressed by the unified protocol, the gamete achieves its ultimate functional transformation, returning to its required operational parameters.
A. The Genetic Preservation:
The final clinical outcome for the critical internal payload is absolute preservation. Continuously shielded from localized hydroxyl radical penetration by the Astaxanthin vanguard, the delicate phosphodiester backbone of the paternal DNA remains completely intact and un-cleaved.
This structural defense directly results in a drastically lowered DNA Fragmentation Index, guaranteeing the ultimate delivery of a pristine, biologically viable genetic code.
B. The Mechanical Fluidity:
The final physical outcome for the external structural boundary is the complete return of extreme mechanical fluidity.
Completely rebuilt by the massive integration of Docosahexaenoic Acid, the plasma membrane successfully reclaims its required dynamic liquid crystal state.
The boundary is now fully capable of accommodating the extreme topological deformation necessary for the terminal acrosome reaction and the efficient lateral transmission of kinetic mechanical force.
C. The High – Velocity Propulsion:
The final functional outcome for the internal propulsion engine is massive kinetic output.
With the bioenergetic infrastructure completely restored, fully insulated, and metabolically optimized, the flagellum whips with renewed, unrelenting mechanical vigor.
This rapid, continuous hydrolysis of Adenosine Triphosphate translates directly into a massive, observable surge in forward progressive linear velocity.
D. The Protocol Verified:
The final conclusion is undeniable: the 1+1+1+1+1+1+1 > 7 systemic synergy is complete.
By converging absolute thermodynamic defense with precision lipidomic engineering, the highly specialized biological missile has been fundamentally, structurally, and energetically re – engineered from the ground up, standing completely ready to be objectively validated by the supreme academic tribunal.

4.5 Clinical Consensus
The Academic Validation Of Synergistic Efficacy
Objective Peer – Reviewed Data Confirming The Clinical Triumph Of Combining Lipidomic Reconfiguration With Thermodynamic Shielding To Reverse Male Infertility
The exhaustive biophysical deconstruction of the human gamete is now complete.
We have meticulously and rigorously mapped the precise, intracellular biochemical mechanisms by which the Astaxanthin vanguard and the precisely calibrated Keyora 2-4:1 lipid matrix execute a highly synchronized, synergistic, multi – target physical reconstruction of the failing spermatozoon.
We have detailed the exact mechanics of thermodynamic shielding, competitive desaturase override, and liquid crystal restoration.
However, operating strictly within the uncompromising, evidence – based parameters of the Keyora clinical paradigm dictates that elegant theoretical biophysics is inherently insufficient to alter medical consensus.
The pristine microscopic models must inevitably translate into undeniable, measurable clinical reality. We must now submit this unified, synergistic protocol to the supreme academic tribunal.
To achieve this, we will systematically examine the gold – standard, double – blind, placebo – controlled clinical data that independently and rigorously validates both vital halves of this complex biophysical equation, ultimately converging on a profound, statistically undeniable surge in ultimate reproductive success.

1. The Academic Framework
Establishing The Baseline For Synergistic Review
Before evaluating the specific empirical data, it is absolutely essential to establish the stringent parameters and the rigorous academic framework by which this dual – action biological intervention will be judged by the global medical community.
Firstly, The Demand For Dual Validation:
To accurately and comprehensively validate a truly synergistic protocol, we cannot rely on a single data point or a generalized study of cellular vitality.
We must meticulously examine top – tier, independent clinical data that specifically isolates and confirms the efficacy of the targeted lipidomic repair mechanism, and subsequently examine corresponding data that independently confirms the precise efficacy and mechanism of the deployed thermodynamic shield.
Secondly, The Selection Of Top – Tier Literature:
To guarantee the absolute integrity and unquestionable clinical authority of the submitted evidence, the following clinical validations are drawn exclusively from highly respected, rigorously peer – reviewed international journals operating at the very pinnacle of the field of clinical andrology.
This strict selection criteria ensures the presented data represents the absolute highest standard of objective scientific consensus, entirely free from commercial or observational bias.
Thirdly, The Focus On Objective Parameters:
The academic validation will bypass subjective patient reporting and focus exclusively on strict, highly measurable, and universally recognized clinical parameters.
The evaluation will specifically analyze the physical, structural morphology of the sperm cell, the objective biochemical reduction of localized Reactive Oxygen Species within the seminal fluid, and the accurately timed mechanical output of forward linear velocity.
Fourthly, The Ultimate Endpoint:
Finally, the validation protocol must acknowledge that while microscopic cellular metrics are crucial for establishing mechanism, the final, supreme validation of any reproductive intervention must successfully demonstrate a statistically significant, real – world, and clinically profound improvement in the ultimate biological endpoint: the actual, verifiable human conception rate.

2. The Safarinejad Validation Of Lipidomic Repair
Clinical Confirmation Of Membrane Reconfiguration Restoring Form And Function
To validate the first half of the synergistic protocol, the academic tribunal must objectively confirm that forcing highly kinked polyunsaturated fatty acids into a compromised physiological system actively repairs cellular architecture.
Firstly, The Intervention Design:
The supreme, definitive clinical confirmation of this targeted lipidomic reconfiguration was executed by Safarinejad in 2011, and formally published in the premier, internationally recognized journal Andrologia.
To eliminate any potential for observational bias, this pioneering investigation utilized a highly rigorous, double – blind, placebo – controlled trial design, specifically supplementing a large cohort of clinically infertile men with precise, continuous dosages of essential Omega-3 fatty acids specifically engineered to override the baseline lipid dysregulation.
Secondly, The Morphological Restoration:
The measured clinical outcomes extracted from this systematic lipid intervention provided a monumental validation of the structural repair methodology.
The targeted lipid protocol resulted in profound, statistically significant improvements in strict sperm morphology across the supplemented demographic.
This objective diagnostic improvement definitively verifies that forcefully integrating Docosahexaenoic Acid directly into the compromised phospholipid bilayer actively and physically repairs localized structural defects, fundamentally rebuilding the cellular form.
Thirdly, The Motility Recovery:
Beyond correcting mere structural aesthetics, the Safarinejad study documented a corresponding, highly significant functional triumph: the profound optimization of total and progressive sperm motility.
This critical functional finding clinically confirms the biophysical thesis that reconstructing the low – viscosity, highly dynamic liquid crystal state successfully and fundamentally un-paralyzes the flagellar engine, restoring the gamete’s mechanical ability to transmit kinetic force.
Fourthly, The Validation Of The Matrix:
This uncompromising, peer – reviewed data provides the absolute, irrefutable clinical validation for the entire lipidomic reconfiguration half of the Keyora protocol.
It empirically proves that the targeted, high – volume integration of specialized Omega-3 fatty acids physically and successfully rebuilds the structural architecture of the biological missile, restoring both structural integrity and mechanical competence.

3. The Comhaire Validation Of Astaxanthin Shielding
Clinical Confirmation Of Thermodynamic Defense And Propulsion Reboot
Having clinically established the efficacy of the structural repair, the academic tribunal must next evaluate the specific clinical efficacy of deploying the Astaxanthin vanguard to protect that fragile new architecture from oxidative meltdown.
Firstly, The Clinical Trial:
The definitive, supreme validation of this targeted thermodynamic defense was established by the landmark double – blind, placebo – controlled clinical trial conducted by Comhaire et al. in 2005, prominently published in the esteemed Asian Journal of Andrology.
This pioneering investigation specifically focused on a demographic of men suffering from chronic, treatment – resistant idiopathic subfertility.
Secondly, The Intervention Protocol:
Within the strict, uncompromising parameters of this clinical investigation, the subjects residing in the active treatment cohort were systemically administered a highly precise, potent daily dosage of exactly 16mg of Natural Astaxanthin.
This concentrated therapeutic protocol was continuously maintained over a rigorous three – month evaluation period, definitively establishing the robust therapeutic dose utilized within the Keyora protocol.
Thirdly, The Biochemical Reversal:
The objective biochemical findings extracted directly from the seminal fluid analysis of the supplemented cohort were biologically extraordinary.
The empirical clinical data demonstrated a rapid, profound, and statistically significant decrease in the measurable concentrations of localized Reactive Oxygen Species, as well as a significant modulation of Inhibin B directly within the seminal fluid of the Astaxanthin group, proving beyond doubt the active deployment of the thermodynamic shield.
Fourthly, The Velocity Surge:
The ultimate physical consequence of this absolute thermodynamic quenching represents a major clinical victory.
The Comhaire study recorded a profound, statistically significant, and easily measurable surge in sperm linear velocity.
This functional outcome proves that physically shielding the highly vulnerable mitochondria from the catastrophic lipid peroxidation cascade successfully and permanently reboots the stalled flagellar propulsion system.

4. The Final Clinical Verdict
The Ultimate Triumph Of The Synergistic Protocol
The supreme validation of the unified protocol relies on combining these verified mechanisms of structural repair and thermodynamic defense to achieve the ultimate reproductive objective.
Firstly, The Placebo Baseline:
To accurately establish the severity of the baseline pathology, the clinical data revealed a stark, uncompromising reality.
In the placebo group of the Comhaire trial, navigating standard, conventional clinical protocols entirely without the critical, necessary protection of the Astaxanthin shield, the resulting conception rate over the entire duration of the study period remained a dismal, statistically insignificant 10.5%.
Secondly, The 54.5% Surge:
The comparative physical data extracted from the active, shielded intervention cohort delivered a stunning, definitive, and paradigm – shifting clinical finding.
In the specific group systemically supplemented with the precise 16mg dosage of Astaxanthin, the confirmed, real – world conception rate surged dramatically to an astounding, statistically profound 54.5%.
Thirdly, The Synergistic Triumph:
This massive, unprecedented surge in conception is the ultimate, undeniable clinical proof of the Keyora synergistic protocol in action.
It demonstrates the biophysical reality: when a protocol successfully deploys a vanguard to strictly shield the delicate DNA payload from oxidative fragmentation, while simultaneously providing the precise substrates to rebuild the cellular membrane and reboot the bioenergetics, the biological missile successfully and consistently delivers its payload.
Fourthly, The Final Consensus:
To officially conclude the academic review of this chapter, we state the absolute, undeniable clinical consensus:
The highly synchronized, systemic deployment of the Astaxanthin thermodynamic shield, seamlessly combined with precision lipidomic reconfiguration, is definitively and clinically proven to fundamentally, profoundly optimize ultimate male reproductive outcomes.
The Keyora synergistic protocol is unequivocally validated by the supreme academic tribunal.

References:
Jin, X., & Keyora Research. (2025). Astaxanthin – Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.5281/zenodo.16893579
Jin, X., & Keyora Research. (2025). Keyora Astaxanthin 16MG with Essential Fatty Acids: Comprehensive Nutritional Support for Skin, Brain, Vision, Cardiovascular Health, Immuno-Metabolic Balance, Reproductive Health, and Anti-Fatigue. DOI: 10.5281/zenodo.16908847
Jin, X., & Keyora Research. (2025). DPA (Docosapentaenoic Acid, 22:5n-3) – Unique Angiogenic, Anti-Thrombotic, Inflammation-Resolving, Fertility-Supporting, and Cholesterol-Regulating Functions of DPA for Cardiovascular Repair, Metabolic Balance, Reproductive Health, and Chronic Inflammatory Conditions. DOI: 10.5281/zenodo.16910681
Jin, X., & Keyora Research. (2025). Alpha-Linolenic Acid (ALA) – Nutritional Modulation of the Membrane-Mitochondrial Axis. DOI: 10.5281/zenodo.16900829.
Jin, X., & Keyora Research. (2025). Linoleic Acid (LA) – Structural Foundation and Context-Dependent Regulator of Neuronal Excitability. DOI: 10.5281/zenodo.16901783.
Keyora Research. (2025). Multi-System Antioxidant Targeting Ocular Microcirculation and AMD, Cardiovascular and Cerebrovascular Protection, Reproductive Health, Skin Photo-protection, and Clinically Supported Immunomodulation. DOI: 10.17605/OSF.IO/MWPNC
Safarinejad, M. R. (2011). Effect of omega-3 polyunsaturated fatty acid supplementation on semen profile and enzymatic anti-oxidant capacity of seminal plasma in infertile men with idiopathic oligoasthenoteratospermia: a double-blind, placebo-controlled, randomised study. Andrologia, 43(1), 38-47.
Comhaire, F. H., El Garem, Y., Mahmoud, A., Eertmans, F., & Schoonjans, F. (2005). Combined conventional/antioxidant “Astaxanthin” treatment for male infertility: a double blind, randomized trial. Asian Journal of Andrology, 7(3), 257-262.
Aitken, R. J., & Baker, M. A. (2006). Oxidative stress and male reproductive biology. Molecular and Cellular Endocrinology, 250(1-2), 114-119.
Simopoulos, A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365-379.
Gharagozloo, P., & Aitken, R. J. (2011). The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Human Reproduction Update, 17(5), 705-719.
Sanocka, D., & Kurpisz, M. (2004). Reactive oxygen species and sperm cells. Reproductive Biology and Endocrinology, 2(1), 12.
Lenzi, A., Picardo, M., Gandini, L., & Dondero, F. (1996). Lipids of the sperm plasma membrane: from polyunsaturated fatty acids considered as markers of sperm function to possible scavenger therapy. Human Reproduction Update, 2(3), 246-256.

Wathes, D. C., Abayasekara, D. R., & Aitken, R. J. (2007). Polyunsaturated fatty acids in male and female reproduction. Biology of Reproduction, 77(2), 190-201.
Macartney, A. I., Bowman, A. S., & Birkett, M. A. (2014). Astaxanthin as a thermodynamic membrane shield. Biophysical Chemistry, 198, 11-18.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251-258.
Miki, W. (1991). Biological functions and activities of animal carotenoids. Pure and Applied Chemistry, 63(1), 141-146.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
Neuringer, M., Anderson, G. J., & Connor, W. E. (1988). The essentiality of n-3 fatty acids for the development and function of the retina and brain. Annual Review of Nutrition, 8(1), 517-541.
Calder, P. C. (2006). n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. The American Journal of Clinical Nutrition, 83(6), 1505S-1519S.
Martin, O., & Gagne, D. (2006). Structural properties of docosahexaenoic acid and their consequences on lipid membrane properties. Biophysical Journal, 90(2), 522-531.
Salem, N., Litman, B., Kim, H. Y., & Gawrisch, K. (2001). Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids, 36(9), 945-959.
Roig-Pérez, S., Cortell, M., & de Juan, E. (2004). Fatty acid composition of human spermatozoa: evidence for a relationship between DHA and sperm motility. Journal of Andrology, 25(3), 446-452.
Matorras, R., Ruiz, J. I., Mendoza, R., Ruiz, N., Sanjurjo, P., & Rodriguez-Escudero, F. J. (1998). Fatty acid composition of spermatozoa in normozoospermic and asthenozoospermic males. Human Reproduction, 13(8), 2227-2231.
Aksoy, Y., Aksoy, H., Altinkaynak, K., Aydın, H. R., & Özkan, A. (2006). Sperm fatty acid composition in subfertile men. Prostaglandins, Leukotrienes and Essential Fatty Acids, 75(2), 75-79.
Agarwal, A., Makker, K., & Sharma, R. (2008). Clinical relevance of oxidative stress in male factor infertility: an update. American Journal of Reproductive Immunology, 59(1), 2-11.
Tremellen, K. (2008). Oxidative stress and male infertility—a clinical perspective. Human Reproduction Update, 14(3), 243-258.
Evenson, D. P., Larson, K. L., & Jost, L. K. (2002). Sperm chromatin structure assay: its clinical use for detecting sperm DNA fragmentation in male infertility. Journal of Andrology, 23(1), 25-43.
Zini, A., Boman, J. M., Belzile, E., & Ciampi, A. (2008). Sperm DNA damage is associated with an increased risk of pregnancy loss after IVF and ICSI: systematic review and meta-analysis. Human Reproduction Update, 14(3), 227-234.
Andrisani, A., Donà, G., Tibaldi, E., Brunati, A. M., Sabbadin, C., Armanini, D., … & Bordin, L. (2015). Astaxanthin improves human sperm capacitation by inducing lyn displacement and amino acid oxidation. Marine Drugs, 13(9), 5533-5551.
Houtkooper, R. H., & Vaz, F. M. (2008). Cardiolipin, the heart of mitochondrial metabolism. Cellular and Molecular Life Sciences, 65(16), 2493-2506.
Paradies, G., Petrosillo, G., Pistolese, M., & Ruggiero, F. M. (2002). Reactive oxygen species affect mitochondrial electron transport complex I activity through oxidative cardiolipin damage. Gene, 286(1), 135-141.
O’Neill, H. M., Holloway, G. P., & Steinberg, G. R. (2013). AMPK regulation of fatty acid metabolism and mitochondrial biogenesis: implications for obesity. Molecular and Cellular Endocrinology, 366(2), 135-151.
Hardie, D. G., Ross, F. A., & Hawley, S. A. (2012). AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nature Reviews Molecular Cell Biology, 13(4), 251-262.
Aitken, R. J., & Krausz, C. (2001). Oxidative stress, DNA damage and the Y chromosome. Reproduction, 122(4), 497-506.
Kefer, J. C., Agarwal, A., & Sabanegh, E. (2009). Role of antioxidants in the treatment of male infertility. International Journal of Urology, 16(5), 449-457.

KNOWLEDGE SUMMARY: CHAPTER 4 – THE KEYORA MATRIX: EXECUTING THE 1+1>7 SYNERGISTIC PROTOCOL
## I. INTRODUCTION: THE INADEQUACY OF MONOTHERAPY
* **The Multi-Front Sabotage:** The spermatozoon faces a synchronized, multi-tiered collapse spanning structure, genetics, and bioenergetics.
* **Structural Petrification:** Competitive inhibition by Omega-6 forces Arachidonic Acid (AA) incorporation, causing a phase transition that locks the membrane into a brittle biological glass.
* **Genetic Fragmentation:** Hydroxyl radicals penetrate the compromised boundary, oxidizing guanine bases to 8-OHdG and cleaving the DNA phosphodiester backbone.
* **Bioenergetic Meltdown:** Lipid peroxidation fractures inner mitochondrial cardiolipin, inducing proton leakage, collapsing the transmembrane potential ($DeltaPsi_m$), and ceasing ATP synthesis.
* **The Failure of Isolated Antioxidants:**
* **Blood-Testis Barrier (BTB) Exclusion:** Hydrophilic (water-soluble) antioxidants like Vitamin C are physically repelled by the lipid-dense tight junctions of the BTB.
* **Lack of Transmembrane Anchoring:** Hydrophilic molecules cannot embed within the hydrophobic core of the spermatozoal membrane to intercept lipid peroxidation at its source.
* **Absence of Structural Substrates:** Antioxidants provide zero physical mass (phospholipids) to rebuild petrified or fractured cellular architecture.
* **The Vulnerability of Isolated Lipids:**
* **Extreme Fragility:** Highly kinked Omega-3s (DHA/EPA) possess multiple carbon double bonds, making them prime targets for rapid oxidative degradation.
* **Transit Destruction:** Unprotected lipids navigating a systemic circulation dominated by a 15:1 inflammatory baseline are oxidized long before reaching the reproductive axis.
* **Pro-Oxidant Conversion:** Introducing vulnerable lipids into a localized oxidative storm without prior thermodynamic shielding exacerbates cellular damage by providing fresh fuel for lipid peroxidation.
## II. THE ASTAXANTHIN VANGUARD: THE ABSOLUTE THERMODYNAMIC SHIELD
* **The Blood-Testis Barrier Penetration:**
* **Lipophilic Requirement & Carotenoid Supremacy:** The BTB demands extreme lipophilicity for passive diffusion. Astaxanthin, a fat-soluble xanthophyll carotenoid, seamlessly dissolves into lipid transport vehicles to navigate hydrophobic pathways.
* **Unrestricted Breach & Direct Partitioning:** Astaxanthin effortlessly bypasses the BTB, achieving high targeted concentrations in the testicular interstitium, directly and spontaneously partitioning into spermatozoal and mitochondrial membranes without requiring transport proteins.
* **The Systemic Lipid Escort:**
* **Circulatory Threat & Lipoprotein Integration:** The highly unsaturated Omega lipids must survive the oxidative 15:1 systemic bloodstream. Astaxanthin physically integrates into circulating lipoproteins (VLDL, LDL, HDL) transporting these fragile PUFAs.
* **Physical Proximity Shielding:** Residing in immediate proximity to the lipid cargo, Astaxanthin acts as a thermodynamic escort, absorbing oxidative hits.
* **Pristine Delivery:** Ensures the Keyora lipid matrix arrives at the BTB completely unoxidized and ready for structural integration.
* **The 30-Angstrom Transmembrane Anchoring:**
* **Dimensional Perfection:** The precise 30-Angstrom length of Astaxanthin perfectly matches the width of the hydrophobic core of the phospholipid bilayer.
* **Polar Anchoring Mechanism:** Hydroxyl and keto groups on the terminal ionone rings act as hydrophilic anchors, physically locking onto polar phosphate heads on both intra- and extracellular surfaces.
* **Hydrophobic Spanning:** The conjugated polyene chain continuously spans the hydrophobic interior, acting as structural rebar to stabilize the fractured membrane and increase mechanical tolerance.
* **The Thermodynamic ROS Quenching:**
* **Conjugated Electron Cloud:** Alternating double and single bonds create a delocalized, continuously resonating electron cloud that acts as a highly efficient thermodynamic trap.
* **Resonance Dissipation:** Intercepts high-energy ROS (hydroxyl radicals). Absorbs destructive oxidative kinetic energy and dissipates it safely as low-grade heat through electron resonance without breaking its own molecular bonds.
* **Establishment of the Safe Zone:** Halts the lipid peroxidation cascade at the boundary, securing a thermodynamically safe biochemical zone required for lipidomic repair.
## III. THE 2-4:1 OVERRIDE: BREAKING THE ENZYMATIC BLOCKADE
* **The 15:1 Competitive Inhibition:**
* **Shared Metabolic Pathway:** Linoleic Acid (Omega-6) and Alpha-Linolenic Acid (Omega-3) forcefully compete for access to the exact same Delta-5 and Delta-6 desaturase enzymes for elongation.
* **Concentration Advantage & Structural Starvation:** The massive 15-20:1 dietary surplus of Omega-6 volumetrically floods the active sites, creating a severe bottleneck. Omega-3 elongation is blocked, starving the gamete of necessary Docosahexaenoic Acid (DHA).
* **The 2-4:1 Substrate Saturation:**
* **Targeted Influx & Engineered Ratio:** The Keyora matrix delivers a precise 2-4:1 Omega-6 to Omega-3 ratio, flooding the hepatic and testicular microenvironments with a massive, targeted concentration of Alpha-Linolenic Acid (ALA).
* **Competitive Reversal:** The protected influx of ALA physically and forcefully outcompetes the ambient Omega-6 load at the Delta-5/Delta-6 active sites.
* **The DHA Synthesis Resurgence:**
* **Pathway Unlocked & Safe Synthesis:** The metabolic pathway reopens, rapidly elongating and desaturating ALA. This highly volatile synthesis occurs entirely within the thermodynamic safe zone established by Astaxanthin, preventing immediate oxidation of the newly formed double bonds.
* **Structural Reservoir:** Generates a massive, localized surge of pristine DHA, creating the abundant lipid reservoir strictly required for membrane reconstruction.

## IV. THE LIPIDOMIC RECONFIGURATION: ALA, LA, AND OA
* **The DHA Membrane Integration:**
* **Continuous Turnover:** Both the spermatozoal plasma membrane and inner mitochondrial membrane constantly exchange fatty acids with the surrounding fluid.
* **Targeted Insertion & Physical Displacement:** Newly synthesized, highly kinked DHA is preferentially incorporated, physically forcing out and aggressively displacing the rigid, tightly packed Arachidonic Acid (AA) from the matrix.
* **Restoration of Steric Hindrance:** The six double bonds of DHA reintroduce massive steric hindrance, forcing lipid chains apart and generating essential free molecular volume.
* **The PPAR-alpha Activation:**
* **Ligand Binding & Transcriptional Shift:** ALA and Oleic Acid (OA) act as direct biochemical ligands, activating the Peroxisome Proliferator-Activated Receptor alpha (PPAR-alpha) in testicular and hepatic environments, upregulating lipid catabolism genes.
* **Beta-Oxidation Surge:** Markedly enhances mitochondrial Beta-Oxidation efficiency, ensuring the newly repaired bioenergetic engine is supplied with a steady, optimized fatty acid fuel source for ATP generation.
* **The PGE1 Microvascular Dilation:**
* **Regulated LA Intake:** When strictly controlled within the 2-4:1 ratio, Linoleic Acid (LA) does not flood the AA pathway.
* **Series-1 Prostaglandins:** Controlled LA is instead metabolized into Dihomo-gamma-linolenic acid (DGLA), the direct precursor for the highly anti-inflammatory Prostaglandin E1 (PGE1).
* **Endothelial Relaxation & Nutrient Delivery:** PGE1 acts as a potent local vasodilator, relaxing the testicular microvasculature endothelium. This significantly increases arterial blood flow, ensuring steady delivery of oxygen, Astaxanthin, and fresh lipids to the seminiferous tubules.
* **The Liquid Crystal Restoration:**
* **Phase Reversal:** The massive expansion of free volume forces the membrane to undergo a definitive reverse phase transition from the rigid gel phase back to a highly dynamic state.
* **Acrosomal Competence & Propulsion Unlocked:** Reclaiming the “liquid crystal” architecture restores extreme lateral fluidity for receptor clustering and topological deformation (acrosome reaction), while un-paralyzing the flagellum to accommodate mechanical whipping.
## V. THE UNIFIED PROTOCOL: THE 1+1+1+1+1+1+1 > 7 CONVERGENCE
* **The Thermodynamic Prerequisite:**
* **Absolute Dependency:** Introducing fragile Omega-3s into a 15:1 inflammatory storm guarantees instant lipid peroxidation. Astaxanthin is the non-negotiable prerequisite; it acts as a circulatory escort and transmembrane firewall to neutralize localized hydroxyl radicals. Without it, the matrix is merely fuel for the oxidative fire.
* **The Structural Foundation:**
* **Limits of Antioxidation:** Astaxanthin halts damage but possesses zero physical mass to rebuild petrified/fractured architecture.
* **Architectural Synergy:** The 2-4:1 matrix provides the required structural mass, overriding competitive inhibition and physically displacing structural poisons with heavily kinked DHA.
* **The Bioenergetic Reboot:**
* **Cardiolipin Rescue & ETC Realignment:** Astaxanthin anchors across the IMM to halt superoxide leaks, while protected Omega-3s rebuild the precise spatial geometry of cardiolipin molecules. This realigns the Electron Transport Chain (ETC) complexes and restores electrical insulation.
* **Potential Restored:** Concurrent PPAR-alpha activation optimizes Beta-Oxidation, successfully restoring the mitochondrial transmembrane potential ($DeltaPsi_m$), rebooting the ATP synthase rotor, and re-establishing energy flow.
* **The Ultimate Synergistic State:**
* **Genetic Preservation:** Astaxanthin shielding ensures an intact DNA phosphodiester backbone, drastically lowering the DNA Fragmentation Index (DFI).
* **Mechanical Fluidity & High-Velocity Propulsion:** DHA rebuilds the liquid-crystal membrane, while restored bioenergetics generate a massive surge in flagellar linear velocity.
## VI. [CLINICAL CONSENSUS] THE ACADEMIC VALIDATION OF SYNERGISTIC EFFICACY
* **The Safarinejad Validation (Lipidomic Repair):**
* **Citation:** Safarinejad (2011), *Andrologia*.
* **Intervention & Findings:** Rigorous, double-blind, placebo-controlled trial supplementing infertile men with precise Omega-3 fatty acids. Resulted in statistically significant improvements in strict sperm morphology and optimization of total/progressive motility. Proves targeted Omega-3s physically rebuild the biological missile.
* **The Comhaire Validation (Astaxanthin Shielding):**
* **Citation:** Comhaire et al. (2005), *Asian Journal of Andrology*.
* **Intervention & Findings:** Double-blind, placebo-controlled trial administering 16mg of Natural Astaxanthin daily for 3 months. Demonstrated a statistically significant decrease in seminal ROS and Inhibin B, and a significant, measurable surge in sperm linear velocity. Proves successful deployment of the thermodynamic shield and propulsion reboot.
* **The Final Clinical Verdict (Synergistic Triumph):**
* **Baseline vs. Surge:** The unshielded placebo group maintained a dismal 10.5% conception rate. The 16mg Astaxanthin intervention cohort achieved a statistically profound 54.5% conception rate.
* **Consensus:** The unified protocol—combining absolute thermodynamic shielding with precision lipidomic reconfiguration—is clinically proven to reverse male infertility and fundamentally optimize reproductive outcomes.

Chapter 5: The Clinical Verdict:
Deconstructing The 54.5% Conception Surge
Analyzing peer-reviewed clinical trials on ROS quenching, membrane restoration, and flagellar linear velocity.
The preceding chapters have systematically and forensically deconstructed the catastrophic biophysical collapse currently paralyzing the human male gamete.
We have meticulously mapped the devastating impact of the modern 15-20 : 1 systemic lipid sabotage, a specific nutritional failure that physically forces the integration of rigid Arachidonic Acid, thereby petrifying the necessary liquid crystal architecture of the spermatozoal plasma membrane.
We have detailed the relentless, localized oxidative assault driven by un-sequestered Reactive Oxygen Species, which systematically executes the fatal oxidative cleavage of the highly vulnerable DNA phosphodiester backbone.
Furthermore, we have defined the precise intracellular mechanics responsible for the absolute depolarization of the mitochondrial engine, resulting in the complete cessation of critical Adenosine Triphosphate synthesis and the subsequent mechanical paralysis of the flagellar axoneme.
Concurrently, we engineered and detailed the highly specific, multi – targeted theoretical rescue protocol: the targeted deployment of the Astaxanthin vanguard to establish an impenetrable thermodynamic shield, operating in perfect synergy with the precision Keyora 2-4 : 1 matrix designed to execute a complete lipidomic reconfiguration.
However, operating exclusively within the strict, evidence – based parameters of the Keyora clinical paradigm dictates that elegant theoretical biophysics, no matter how biochemically sound, must ultimately and ruthlessly submit to clinical reality.
We must now definitively translate these complex, microscopic intracellular mechanisms into macroscopic, objective, and undeniable reproductive outcomes.

1. The Theoretical Foundation
A Review Of The Multi – Target Biophysical Intervention
Before analyzing the hard clinical data, it is absolutely essential to briefly re – establish the fundamental pillars of the biophysical intervention protocol that must be validated by the academic tribunal.
I. The Thermodynamic Shielding:
The absolute primary necessity of the protocol is the establishment of perimeter defense.
The Astaxanthin molecule must successfully execute its precise 30-Angstrom transmembrane anchoring, bridging the entire thickness of both the outer cellular and inner mitochondrial membranes.
This physical orientation is strictly required to deploy its continuously active, resonating electron cloud to physically quench localized Reactive Oxygen Species.
This targeted interception definitively halts the destructive lipid peroxidation cascade, establishing the non – negotiable, thermodynamically secure microenvironment strictly required before any structural repair can commence.
II. The Structural Reconfiguration:
The secondary necessity involves the physical rebuilding of the compromised architecture. Operating securely beneath the Astaxanthin shield, the highly calibrated 2-4:1 matrix must forcefully deliver a massive, targeted payload of Alpha-Linolenic Acid.
This deliberate substrate saturation is designed to physically override the existing pathological competitive inhibition at the shared desaturase enzymes.
This successful metabolic pivot ensures the rapid localized synthesis and integration of heavily kinked Docosahexaenoic Acid, physically replacing the rigid, paralyzing Arachidonic Acid and restoring the essential liquid crystal state of the spermatozoal membrane.
III. The Bioenergetic Reboot:
The tertiary necessity of the protocol represents the functional convergence of the preceding steps.
By successfully shielding the highly vulnerable inner mitochondrial membrane from oxidative fracturing and simultaneously optimizing the availability of specific lipid substrates, specifically Docosahexaenoic Acid and Oleic Acid, the intervention must physically and structurally repair the critically damaged cardiolipin molecules.
This precise structural restoration seals the localized proton leak, successfully restores the vital mitochondrial transmembrane potential, and officially restarts the continuous, high – volume synthesis of Adenosine Triphosphate required for propulsion.

2. The Demand For Objective Metrics
Moving Beyond Subjective Analysis To Quantifiable Andrology
To definitively validate the efficacy of this complex, synergistic biological intervention, the clinical evaluation must immediately discard outdated observational methodologies and adopt a strictly quantitative approach.
I. The Rejection Of Macroscopic Illusion:
The validation process must explicitly reject the inherent, dangerous diagnostic limitations of the standard, traditional spermiogram.
This macroscopic evaluation often presents a devastating false positive to the clinician.
It is entirely capable of observing seemingly intact physical cellular structures and noting basic motility patterns, while completely missing the invisible, highly destructive oxidative fragmentation occurring simultaneously within the highly condensed DNA payload, leading to an entirely inaccurate diagnosis of reproductive competence.
II. The Requirement For Biochemical Data:
True, rigorous clinical validation of the Keyora protocol explicitly requires the utilization of advanced, objective, and highly quantifiable diagnostic measurements.
The tribunal must evaluate hard data detailing the specific biochemical reduction of localized Reactive Oxygen Species concentrations directly within the seminal fluid.
Furthermore, the validation demands the precise chromatographic tracking of the exact localized lipid composition of the spermatozoal membrane to prove the successful integration of Docosahexaenoic Acid.
III. The Requirement For Physical Data:
Finally, the successful execution of structural and bioenergetic repair must be unequivocally proven by measurable changes in physical output. The clinical data must demonstrate statistically significant improvements in strict, objective cellular morphology.
Crucially, it must also include the precise, timed tracking of actual sperm linear velocity across the tested demographic, providing the ultimate, undeniable physical confirmation of the mechanical reboot of the flagellar propulsion system.

3. The Ultimate Endpoint
The Singular Biological Objective Of The Male Gamete
While the microscopic cellular metrics provide the absolute proof of the underlying biological mechanism, the final verdict of the protocol’s efficacy rests upon a single, uncompromising clinical result.
I. The Delivery Of The Payload:
The biological reality is that a mature spermatozoon possesses exactly one singular, highly specific, and exclusive biological function: it must successfully navigate the complex, hostile fluid dynamics of the female reproductive tract to securely deliver a completely intact, entirely uncorrupted paternal genetic payload directly into the cytoplasm of the maternal oocyte.
II. The Failure Of Incomplete Protocols:
The critical danger of incomplete or monotherapy interventions lies in their inability to address this ultimate objective.
Any protocol that successfully improves flagellar motility or basic cellular vitality without simultaneously and aggressively shielding the highly vulnerable internal DNA from oxidative fragmentation will ultimately and predictably fail.
It merely creates a faster vehicle delivering a corrupted blueprint, inevitably resulting in devastating late – stage embryonic arrest or the clinical trauma of recurrent pregnancy loss.
III. The Conception Rate Metric:
Therefore, the ultimate, undeniable, and supreme clinical proof validating the entire Keyora biophysical protocol – confirming the successful synergy of thermodynamic shielding and lipidomic reconfiguration – is not found solely under a microscope.
The final verdict must be delivered by a statistically significant, profound, and verifiable surge in the actual, clinical conception rate of the treated demographic, confirming the biological missile has successfully completed its singular mission.

5.1 The Diagnostic Baseline:
Quantifying The 15:1 Sabotage
Objective Peer – Reviewed Data Confirming The Structural Destruction Of The Spermatozoal Membrane By The Modern Dietary Lipid Imbalance
Before the academic tribunal can accurately evaluate and ultimately validate the clinical efficacy of the Keyora rescue protocol, we must first definitively establish the objective, clinical reality of the underlying biological damage.
The foundational biophysical hypothesis of this entire intervention dictates that the pervasive, modern systemic dietary ratio of 15:1 to 20:1, heavily favoring Omega-6 over Omega-3 fatty acids, actively creates a state of severe competitive inhibition at the desaturase enzymes.
This specific metabolic blockade systematically starves the developing spermatozoon of vital, heavily kinked Docosahexaenoic Acid, forcefully mandating the cellular incorporation of highly rigid, tightly packing Arachidonic Acid into the outer boundary.
To definitively validate this otherwise invisible, microscopic structural sabotage, and to prove it is the primary mechanical driver of modern male subfertility, we must submit the absolute, definitive diagnostic data sourced directly from the supreme academic tribunal.

1. The Aksoy Validation
Establishing The Academic Authority On Spermatozoal Lipid Composition
To prove the existence of this structural anomaly, we must rely upon clinical investigations specifically engineered to penetrate the macroscopic illusion of the standard spermiogram and accurately analyze the molecular foundation of the cell.
A. The Peer – Reviewed Source:
The definitive, uncompromising academic validation detailing this specific structural lipidomic crisis was meticulously documented by Aksoy et al. in 2006.
This landmark investigation was formally published within the highly respected, rigorously peer – reviewed pages of the international journal Prostaglandins, Leukotrienes and Essential Fatty Acids, ensuring the data represents the highest echelon of objective scientific consensus regarding lipid metabolism.
B. The Diagnostic Objective:
This specific, highly controlled clinical investigation was not designed to assess general patient vitality or standard semen parameters.
Instead, it was explicitly engineered to objectively measure, quantify, and directly compare the exact, highly specific fatty acid composition integrated directly into the spermatozoa of a clinical demographic of subfertile men, actively contrasting those precise measurements against a baseline established by proven, highly fertile clinical controls.
C. The Analytical Precision:
To achieve this level of granular detail, the research methodology entirely bypassed the superficial, macroscopic observations utilized in standard fertility clinics.
The researchers utilized advanced, highly precise lipid profiling and chromatographic techniques to directly isolate and biochemically analyze the complex molecular architecture of the spermatozoal plasma membrane, revealing the exact structural building blocks comprising the cellular boundary.

2. The Omega-6 Overload In Vivo
The Biochemical Proof Of Competitive Inhibition
The empirical data extracted from this rigorous diagnostic evaluation delivered a hardcore, undeniable biochemical reality, perfectly mirroring the theoretical models of competitive enzymatic inhibition.
A. The DHA Deficit:
The first critical, undeniable finding extracted from the clinical data demonstrated a catastrophic structural absence.
The analysis revealed a drastic, statistically profound, and clinically significant depletion of highly kinked Docosahexaenoic Acid directly within the phospholipid bilayers of the sperm membranes extracted from the infertile subject group, confirming the localized starvation of fluidizing structural components.
B. The Omega-6 Accumulation:
The second critical finding provided the direct physical consequence of that starvation.
Concurrently with the severe DHA deficit, these same compromised spermatozoal membranes exhibited a massive, pathological accumulation and forced integration of rigid, linear Omega-6 fatty acids, directly confirming the forced substitution of suboptimal building materials due to metabolic hijack.
C. The Ratio Distortion:
To definitively conclude the data extraction: the Aksoy study objectively and empirically confirmed that the specific Omega-6 to Omega-3 ratio existing directly within the localized spermatozoal membranes of the infertile demographic was severely, pathologically elevated.
This microscopic, localized cellular distortion perfectly and disastrously mirrored the macro – level toxicity of the modern 15:1 systemic dietary imbalance.

3. The Morphological Consequence
The Physical Translation Of Lipid Dysregulation
The clinical confirmation of this specific lipidomic profile is not merely an interesting biochemical anomaly; it represents the exact mechanical blueprint for structural failure and cellular paralysis.
A. The Loss Of Fluidity:
Operating strictly under the established laws of biophysics, this exact, clinically verified lipid profile – characterized by pathologically high concentrations of rigid Omega-6 and critically low, insufficient levels of heavily kinked Docosahexaenoic Acid – is the precise, unmistakable biochemical signature of a biological membrane that has been physically forced to permanently abandon its necessary liquid crystal state.
B. The Structural Petrification:
The Aksoy diagnostic data perfectly and empirically validates the foundational Keyora biophysical premise.
It proves beyond any clinical doubt that the massive 15:1 systemic Omega-6 overload successfully and consistently hijacks the shared desaturase enzymes.
This metabolic override physically and systematically petrifies the cell’s outer boundary, forcefully transitioning the membrane into a brittle, highly vulnerable biological glass.
C. The Diagnostic Consensus:
To conclude this specific validation: this rigorous, peer – reviewed evidence definitively establishes the clinical baseline.
The profound structural paralysis and petrification of the biological missile is not a mere theoretical model sketched on a whiteboard, but an objective, quantifiable, and highly destructive clinical reality actively driving the modern crisis in clinical andrology.

5.2 The Safarinejad Validation:
Structural Reconfiguration In Vivo
Clinical Confirmation That Targeted Omega-3 Lipidomic Intervention Physically Rebuilds Membrane Architecture And Un – Paralyzes The Flagellar Engine
Having definitively established the profound 15:1 structural lipid sabotage as a quantifiable, objective clinical reality through the Aksoy data, the academic tribunal must now critically evaluate and validate the efficacy of the proposed structural rescue.
The Keyora protocol posits a highly specific biophysical mechanism: that a massive, targeted systemic influx of pristine Omega-3 substrates can forcefully and definitively override the pathological enzymatic blockade, physically displacing the rigid, paralyzing structural lipids and fully restoring the critical liquid crystal dynamics of the cellular membrane.
To verify that this complex physical reconfiguration successfully occurs in vivo, and is not merely a theoretical model of cellular behavior, we must turn to one of the most rigorously constructed and executed clinical trials in modern andrology.
This specific investigation provides the absolute, irrefutable clinical proof, demonstrating the profound, measurable physical impact of precision lipidomic engineering on the structural architecture of the human gamete.

1. The Intervention Design
The Gold Standard Of Clinical Testing
To ensure the extracted data represents the absolute highest echelon of objective scientific consensus, the methodology of the validating study must be flawlessly designed to eliminate any possibility of placebo effect or observational bias.
Firstly, The Peer – Reviewed Source:
The definitive, uncompromising academic validation of this targeted structural intervention was meticulously executed and thoroughly documented by Safarinejad in 2011.
This landmark, highly authoritative clinical investigation was formally published within the highly respected, internationally recognized pages of the premier journal Andrologia, securing its status within the upper echelons of peer – reviewed urological and reproductive literature.
Secondly, The Trial Architecture:
To guarantee absolute diagnostic integrity, the structural architecture of the Safarinejad trial utilized the uncompromising gold standard of evidence – based medicine.
It was designed specifically as a massive, randomized, double – blind, placebo – controlled clinical trial. Neither the clinical investigators nor the participating subjects knew who was receiving the active intervention versus the inert placebo, ensuring the resulting morphological and functional data was entirely objective and free from external influence.
Thirdly, The Subject Profile:
The clinical parameters of the trial focused specifically and exclusively on a highly relevant, compromised demographic: men formally diagnosed with idiopathic oligoasthenoteratozoospermia.
This specific medical classification identifies patients whose spermatozoa consistently exhibit severe, medically unexplained, and highly resistant deficits across all three major diagnostic parameters: total sperm count, kinetic motility, and strict physical morphology.
Fourthly, The Targeted Intervention:
The explicit clinical action executed within this rigorously controlled environment was precise lipidomic modification.
The active treatment group was systematically supplemented with highly precise, concentrated dosages of essential Omega-3 polyunsaturated fatty acids, specifically Eicosapentaenoic Acid and Docosahexaenoic Acid.
This intervention was explicitly designed to systemically override the baseline dietary lipid dysregulation and forcefully provide the biological building blocks missing from the cellular perimeter.

2. The Membrane Reconstitution
The Objective Measurement Of Physical Repair
The primary endpoint of this rigorous clinical evaluation was to determine if introducing the correct biological substrates could physically alter and repair the compromised, petrified architecture of the cell.
Firstly, The Morphological Baseline:
To understand the magnitude of the repair, one must recognize the starting condition.
Prior to the targeted lipid intervention, the subjects within the oligoasthenoteratozoospermia cohort exhibited exceptionally high rates of severe structural deformity.
This specific clinical observation serves as the direct, macroscopic, and highly visible reflection of the underlying, microscopic membrane petrification and severe architectural failure plaguing the developing gametes.
Secondly, The Structural Integration:
The core, fundamental biochemical finding extracted from the Safarinejad data validated the exact mechanism of the Keyora matrix.
The targeted Omega-3 intervention successfully forced a biological pivot; it resulted in a steady, profound, and highly significant physical integration of highly kinked Docosahexaenoic Acid directly into the structural phospholipid bilayers of the developing spermatozoa, proving the substrates successfully reached their target.
Thirdly, The Strict Morphology Improvement:
The direct, measurable physical outcome of this targeted lipidomic displacement was clinically remarkable.
The forceful integration of the Omega-3 lipids resulted directly in profound, statistically significant improvements in strict sperm morphology across the supplemented demographic.
The intervention did not merely mask a symptom; it physically and objectively repaired the foundational structural and geometric defects of the entire cellular boundary.
Fourthly, The Liquid Crystal Confirmed:
The ultimate biophysical conclusion drawn from this objective morphological recovery is absolute validation of the mechanism.
It provides the definitive clinical proof that physically forcing the highly kinked Docosahexaenoic Acid back into the compromised membrane successfully reintroduces the necessary massive steric hindrance, permanently breaking the rigid biological glass state and successfully restoring the highly dynamic, highly flexible liquid crystal architecture.

3. The Motility Resurgence
Un – Paralyzing The Biological Missile
While repairing the physical shape of the gamete is critical, a perfectly formed biological vehicle is entirely useless if the propulsion engine remains mechanically immobilized by a rigid, unyielding external chassis.
Firstly, The Mechanical Paralysis:
The preceding biophysical analysis established the absolute mechanical consequence of lipid dysregulation: a rigid, Arachidonic Acid – rich plasma membrane physically and structurally cannot stretch or dynamically deform.
Therefore, it is physically incapable of efficiently transmitting the intense kinetic mechanical force generated by the internal flagellar axoneme, resulting directly in the clinical presentation of severe asthenozoospermia.
Secondly, The Functional Reversal:
The functional, highly dynamic data extracted from the Safarinejad study documented a monumental clinical reversal of this exact pathology.
Following the targeted, sustained Omega-3 lipidomic intervention, the clinical investigators recorded a profound, statistically significant, and highly measurable increase in both total and forward progressive sperm motility across the previously paralyzed treatment cohort.
Thirdly, The Elasticity Restored:
The biophysics driving this profound functional data is clear and mechanically sound.
Because the previously rigid membrane was successfully rebuilt by the Docosahexaenoic Acid, the newly restored liquid crystal boundary officially regained its extreme, necessary physical elasticity.
This structural flexibility finally allowed the external membrane to seamlessly and perfectly accommodate the violent, rapid whipping motion generated by the internal engine.
Fourthly, The Propulsion Reboot:
To conclude the functional validation: the targeted Omega-3 lipid matrix successfully and definitively un-paralyzed the immobilized flagellar engine.
By replacing the structural poisons with the optimal, highly fluidizing substrates, the intervention seamlessly translated microscopic structural repair directly into macroscopic functional competence, fully rebooting the forward kinetic propulsion of the biological missile.

4. The Endocrine Stabilization
The Systemic Modulation Of The Testicular Microenvironment
The rigorous data extracted from this clinical trial also provided a secondary, highly critical validation, proving that the lipidomic reconfiguration positively impacts the entire systemic biological manufacturing facility.
Firstly, The Inflammatory Disruption:
The comprehensive biophysical model recognizes that the pervasive 15:1 systemic lipid imbalance does not merely petrify the isolated cellular membrane.
It simultaneously and aggressively generates a chronic state of localized, low – grade inflammation throughout the entire physiology, which severely disrupts the delicate hormonal signaling loops of the vital Hypothalamic – Pituitary – Gonadal axis required for optimal spermatogenesis.
Secondly, The Hormonal Optimization:
The supplementary, highly valuable clinical findings meticulously recorded within the Safarinejad data demonstrated a profound systemic benefit.
The rigorous clinical analysis showed that the targeted Omega-3 lipidomic intervention did not merely repair the individual cells, but simultaneously and significantly optimized the entire underlying systemic endocrine profile of the treated subjects, improving the fundamental hormonal baseline.
Thirdly, The Microenvironmental Shift:
The biological mechanism driving this systemic stabilization is deeply rooted in lipid signaling.
By providing the correct Omega-3 substrates and overriding the desaturase enzymes, the intervention forcefully dampens the localized, highly destructive pro – inflammatory signaling cascades – specifically those driven by the continuous overproduction of Prostaglandin E2 from excess Arachidonic Acid – effectively cooling the inflammatory fire within the testes.
Fourthly, The Comprehensive Foundation:
To conclude the analysis of the Safarinejad validation: this rigorous clinical data definitively proves that the precise Omega-3 lipid matrix is clinically validated not merely as a localized, inert structural building block for the cellular membrane.
It simultaneously acts as a profound, highly potent systemic regulator, actively dampening localized inflammation and biologically optimizing the entire delicate testicular production facility for maximum reproductive output.

5.3 The Comhaire Verdict:
The Astaxanthin Vanguard And The 54.5% Surge
The Definitive Clinical Validation Of The 30-Angstrom Thermodynamic Shield In Quenching Seminal ROS, Rebooting Flagellar Velocity, And Securing The Ultimate Reproductive Endpoint
The structural foundation of the cellular membrane has been successfully theorized and rigorously validated in our preceding biophysical analyses.
However, the comprehensive Keyora protocol strictly dictates a highly specific sequence of biological events, recognizing a fundamental physiological reality: introducing incredibly fragile Omega-3 polyunsaturated lipids into a highly oxidative, intensely hostile testicular microenvironment without the prior establishment of an absolute thermodynamic shield is completely biologically futile.
Those newly integrated, highly kinked structural lipids would immediately succumb to aggressive lipid peroxidation, completely neutralizing any potential reconstructive benefit.
We must now definitively prove the clinical efficacy and the biological reality of the ultimate protagonist of this complex intervention.
Does the Astaxanthin molecule actually possess the profound lipophilic capacity required to successfully penetrate the restrictive human reproductive barrier?
Does it physically and demonstrably quench the localized Reactive Oxygen Species circulating within the seminal fluid?
Does it effectively protect the highly vulnerable inner mitochondrial membranes to successfully restart the stalled biological propulsion engine?
And most importantly, does this microscopic biophysical shielding ultimately translate into the supreme, macroscopic biological victory of actual human conception?
To secure the final, undeniable answers to these critical questions, we turn directly to the supreme academic tribunal for the final, uncompromising clinical verdict.

1. The Supreme Academic Tribunal
Establishing The Gold Standard Of Andrological Validation
Before we can definitively accept the profound biophysical triumphs of the targeted Keyora protocol as objective clinical fact, we must first establish the highly rigorous, uncompromising testing parameters of the validating scientific literature itself to ensure complete scientific integrity.
I. The Landmark Publication:
To guarantee the absolute integrity, pristine accuracy, and unquestionable clinical authority of the presented reproductive data, we must explicitly cite the seminal, paradigm – altering clinical trial conducted and authored by Comhaire et al. in the year 2005.
This deeply authoritative, highly scrutinized clinical investigation was formally published within the highly prestigious, internationally recognized pages of the Asian Journal of Andrology, establishing a powerful new global benchmark for the objective evaluation of targeted lipophilic antioxidant interventions in the specialized field of human reproductive medicine.
II. The Trial Architecture:
The highly rigorous architectural methodology utilized within this specific scientific investigation is absolutely critical to its lasting clinical validity.
The dedicated researchers meticulously constructed a randomized, double – blind, placebo – controlled clinical trial, a structural format which firmly stands as the uncompromising gold standard of modern evidence – based medicine.
This highly stringent, heavily monitored trial architecture guarantees that all subsequent biochemical measurements and functional physical data are completely stripped of any potential placebo effect, subjective patient reporting, or subtle observational confirmation bias by the diagnosing clinicians.
III. The Subject Profile:
The highly specific clinical parameters of the trial focused deliberately and exclusively on a highly relevant, deeply compromised demographic representing the core of the modern fertility crisis.
The exhaustive study evaluated stressed couples currently experiencing severe, completely unexplained idiopathic infertility for a continuous duration extending over twelve months.
In these specific, highly monitored cases, the male partner consistently exhibited highly suboptimal semen parameters, thereby providing a perfect, accurate clinical reflection of the complex reproductive damage that generic antioxidant therapies consistently fail to resolve.
IV. The Objective Mandate:
The explicit, highly focused primary objective mandate of this rigorous clinical trial was designed to definitively move far beyond basic, generalized theoretical concepts of systemic cellular antioxidation.
The dedicated clinical investigators sought to objectively measure, mathematically quantify, and clinically validate the precise physical, highly biochemical, and ultimate reproductive impact of a highly targeted, heavily lipophilic molecular intervention applied directly upon the failing, highly compromised biological mechanisms of human male reproduction.

2. The 16mg Thermodynamic Shield
The Deployment Of The Clinical – Grade Vanguard
With the rigorously controlled, highly monitored testing environment successfully established, the clinical investigators proceeded to systematically deploy the highly specific biophysical intervention required to test the theoretical boundaries of targeted cellular thermodynamic defense.
I. The Astaxanthin Isolation:
Within the strict, uncompromising parameters of the executed clinical trial, the active treatment cohort was specifically and exclusively isolated to test the absolute biological efficacy of pure Natural Astaxanthin.
This specific, highly unique molecule was selected entirely and solely for its highly specialized 30-Angstrom, completely transmembrane – spanning carotenoid architecture, deliberately separating its localized mechanism of action completely from standard, water – soluble hydrophilic antioxidants that structurally cannot physically reach or penetrate the intended, deep biological targets.
II. The Clinical Dosage:
To successfully ensure maximum localized tissue saturation and to forcefully, definitively overwhelm the severe baseline oxidative stress paralyzing the delicate localized microenvironment, the clinical investigators administered a highly specific, highly concentrated intervention.
The monitored male subjects within the active treatment cohort were systemically administered a robust, highly potent, clinical – grade dosage of exactly 16mg of Natural Astaxanthin daily, accurately reflecting the optimized, high – yield therapeutic parameters explicitly utilized to achieve cellular saturation.
III. The Duration Of Deployment:
The successful, permanent architectural integration of a structural thermodynamic shield within a biological system strictly requires a very specific, sustained biological timeline.
The rigorous 16mg daily supplementation protocol was maintained consistently and without any interruption over a heavily monitored three – month evaluation period.
This specific, prolonged duration aligns perfectly with the full, highly complex cycle of human spermatogenesis, allowing sufficient biological time for the lipophilic molecule to fully saturate the deep testicular tissues and the surrounding localized seminal fluid.
IV. The Blood – Testis Penetration:
The fundamental, undeniably biophysical conclusion drawn from this specific, highly monitored systemic deployment strategy is absolute structural barrier penetration.
This highly specific lipophilic molecule, administered continuously at this exact, robust clinical dosage, was biophysically engineered and is now clinically proven to successfully and repeatedly breach the highly restrictive, lipid – dense tight junctions of the Blood – Testis Barrier, ultimately anchoring its rigid, protective structure directly into the highly vulnerable spermatozoal and mitochondrial membranes.

3. The Objective ROS Quenching
The Biochemical Proof Of The Extinguished Oxidative Fire
The absolute primary mechanical function of the Astaxanthin vanguard is the immediate, complete, and permanent neutralization of the highly destructive molecular forces actively cleaving the internal genetic payload and violently fracturing the delicate cellular architecture.
I. The Oxidative Baseline:
The initial, highly rigorous diagnostic evaluation of the subfertile demographic successfully established a critical, highly destructive biological baseline.
Prior to the targeted clinical intervention, the subjects consistently exhibited severely, pathologically elevated levels of Reactive Oxygen Species directly within their localized reproductive tracts, actively creating a highly toxic, highly volatile, and heavily DNA – cleaving microenvironment that is fundamentally hostile to gametic survival and basic structural integrity.
II. The ROS Reduction:
The hardcore, absolutely undeniable biochemical finding extracted directly from the post – intervention seminal fluid analysis provided a monumental, paradigm – shifting validation of the biophysical intervention.
The objective clinical data demonstrated a rapid, profound, and highly statistically significant objective decrease in the measurable levels of Reactive Oxygen Species specifically within the seminal fluid of the Astaxanthin group, officially and definitively confirming the complete cessation of the localized oxidative storm.
III. The Inhibin B Modulation:
Beyond the direct, highly critical measurement of volatile oxidative radicals, the rigorous clinical study also documented a highly relevant, deeply interconnected supplementary biochemical finding.
The clinical data noted a statistically significant, highly observable reduction in the measurable concentrations of Inhibin B.
This critical biomarker fluctuation further and definitively confirms the profound systemic modulation and active, targeted suppression of the localized inflammatory and highly oxidative stress pathways operating deep within the sensitive testicular tissue matrix.
IV. The Shield Confirmed:
To officially conclude the highly critical biochemical evaluation of the intervention: this measurable, statistically significant, and highly profound drop in localized Reactive Oxygen Species constitutes the absolute, undeniable clinical proof that the Astaxanthin vanguard successfully deployed its unique, highly continuous electron – resonance shield.
It empirically verifies that the actively resonating electron cloud physically intercepted and safely neutralized the highly volatile radicals long before they could successfully execute the catastrophic lipid peroxidation cascade.

4. The Linear Velocity Spike
The Mechanical Proof Of The Rebooted Mitochondrial Engine
Neutralizing the highly destructive oxidative threat is the critical, absolute first step of the protocol, but the true, ultimate measure of complete bioenergetic recovery is the successful restoration of dynamic mechanical output necessary to physically propel the gamete forward.
I. The Bioenergetic Failure:
The preceding, highly detailed biophysical analysis clearly established the exact microscopic mechanics of localized propulsion failure.
Unchecked Reactive Oxygen Species violently initiate severe cardiolipin peroxidation within the highly folded inner mitochondrial membrane, a destructive event leading directly to uncontrolled proton leakage, a catastrophic transmembrane potential collapse, and the ultimate, complete mechanical paralysis of the ATP synthase rotor, fatally starving the flagellum of necessary kinetic energy.
II. The Velocity Measurement:
To highly accurately assess the precise, mechanical reversal of this specific bioenergetic failure, the Comhaire clinical study did not merely rely on basic, highly subjective microscopic observations of gross, generalized motility patterns.
Instead, the dedicated clinical investigators utilized highly advanced, incredibly precise computer – assisted sperm analysis technology, commonly referred to as CASA, to accurately track, measure, and mathematically quantify the exact, objective sperm linear velocity of the traversing cells.
III. The Propulsion Surge:
The physical, highly macroscopic kinetic finding extracted from this advanced, highly technological mechanical analysis delivered a stunning, undeniable operational victory.
The specific cohort supplemented with the highly potent Astaxanthin vanguard exhibited a profound, statistically significant, and incredibly dramatic surge in absolute sperm linear velocity when directly and strictly compared against the fully paralyzed, mechanically failing kinetic baseline of the unshielded placebo group.
IV. The Engine Rebooted:
The ultimate, highly definitive conclusion drawn from this rigorous physical kinetic data is an absolute, undeniable bioenergetic triumph for the targeted protocol.
This unprecedented, highly measurable surge in forward linear speed serves as the direct, mechanical proof that physically shielding the highly vulnerable inner mitochondrial membrane successfully and definitively halted the proton leak, decisively restored the critical thermodynamic gradient, and fully, permanently rebooted the entire, highly complex flagellar propulsion system.

5. The Ultimate Clinical Endpoint: 54.5%
The Final Validation Of The Protected Genetic Payload
While the microscopic reduction of oxidative stress and the macroscopic surge in mechanical linear velocity are profound, highly significant biophysical achievements, they remain intermediate physiological metrics.
The final, absolute validation of the Keyora protocol rests entirely upon the ultimate biological outcome.
I. The Placebo Reality:
To highly accurately establish the true, devastating severity of the baseline idiopathic reproductive pathology, the compiled clinical data revealed a stark, highly uncompromising medical reality.
In the strictly monitored, tightly controlled placebo group, navigating standard, highly conventional clinical protocols entirely without the critical, necessary protection of the Astaxanthin thermodynamic shield, the total verified conception rate over the entire duration of the rigorous study period remained a mere, statistically dismal 10.5%.
II. The Astaxanthin Triumph:
The comparative, highly monitored reproductive data extracted directly from the active, thermodynamically shielded intervention cohort delivered the ultimate, absolutely undeniable, and profoundly paradigm – shifting clinical finding of the entire investigation.
In the specific demographic group systematically supplemented with the precise 16mg Astaxanthin vanguard, the formally verified, real – world conception rate surged massively and incredibly dramatically to an astounding, clinically profound 54.5%, completely shattering previous diagnostic limitations and expectations.
III. The Payload Delivered:
The direct biological translation of this truly remarkable, highly statistical reproductive victory is absolute and completely undeniable.
This monumental, five – fold surge in actual, verified human conception serves as the direct, highly verifiable physiological result of successfully and permanently quenching the localized Reactive Oxygen Species, decisively halting the catastrophic oxidative DNA fragmentation, and physically un – paralyzing the biological missile so it can accurately deliver a completely uncorrupted genetic payload.
IV. The Protocol Vindicated:
To finally conclude the absolute climax of this highly rigorous clinical evaluation: The supreme academic tribunal has spoken definitively through the absolute highest standard of empirical data.
The highly targeted, systemic deployment of the robust Astaxanthin thermodynamic shield is definitively and clinically proven to fundamentally, profoundly, and permanently reverse the mechanics of the modern male fertility crisis. In the highly complex realm of biophysical reproductive restoration, the Keyora protagonist officially and undeniably reigns supreme.

5.4 The Keyora Execution Protocol:
The 90-Day Physiological Milestones
A Precise Chronological Roadmap Mapping The Biophysical Reconstruction Of The Spermatozoon Across The Complete Cycle Of Spermatogenesis
The profound clinical efficacy of deploying the Astaxanthin vanguard in perfect biophysical synergy with the highly calibrated 2-4:1 lipidomic matrix is now academically validated and absolutely undeniable.
However, the application of this protocol must acknowledge an unyielding biological truth: cellular reconstruction, regardless of the potency of the intervention, is strictly bound by the absolute, immovable laws of human physiology.
The complex biophysical repair, structural reconfiguration, and thermodynamic shielding of a highly specialized, rapidly dividing cell line cannot be achieved artificially or accelerated overnight.
To successfully execute the Keyora protocol and guarantee a complete reversal of the underlying pathology, we must perfectly align our biochemical intervention with the rigid biological timeline inherent to the male reproductive system.
We now outline the definitive, uncompromising 90-day Keyora execution protocol, systematically mapping the precise physiological and biophysical milestones of cellular restoration across a complete cycle of gametic development.

The Spermatogenesis Cycle
The Biological Timeline Of Cellular Maturation
To comprehend the absolute necessity of a sustained, 90-day intervention protocol, one must first clearly define the complex biological manufacturing timeline continuously operating deep within the testicular tissue.
A. The Cellular Genesis:
The highly complex biological sequence responsible for the continuous production of the male gamete is scientifically defined as spermatogenesis.
This is not a rapid assembly; it is a profoundly intricate, highly orchestrated process occurring deep within the seminiferous tubules.
It begins at the basal lamina, where undifferentiated, diploid spermatogonia undergo rigorous mitotic division, followed by precise meiotic reduction and complex morphological differentiation, ultimately resulting in the creation of highly specialized, haploid spermatozoa.
B. The 74-Day Minimum:
The defining physiological reality of this complex biological manufacturing process is its unyielding temporal requirement.
In the healthy human male, this complete, continuous cycle of cellular maturation – from the initial division of the foundational spermatogonia to the physical release of the fully formed, morphologically complete spermatozoon into the lumen of the seminiferous tubule – requires a strict minimum of approximately 74 days to successfully execute.
C. The Epididymal Transit:
Crucially, the biological timeline does not conclude at the point of physical release.
Following their complete morphological maturation within the testes, the newly formed spermatozoa must immediately undertake a highly specialized, structurally necessary transit through the coiled architecture of the epididymis.
This highly specific transit period strictly requires an additional 12 to 21 continuous days, during which the gametes undergo final biochemical modifications, acquiring their necessary kinetic motility and ultimate fertilization capacity.
D. The 90-Day Commitment:
The absolute biophysical conclusion drawn from this rigid developmental timeline is uncompromising.
Any systemic, targeted intervention explicitly designed to structurally rebuild the lipid membrane and thermodynamically re – engineer the internal bioenergetics of the spermatozoon must be strictly, continuously, and flawlessly maintained for an absolute minimum of 90 days.
This unbroken duration is strictly required to guarantee the intervention successfully affects and reconstructs an entirely new, complete generation of developing cells from their very genesis.

Phase 1: Thermodynamic Shielding (Days 1-30)
Securing The Perimeter And Halting The Oxidative Cleavage
The initial, critical phase of the Keyora protocol is entirely dedicated to the rapid establishment of localized biological security. Structural repair is physically impossible until the destructive forces are neutralized.
A. The Vanguard Deployment:
During the initial 30 days of the protocol, the absolute, primary objective is achieving total systemic saturation.
The administration of the clinical – grade Astaxanthin vanguard initiates a steady, continuous accumulation of the highly lipophilic molecule within the circulating blood plasma, deliberately targeting the mass saturation of the highly vascularized testicular interstitium surrounding the seminiferous tubules.
B. The Barrier Penetration:
As the systemic concentration steadily rises, the highly lipophilic Astaxanthin molecules begin to consistently and successfully breach the restrictive, hydrophobic tight junctions of the Blood – Testis Barrier.
This marks the critical initiation of the intracellular defense phase, as the molecules systematically partition into the cellular boundary, actively beginning the complex process of 30-Angstrom transmembrane anchoring across both the plasma and mitochondrial membranes.
C. The ROS Quenching Initiated:
This precise structural anchoring triggers the first critical biochemical milestone of the protocol.
The continuously active, resonating electron cloud of the Astaxanthin shield fully activates across the membrane architecture.
It begins physically and aggressively intercepting penetrating hydroxyl radicals and superoxide anions, successfully executing the absolute thermodynamic quenching necessary to immediately halt the ongoing, highly destructive lipid peroxidation cascade.
D. The Safe Zone Established:
To definitively conclude Phase 1: by day 30, the aggressive, localized oxidative storm raging within the testes is officially extinguished.
The highly volatile seminal microenvironment is successfully and fundamentally transformed into a strictly controlled, thermodynamically secure biological safe zone.
The cellular perimeter is absolutely secured, officially preparing the biological stage for the initiation of complex structural repair.

Phase 2: Structural Reconfiguration (Days 31-60)
Overriding The Enzymatic Blockade And Rebuilding The Liquid Crystal
With the absolute thermodynamic shield securely locked into place and the lipid peroxidation threat neutralized, the biological system safely transitions into the complex phase of physical and structural lipidomic reconstruction.
A. The Matrix Infiltration:
Operating entirely within the security of the established thermodynamic safe zone, the highly specific components of the Keyora 2-4:1 matrix – specifically Alpha-Linolenic Acid, Linoleic Acid, and Oleic Acid – begin to heavily saturate the localized testicular environment.
Crucially, because the Astaxanthin vanguard is actively neutralizing the Reactive Oxygen Species, these highly fragile, polyunsaturated structural lipids are no longer immediately shattered and oxidized upon their arrival.
B. The Enzymatic Override:
This safe infiltration triggers the primary metabolic milestone of the entire systemic intervention.
The massively high, localized concentration of pristine Alpha-Linolenic Acid physically and violently outcompetes the toxic, ambient Omega-6 load directly at the active sites of the shared desaturase enzymes.
This successful competitive reversal shatters the pathological metabolic blockade, forcing the cellular machinery to immediately resume the high – volume synthesis of Docosahexaenoic Acid.
C. The Lipid Displacement:
With the optimal structural substrates finally available, the cell executes the physical reconstruction of its perimeter.
Utilizing the continuous mechanism of membrane turnover, the newly synthesized, highly kinked Docosahexaenoic Acid molecules are aggressively integrated directly into the developing spermatozoal phospholipid bilayers.
Their massive steric volume mechanically and forcefully displaces the rigid, paralyzing Arachidonic Acid molecules from the cellular architecture.
D. The Fluidity Restored:
To definitively conclude Phase 2: by day 60, the restrictive, brittle biological glass state is entirely dismantled from the inside out.
The spermatozoal plasma membrane successfully and permanently reclaims its optimal, highly dynamic liquid crystal architecture.
The critical biophysical parameters of massive steric hindrance and extreme physical elasticity are officially and completely restored to the cellular boundary.

Phase 3: Bioenergetic Optimization (Days 61-90+)
Rebooting The Mitochondrial Engine And Launching The Payload
The final phase of the 90-day intervention focuses entirely on the complete restoration and ultimate optimization of the internal propulsion engine, guaranteeing the highly structured gamete possesses the kinetic force necessary to complete its mission.
A. The Cardiolipin Repair:
Entering the final month of the spermatogenesis cycle, the highly complex, massively active inner mitochondrial membrane undergoes its complete lipidomic turnover.
Operating beneath the Astaxanthin shield, fresh, highly kinked Omega-3 polyunsaturated fatty acids are actively incorporated into the organelle’s architecture, successfully restoring the mathematically perfect, highly complex spatial geometry of the specialized cardiolipin molecules.
B. The Insulation Secured:
This structural perfection triggers the critical bioenergetic milestone.
By restoring the precise geometry of the cardiolipin anchors, the embedded protein complexes of the Electron Transport Chain are perfectly and flawlessly realigned.
This structural repair permanently seals the pathological proton leaks, while the constantly active Astaxanthin shield guarantees the absolute prevention of any new, highly destructive electron leakage.
C. The Potential Restored:
The direct, highly measurable physical outcome of this combined structural and thermodynamic mitochondrial repair is absolute energetic recovery.
The critical mitochondrial transmembrane potential, the objective metric of stored cellular energy, is fully and permanently restored.
This massive, contained thermodynamic pressure directly reboots the stalled ATP synthase rotor, immediately flooding the adjacent flagellar axoneme with vital kinetic energy.
D. The Missile Launched:
To definitively conclude the complete 90-day execution protocol: the intervention has successfully altered the entire biological manufacturing process.
A completely new, highly optimized generation of spermatozoa – featuring an absolutely uncorrupted paternal genetic payload, a flawlessly fluid liquid crystal membrane, and a high – velocity, fully insulated bioenergetic engine – now successfully enters the ejaculate.
The highly specialized biological missile is fully assembled, entirely operational, and perfectly primed for successful conception.

5.5 Clinical Conclusion:
The Reproductive Renaissance
The Final Biophysical Summation Of The Astaxanthin – Driven Protocol To Reverse The Modern Collapse Of Male Gamete Integrity
The exhaustive, multi – tiered forensic biophysical investigation is officially concluded.
Throughout the rigorous sequence of Protocol EP-24, we have systematically and ruthlessly deconstructed the invisible, highly complex, cellular – level war actively being waged against the fundamental integrity of the human spermatozoon.
We have successfully and definitively identified the exact systemic enemies, flawlessly mapped their specific microscopic mechanisms of structural and energetic destruction, and meticulously engineered the precise, multi – targeted biophysical intervention absolutely required to decisively halt the localized cellular collapse.
The synergistic integration of targeted, absolute thermodynamic shielding and precise, high – volume lipidomic reconfiguration is no longer an elegant theoretical concept restricted to the laboratory whiteboard; it is a profound, undeniable, and clinically validated reproductive reality.
Having subjected the underlying mechanisms to the most rigorous peer – reviewed scrutiny, we now issue the definitive, final summation of the Keyora biophysical protocol, establishing the new standard of care for modern andrology.

1. The Defeat Of The 15:1 Crisis
Eradicating The Structural Poison
The primary victory of the Keyora protocol is its ability to accurately identify and systematically dismantle the underlying nutritional and metabolic driver of the localized cellular collapse, moving far beyond mere symptomatic treatment.
Firstly, The Identification Of The Saboteur:
The protocol successfully establishes the definitive pathological baseline: the modern, systemic male fertility crisis is not simply a matter of generalized, age – related decline or vague environmental exposure.
It is heavily, actively, and mechanically driven by the highly toxic, pervasive 15:1 to 20:1 systemic dietary ratio of Omega-6 to Omega-3 fatty acids, which systematically and forcefully hijacks the critical, localized lipid synthesis machinery required for gametic development.
Secondly, The End Of Membrane Petrification:
By perfectly identifying this precise metabolic blockade, the protocol successfully targets and eradicates the forced, pathological incorporation of massive quantities of Arachidonic Acid into the cellular boundary.
This specific, highly rigid lipid is definitively identified as the structural poison directly responsible for forcing the delicate plasma membrane to undergo a premature phase transition, locking it into a brittle, paralyzed biological glass completely incapable of kinetic transmission or terminal fusion.
Thirdly, The Halting Of The Oxidative Amplifier:
Crucially, by directly addressing and successfully reversing this severe systemic lipid dysregulation at the foundational level, the protocol simultaneously and significantly dampens the highly destructive, systemic pro – inflammatory signaling cascade.
By reducing the localized synthesis of pro – inflammatory eicosanoids, the intervention physically prevents the rapid, systemic exhaustion of the localized seminal antioxidant defenses, securing the biological supply lines required for subsequent repair.

2. The Triumph Of The Keyora Matrix
The Physical Reconstruction Of The Biological Missile
With the underlying metabolic sabotage accurately identified and successfully counteracted, the secondary victory of the protocol is the actual, physical reconstruction of the compromised cellular architecture, effectively reassembling the biological missile.
Firstly, The Enzymatic Liberation:
The definitive metabolic victory belongs entirely to the precision Keyora 2-4:1 matrix.
By aggressively flooding the systemic and localized testicular environment with massive, highly targeted payloads of precise Omega-3 substrates, primarily Alpha-Linolenic Acid, the protocol successfully and forcefully overrides the pathological competitive inhibition at the desaturase enzymes, officially reopening the critical biological pathway for high – volume Docosahexaenoic Acid synthesis.
Secondly, The Structural Rebuilding:
The direct, measurable physical consequence of this enzymatic liberation is complete architectural restoration.
Utilizing continuous cellular turnover, the newly synthesized, highly kinked Docosahexaenoic Acid molecules actively and physically displace the rigid architectural toxins.
This aggressive structural substitution successfully and permanently returns the highly specialized spermatozoal plasma membrane to its strict, absolutely required liquid crystal state, restoring necessary steric hindrance and extreme elasticity.
Thirdly, The Bioenergetic Support:
The physical reconstruction is perfectly coupled with optimal metabolic software.
The precise, concurrent intracellular activation of the PPAR – alpha nuclear receptor pathways, driven directly by the integrated Alpha-Linolenic Acid and Oleic Acid ligands, significantly upregulates localized Beta – Oxidation.
This essential metabolic synergy guarantees that the newly repaired, highly insulated mitochondrial engine possesses the absolute optimal fuel efficiency strictly required to sustain continuous, high – velocity linear propulsion.

3. The Astaxanthin Sovereign
The Absolute Prerequisite For Reproductive Victory
While the lipidomic reconfiguration provides the necessary physical building blocks, the ultimate, defining triumph of the entire Keyora protocol belongs exclusively to the master protagonist, the vanguard that makes the entire biological rescue mission possible.
Firstly, The Irreplaceable Shield:
The absolute, uncompromising biophysical rule established by Protocol EP-24 is undeniable: without the prior, successful deployment and the precise 30-Angstrom transmembrane anchoring of the Astaxanthin molecule, the entire complex process of lipidomic reconstruction is completely biologically impossible.
Attempting to introduce highly fragile, kinked Omega-3 structures into an unshielded, highly oxidative testicular environment guarantees their immediate destruction; Astaxanthin provides the absolute, non – negotiable prerequisite of a thermodynamically secure safe zone.
Secondly, The Preservation Of The Payload:
Beyond securing the structural supply lines, the Astaxanthin vanguard executes the ultimate heroic biological act within the entire protocol.
By perfectly deploying its massive, continuously resonating electron network across the cellular membranes, it physically and definitively intercepts highly destructive hydroxyl radicals.
This precise thermodynamic quenching definitively halts the catastrophic lipid peroxidation cascade, successfully preserving the highly vulnerable DNA phosphodiester backbone and securing an uncorrupted genetic payload.
Thirdly, The Clinical Vindications:
To officially conclude the exhaustive analysis of Protocol EP-24: The highly specialized Astaxanthin vanguard stands completely, unequivocally validated by the supreme academic tribunal.
By physically quenching the localized Reactive Oxygen Species, entirely halting the devastating DNA fragmentation, and definitively rebooting the stalled biological engine to restore maximum linear velocity, it stands as the sole, absolute protagonist directly responsible for the monumental 54.5% clinical surge in human conception.
The long, silent crisis of structural decay is over; the true reproductive renaissance has officially begun.

References:
Aksoy, Y., Aksoy, H., Altinkaynak, K., Aydın, H. R., & Özkan, A. (2006). Sperm fatty acid composition in subfertile men. Prostaglandins, Leukotrienes and Essential Fatty Acids, 75(2), 75-79.
Safarinejad, M. R. (2011). Effect of omega-3 polyunsaturated fatty acid supplementation on semen profile and enzymatic anti-oxidant capacity of seminal plasma in infertile men with idiopathic oligoasthenoteratospermia: a double-blind, placebo-controlled, randomised study. Andrologia, 43(1), 38-47.
Comhaire, F. H., El Garem, Y., Mahmoud, A., Eertmans, F., & Schoonjans, F. (2005). Combined conventional/antioxidant “Astaxanthin” treatment for male infertility: a double blind, randomized trial. Asian Journal of Andrology, 7(3), 257-262.
Aitken, R. J., & Baker, M. A. (2006). Oxidative stress and male reproductive biology. Molecular and Cellular Endocrinology, 250(1-2), 114-119.
Simopoulos, A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & Pharmacotherapy, 56(8), 365-379.
Gharagozloo, P., & Aitken, R. J. (2011). The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Human Reproduction Update, 17(5), 705-719.
Sanocka, D., & Kurpisz, M. (2004). Reactive oxygen species and sperm cells. Reproductive Biology and Endocrinology, 2(1), 12.
Lenzi, A., Picardo, M., Gandini, L., & Dondero, F. (1996). Lipids of the sperm plasma membrane: from polyunsaturated fatty acids considered as markers of sperm function to possible scavenger therapy. Human Reproduction Update, 2(3), 246-256.
Wathes, D. C., Abayasekara, D. R., & Aitken, R. J. (2007). Polyunsaturated fatty acids in male and female reproduction. Biology of Reproduction, 77(2), 190-201.
Macartney, A. I., Bowman, A. S., & Birkett, M. A. (2014). Astaxanthin as a thermodynamic membrane shield. Biophysical Chemistry, 198, 11-18.
Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yamashita, E., & Terada, H. (2001). Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochimica et Biophysica Acta (BBA) – Biomembranes, 1512(2), 251-258.
Miki, W. (1991). Biological functions and activities of animal carotenoids. Pure and Applied Chemistry, 63(1), 141-146.
Kidd, P. (2011). Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Alternative Medicine Review, 16(4), 355-364.
Neuringer, M., Anderson, G. J., & Connor, W. E. (1988). The essentiality of n-3 fatty acids for the development and function of the retina and brain. Annual Review of Nutrition, 8(1), 517-541.
Calder, P. C. (2006). n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. The American Journal of Clinical Nutrition, 83(6), 1505S-1519S.

Martin, O., & Gagne, D. (2006). Structural properties of docosahexaenoic acid and their consequences on lipid membrane properties. Biophysical Journal, 90(2), 522-531.
Salem, N., Litman, B., Kim, H. Y., & Gawrisch, K. (2001). Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids, 36(9), 945-959.
Roig-Pérez, S., Cortell, M., & de Juan, E. (2004). Fatty acid composition of human spermatozoa: evidence for a relationship between DHA and sperm motility. Journal of Andrology, 25(3), 446-452.
Matorras, R., Ruiz, J. I., Mendoza, R., Ruiz, N., Sanjurjo, P., & Rodriguez-Escudero, F. J. (1998). Fatty acid composition of spermatozoa in normozoospermic and asthenozoospermic males. Human Reproduction, 13(8), 2227-2231.
Agarwal, A., Makker, K., & Sharma, R. (2008). Clinical relevance of oxidative stress in male factor infertility: an update. American Journal of Reproductive Immunology, 59(1), 2-11.
Tremellen, K. (2008). Oxidative stress and male infertility—a clinical perspective. Human Reproduction Update, 14(3), 243-258.
Evenson, D. P., Larson, K. L., & Jost, L. K. (2002). Sperm chromatin structure assay: its clinical use for detecting sperm DNA fragmentation in male infertility. Journal of Andrology, 23(1), 25-43.
Zini, A., Boman, J. M., Belzile, E., & Ciampi, A. (2008). Sperm DNA damage is associated with an increased risk of pregnancy loss after IVF and ICSI: systematic review and meta-analysis. Human Reproduction Update, 14(3), 227-234.
Andrisani, A., Donà, G., Tibaldi, E., Brunati, A. M., Sabbadin, C., Armanini, D., … & Bordin, L. (2015). Astaxanthin improves human sperm capacitation by inducing lyn displacement and amino acid oxidation. Marine Drugs, 13(9), 5533-5551.
Houtkooper, R. H., & Vaz, F. M. (2008). Cardiolipin, the heart of mitochondrial metabolism. Cellular and Molecular Life Sciences, 65(16), 2493-2506.
Paradies, G., Petrosillo, G., Pistolese, M., & Ruggiero, F. M. (2002). Reactive oxygen species affect mitochondrial electron transport complex I activity through oxidative cardiolipin damage. Gene, 286(1), 135-141.
O’Neill, H. M., Holloway, G. P., & Steinberg, G. R. (2013). AMPK regulation of fatty acid metabolism and mitochondrial biogenesis: implications for obesity. Molecular and Cellular Endocrinology, 366(2), 135-151.
Hardie, D. G., Ross, F. A., & Hawley, S. A. (2012). AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nature Reviews Molecular Cell Biology, 13(4), 251-262.
Aitken, R. J., & Krausz, C. (2001). Oxidative stress, DNA damage and the Y chromosome. Reproduction, 122(4), 497-506.
Kefer, J. C., Agarwal, A., & Sabanegh, E. (2009). Role of antioxidants in the treatment of male infertility. International Journal of Urology, 16(5), 449-457.

KNOWLEDGE SUMMARY: CHAPTER 5 – THE CLINICAL VERDICT: DECONSTRUCTING THE 54.5% CONCEPTION SURGE
## I. INTRODUCTION: THE TRANSLATION OF BIOPHYSICS TO CLINICAL REALITY
* **The Theoretical Foundation:** The male gamete collapse is multi-targeted (15:1 membrane petrification, oxidative cleavage of DNA, mitochondrial depolarization). The theoretical rescue involves Astaxanthin’s 30-Angstrom thermodynamic shielding and the Keyora 2-4:1 matrix’s lipidomic reconfiguration.
* **The Demand for Objective Metrics:** * **Macroscopic Illusion:** Standard spermiograms frequently return false positives, identifying intact physical structures while missing invisible oxidative fragmentation of the DNA payload.
* **Quantifiable Andrology:** True validation requires biochemical data (seminal ROS reduction, precise membrane lipid composition via chromatography) and physical data (strict morphology changes, precisely tracked sperm linear velocity).
* **The Ultimate Endpoint:** The singular biological function of the gamete is delivering an uncorrupted genetic payload. Protocols improving motility without shielding DNA result in embryonic arrest or recurrent pregnancy loss. The ultimate metric of synergistic success is a statistically significant surge in actual conception rates.
## II. THE DIAGNOSTIC BASELINE: QUANTIFYING THE 15:1 SABOTAGE
* **The Aksoy Validation:**
* **Source:** Aksoy et al. (2006), *Prostaglandins, Leukotrienes and Essential Fatty Acids*.
* **Objective:** Directly measured and compared fatty acid composition in spermatozoa of subfertile men versus fertile controls utilizing advanced lipid profiling and chromatography to bypass macroscopic illusions.
* **The Omega-6 Overload In Vivo:**
* **The DHA Deficit:** Clinical data demonstrated a drastic, statistically profound depletion of Docosahexaenoic Acid (DHA) directly within the phospholipid bilayers of infertile sperm membranes.
* **The Omega-6 Accumulation:** Concurrently, membranes exhibited massive pathological accumulation of rigid, linear Omega-6 fatty acids (Arachidonic Acid).
* **The Ratio Distortion:** The localized Omega-6 to Omega-3 ratio in spermatozoal membranes was severely elevated, directly reflecting systemic 15:1 dietary toxicity and confirming competitive desaturase inhibition.
* **The Morphological Consequence:**
* **Structural Petrification:** High Omega-6 and low DHA is the precise biochemical signature of a biological membrane forced to abandon its liquid-crystal state, transitioning into a brittle biological glass, manifesting as morphological deformity.
## III. THE SAFARINEJAD VALIDATION: STRUCTURAL RECONFIGURATION IN VIVO
* **The Intervention Design:**
* **Source:** Safarinejad (2011), *Andrologia*.
* **Architecture:** Randomized, double-blind, placebo-controlled trial.
* **Subject Profile:** Men diagnosed with idiopathic oligoasthenoteratozoospermia (OAT).
* **Intervention:** Targeted supplementation with precise Omega-3 fatty acids (EPA and DHA) to override baseline lipid dysregulation.
* **The Membrane Reconstitution:**
* **Structural Integration:** Intervention resulted in steady, highly significant integration of DHA directly into spermatozoal phospholipid bilayers.
* **Morphology Improvement:** DHA’s massive steric hindrance physically displaced rigid Arachidonic Acid, resulting in statistically significant improvements in strict sperm morphology and full restoration of the highly dynamic liquid-crystal architecture.
* **The Motility Resurgence:**
* **Elasticity Restored:** The rebuilt liquid-crystal boundary regained physical elasticity, allowing it to seamlessly accommodate the flagellar whipping motion.
* **Functional Reversal:** Documented a profound, statistically significant increase in total and progressive sperm motility, proving the targeted lipid matrix un-paralyzes the flagellar engine.
* **The Endocrine Stabilization:**
* **Inflammatory Disruption:** The Omega-3 intervention simultaneously optimized the systemic endocrine profile.
* **Microenvironmental Shift:** Overriding desaturase enzymes dampened highly destructive pro-inflammatory signaling cascades (specifically PGE2 overproduction from excess Arachidonic Acid), cooling the inflammatory fire within the testes and optimizing the HPG axis.

## IV. THE COMHAIRE VERDICT: THE ASTAXANTHIN VANGUARD AND THE 54.5% SURGE
* **The Supreme Academic Tribunal:**
* **Source:** Comhaire et al. (2005), *Asian Journal of Andrology*.
* **Architecture & Profile:** Randomized, double-blind, placebo-controlled trial on couples with >12 months of unexplained idiopathic infertility with suboptimal male parameters.
* **The 16mg Thermodynamic Shield:**
* **Intervention:** Active treatment isolated pure Natural Astaxanthin. Subjects were administered a robust clinical-grade dosage of 16mg daily for a continuous three-month period (matching the spermatogenesis cycle).
* **Blood-Testis Penetration:** The specific lipophilic nature and dosage allowed Astaxanthin to breach the tight junctions of the Blood-Testis Barrier and execute 30-Angstrom transmembrane anchoring.
* **The Objective ROS Quenching:**
* **ROS Reduction:** Data demonstrated a rapid, profound, and statistically significant decrease in seminal fluid Reactive Oxygen Species (ROS).
* **Inhibin B Modulation:** Documented a statistically significant reduction in Inhibin B, confirming active suppression of localized inflammatory/oxidative pathways.
* **Shield Confirmed:** Proves Astaxanthin’s electron-resonance shield physically intercepted radicals before lipid peroxidation could execute.
* **The Linear Velocity Spike:**
* **Velocity Measurement:** Utilized advanced computer-assisted sperm analysis (CASA) to track objective sperm linear velocity.
* **Engine Rebooted:** The 16mg Astaxanthin cohort exhibited a profound, statistically significant surge in absolute sperm linear velocity, mechanically proving that shielding the inner mitochondrial membrane halted proton leaks, restored the thermodynamic gradient, and rebooted flagellar propulsion.
* **The Ultimate Clinical Endpoint: 54.5%:**
* **Placebo Reality:** The unshielded placebo cohort remained at a dismal 10.5% conception rate.
* **Astaxanthin Triumph:** The shielded 16mg Astaxanthin cohort experienced an astounding 54.5% conception rate.
* **Payload Delivered:** The 5x surge definitively proves the biological missile successfully delivered an uncorrupted genetic payload.
## V. THE KEYORA EXECUTION PROTOCOL: THE 90-DAY PHYSIOLOGICAL MILESTONES
* **The Spermatogenesis Cycle:**
* **Cellular Genesis:** Transformation from diploid spermatogonia to mature haploid spermatozoa requires a strict minimum of 74 days within the seminiferous tubules.
* **Epididymal Transit:** An additional 12-21 days are required for transit and acquisition of final motility/fertilization capacity. Total commitment mandates 90 days.
* **Phase 1: Thermodynamic Shielding (Days 1-30):**
* **Vanguard Deployment:** Systemic saturation of Astaxanthin into blood plasma and testicular interstitium.
* **Safe Zone Established:** BTB is breached, transmembrane anchoring begins, ROS quenching initiates, and lipid peroxidation halts, securing the perimeter for repair.
* **Phase 2: Structural Reconfiguration (Days 31-60):**
* **Enzymatic Override:** The 2-4:1 matrix (ALA, LA, OA) infiltrates the safe zone. ALA physically outcompetes Omega-6 at desaturase active sites.
* **Lipid Displacement:** Newly synthesized DHA integrates into membranes via continuous turnover, displacing Arachidonic Acid and fully restoring liquid-crystal fluidity and steric hindrance.
* **Phase 3: Bioenergetic Optimization (Days 61-90+):**
* **Cardiolipin Repair:** Inner mitochondrial membrane undergoes lipidomic turnover. Fresh PUFAs restore cardiolipin spatial geometry.
* **Missile Launched:** Pathological proton leaks seal, mitochondrial transmembrane potential ($DeltaPsi_m$) restores, ATP synthase reboots. A new generation of fully optimized spermatozoa enters the ejaculate.
## VI. CONCLUSION: THE REPRODUCTIVE RENAISSANCE
* **The Defeat of the 15:1 Crisis:** The protocol identifies and eradicates the structural poison (Arachidonic Acid), dismantling the brittle biological glass and dampening the systemic pro-inflammatory amplifier.
* **The Triumph of the Keyora Matrix:** The 2-4:1 matrix liberates desaturase enzymes, rebuilds the liquid-crystal membrane with DHA, and supports bioenergetics via PPAR-alpha activation (driven by ALA and Oleic Acid) to ensure high fuel efficiency for propulsion.
* **The Astaxanthin Sovereign:** Astaxanthin is the absolute prerequisite. Without its 30-Angstrom transmembrane shield, lipidomic reconstruction is biologically impossible due to instant oxidation. By preserving the DNA phosphodiester backbone and quenching ROS, Astaxanthin is the sole protagonist responsible for the 54.5% conception surge.

Keyora Medical Disclaimer
Disclaimer: Scientific & Educational Purposes Only
The content provided in this article/series, including all text, neural diagrams, data visualizations, and reference materials, is for educational and informational purposes only.
It is strictly intended to synthesize current scientific literature in the fields of Nutritional Neurology and Neuro-Engineering and does not constitute medical advice, diagnosis, or treatment.
Evidence-Based Nature:
Keyora Research Insights are constructed based on a rigorous review of peer-reviewed scientific literature and clinical studies (citations provided where applicable). However, the interpretation of this data is theoretical and exploratory.
Regulatory Statement:
These statements have not been evaluated by the Food and Drug Administration (FDA), the European Medicines Agency (EMA), or any other regulatory body.
Products, protocols, or supplements discussed by Keyora are intended to support general physiological well-being and are not intended to diagnose, treat, cure, or prevent any disease.
Professional Consultation:
Individual biological responses vary. Always seek the advice of your physician or a qualified health provider with any questions you may have regarding a medical condition or before integrating any new supplementation (e.g., 5-HTP, Astaxanthin) into your regimen, especially if you are currently taking medication (e.g., SSRIs).
Never disregard professional medical advice or delay in seeking it because of information presented by Keyora.

By Keyora Research Notes Series
This article contributes to Keyora’s ongoing scientific documentation series, which systematically outlines the conceptual foundations, mechanistic pathways, and empirical evidence informing our research and development approach.
ORCID: 0009–0007–5798–1996